![]() Kalebe S. Pinto1
Kalebe S. Pinto1 ![]() ,
, ![]() Jansen Zuanon1,2 and
Jansen Zuanon1,2 and ![]() Tiago H. S. Pires1
Tiago H. S. Pires1
PDF: Download Here | Supplementary: Sup | Cite this article
Associate Editor: ![]() Eliane Gonçalves de Freitas
Eliane Gonçalves de Freitas
Section Editor: ![]() Fernando Pelicice
Fernando Pelicice
Editor-in-chief: ![]() José Birindelli
José Birindelli
Abstract
A navegação espacial é um componente central da ecologia de peixes, influenciando forrageamento, escolha de parceiros e defesa territorial, e pode diferir entre os sexos sob pressões ecológicas distintas. Investigamos se o tetra amazônico Crenuchus spilurus, uma espécie sexualmente dimórfica e dicromática, utiliza marcos visuais para orientação e se há diferenças sexuais na aprendizagem espacial. Em seu habitat natural, machos solitários são territorialistas e relativamente sedentários, defendendo ninhos onde realizam cuidado paternal exclusivo, enquanto as fêmeas deslocam-se em cardumes em busca de alimento e parceiros. Testamos indivíduos em um labirinto no qual deveriam localizar a saída correta orientando-se por um estímulo visual, permitindo avaliar aprendizagem e flexibilidade. Ambos os sexos apresentaram redução progressiva no tempo de resolução; contudo, somente as fêmeas mostraram evidências claras de aprendizagem baseada em acurácia, enquanto os machos mantiveram baixa precisão mesmo após repetições. Além disso, fêmeas completaram a tarefa mais rapidamente e com maior acurácia na correção de rota, indicando maior flexibilidade cognitiva e capacidade de atualização comportamental, possivelmente associadas à necessidade de explorar novos locais de reprodução. Machos, em contraste, exibiram um estilo de navegação mais rígido e menos eficiente, consistente com territorialidade e fidelidade espacial. Esses resultados indicam que fêmeas de C. spilurus utilizam marcos visuais para resolver tarefas espaciais e revelam estratégias cognitivas sexualmente diferenciadas.
Palavras-chave: Cognição, Diferenças baseadas no sexo, Habilidades espaciais, Navegação, Pistas visuais.
Introduction
Efficient and safe locomotion is crucial for vital activities such as foraging (Warburton, 2003; Rosati, 2017), searching for reproductive partners (Kotrschal et al., 2015a; Fuss, 2021), and escaping predators (Brown, Braithwaite, 2005; Kelley, Brown, 2011). Animals use multiple sensory cues as sources of directional information to orient themselves and navigate (Akre, Johnsen, 2014; Green et al., 2020), while also integrating past experiences into their spatial behavior, which leads to adaptation and learning processes (Kotrschal, Taborsky, 2010). Spatial orientation in environments with constantly changing sensory cues presents a challenge that drives the evolution of navigational skills. These skills require attention to signal variation, risk assessment, memory of stimuli, and making inferences from incomplete information, thereby supporting adaptive behavior across different contexts (Brydges et al., 2008; Mettke‐Hofmann, 2014; White, Brown, 2015; Reichert, Quinn, 2017; Cauchoix et al., 2020).
Animals employ diverse navigation strategies, ranging from simple movement rules to complex spatial mapping, which can be analyzed both in terms of behavior and the underlying neural mechanisms (Broglio, Salas, 2003; Rodríguez et al., 2021; Vinepinsky, Segev, 2023). Among these strategies, egocentric navigation stands out, in which individuals follow directional instructions (such as turning right or left) based on previous experiences, and allocentric navigation, in which they rely on global landmarks such as visual cues in the environment (Tolman, 1948; Rodriguez et al., 1994; Salas et al., 2017). Allocentric navigation allows the use of visual landmarks as direct, indirect, or sequential references, integrating information from different directions and establishing complex geometric relationships for spatial localization (Perera, Garcia, 2003; Perera, 2004; Braithwaite, Perera, 2006).
Sexual differences in spatial abilities have been documented in several animal groups (Jones et al., 2003; Healy, Hurly, 2004; Jozet-Alves et al., 2008; Perdue et al., 2011; Guigueno et al., 2014; Lucon-Xiccato, Bisazza, 2017a; Wallace et al., 2020). Males and females often make different decisions, even within the same social context (Van der Bijl et al., 2015). These differences frequently reflect distinct ecological pressures and sex-specific fitness demands (Watson, Platt, 2008; Kotrschal et al., 2015b; Keagy et al., 2019; Corral‐López et al., 2020). For example, in some bird and fish species, females exhibit greater mobility than males due to the need to locate spawning sites (Costa et al., 2011; Guigueno et al., 2014). This behavioral requirement in females is correlated with increased volume in brain regions linked to spatial cognition, whereas males of the same species, which do not need to engage in extensive searching for spawning sites, do not show the same enlargement in these brain areas (Guigueno et al., 2014). Conversely, in territorial species, across several vertebrate groups, including fishes, birds, and mammals, competition for mates has been shown to enhance spatial learning and cognitive performance, with the more territorial sex (generally males) displaying superior spatial performance (Jones et al., 2003; Saucier et al., 2008; Carazo et al., 2014). These findings highlight the significant influence of ecological and social pressures on sex differences in cognition.
Crenuchus spilurus Günther, 1863 is a sexually dimorphic and dichromatic species that inhabits clear-water Amazonian streams with structurally complex habitats (Pires et al., 2016). During reproduction, males display hypertrophied fins and bold courtship behaviors, whereas females indicate receptivity through abdominal darkening. Mate choice is mutual, and visual cues play a key role in this process (Pinto et al., 2021; Borghezan et al., 2023). Males are highly territorial and show strong site fidelity, defending fixed shelters that function both as refuges and nesting sites, where they provide exclusive paternal care (Pires et al., 2016, 2021). Males also exhibit fine visual discrimination when assessing rivals, avoiding escalated aggression with larger and/or more ornamented individuals (Pires et al., 2021). Previous work suggests that C. spilurus tends to rely on egocentric navigation when solving maze tasks (Pinto et al., 2021), but the species inhabits clear waters with low turbidity and abundant structurally complex (Pires et al., 2016) conditions in which visual landmarks may be ecologically relevant for orientation (Braithwaite, Perera, 2006; Sovrano et al., 2020). Given this ecological and behavioral context, we aimed to investigate whether C. spilurus can use visual landmarks for spatial navigation and whether males and females differ in this ability. Based on their territorial behavior and site fidelity, we hypothesized that males would exhibit greater spatial navigational ability than females. To test this, we employed a spatial task in which individuals had to follow visual cues to locate the exit of a maze.
Material and methods
Subjects. Adult Crenuchus spilurus of both sexes (Figs. 1A–B) were collected with small fine-mesh hand nets in a stream located in an urban forest area within the city of Manaus, Amazonas, Brazil (03º06’22.7”S 59º58’40.5”W) in December 2021. Our laboratory is located approximately 3 km from the sampling site, which allowed the transport of specimens from the field to the laboratory aquaria in less than 1 h, minimizing stress and potential injuries. To ensure welfare during transport, the fish were placed in plastic bags with water inside buckets equipped with portable aeration devices. Voucher specimens were deposited in the Fish Collection of the Instituto Nacional de Pesquisas da Amazônia (INPA), municipality of Manaus, Amazonas State, Brazil, as Crenuchus spilurus from the Negro River (INPA-ICT 61815). Fish were housed in the laboratory until the beginning of the experiments, which were conducted between April and September 2022.

FIGURE 1| Representative images of the sexually dimorphic Amazonian fish Crenuchus spilurus. A. Male; B. Female. Images reproduced from Borghezan et al. (2019).
Housing. Following housing procedures previously adopted in experimental studies with this species (Pinto et al., 2021; Pires et al., 2021), males and females were housed separately in aquaria (60 Length × 30 Width × 30 Depth cm), with a maximum density of 40 individuals per tank. This approach was used to reduce overt male-male aggressive interactions associated with territorial defense during the acclimation period. To simulate the natural environment, natural and artificial plants were added to each tank, along with approximately 10 sections of PVC tubes (10 cm length and 25 mm diameter) to provide shelter. Individuals of this species do not move extensively within the aquaria, and males defend territories only around the PVC tubes; therefore, aggressive interactions were rarely observed in the holding tanks. Each tank was equipped with a mechanical filter and an air pump, and weekly water changes were carried out to maintain water quality. Laboratory water supply originated from groundwater, and the only difference compared to stream water quality was the lack of dissolved organic carbon. To compensate for this, we regularly collected dead leaves from the forest stream and added them to a 100-liter reservoir, simulating the natural incorporation of dissolved organic carbon into the water, following procedures previously adopted in studies from our group (Pinto et al., 2021; Pires et al., 2021). Water was kept in this reservoir for at least three days before being used in the tanks. The laboratory received indirect natural light through large glass windows, and since it is located in an equatorial region (Manaus, Amazonas State, Brazil; 03°05’45”S 59°59’21”W), it followed a natural 12:12 h light-dark cycle. On experimental days, artificial lights were turned on from 08:00 to 18:00 to ensure consistent illumination during setup and testing. On non-experimental days, tanks were exposed primarily to the natural daylight entering the room. The room was maintained at a constant temperature of 24 °C with air conditioning, simulating the prevailing water temperature of Central Amazonian forest streams. Fish were fed twice daily with high-quality commercial ornamental fish food (Sera GVG Mix, granules), which was readily consumed by the individuals. Only adult individuals were included in the study.
Apparatus. The experiments were conducted in a glass tank (70 L × 30 W × 20 D cm) filled with a 15 cm water column (Fig. 2). To ensure that no visual cues were available beyond those provided in the experimental setup, the tank walls were externally covered with opaque cloth. The experimental apparatus (50 L × 25 W × 3 D cm) was built using green plastic bricks (Lego Bricks, Billund, Denmark), positioned at the center of the tank. The apparatus consisted of an initial corridor leading to a first sector (9 L × 22 W × 3 D cm) with two exits: one to the right and one to the left. One exit led to a dead end, while the other led to a second sector, with the same dimensions, which also had two exits, one leading to a dead end and the other to the goal zone. The correct paths connecting sector 1 to sector 2 and sector 2 to the goal zone were marked with a red visual cue, which was always placed in opposite positions in each sector. A glass plate was placed on top of the apparatus, maintaining a 3 cm water column to restrict the vertical movement of the fish. Restricting water column height functions as a stimulus for fish to swim forward and solve the spatial task (Kellogg, Gavin, 1960; Lucon-Xiccato, Bisazza, 2017b). The remaining spaces of approximately 10 × 30 cm at each end of the tank were used as the initial sector, where the fish was placed before the trial, and as the goal sector, reached upon solving the task. These two sectors were connected by a lateral corridor approximately 5 cm wide, allowing the fish to be guided back from the goal sector to the initial sector without being removed from the water. In the initial sector, we built a shelter with two open ends made of green plastic bricks (3 L × 6 W × 3 D cm), directly attached to the experimental apparatus, to facilitate guiding the individuals into the initial corridor. All individuals used this structure from the start of habituation. In the goal sector, located at the opposite side of the tank, we placed as a reward a PVC tube identical to those in the housing aquaria. Additionally, the goal sector was adjacent to a smaller aquarium (30 L × 20 W × 25 D cm, not connected to the apparatus) containing social stimuli (four females of C. spilurus), which served as an additional reward. We used four females to provide a stronger and more stable social stimulus, as males of this species are highly motivated to approach and court potential reproductive partners, and females are naturally gregarious and tend to remain in shoals with other females (Pires et al., 2016). The laboratory space used to house the apparatus was surrounded by opaque cloth to prevent visual contact with the experimenter and was indirectly illuminated by room lights. A video camera (Logitech C920) mounted above the tank recorded the trials.
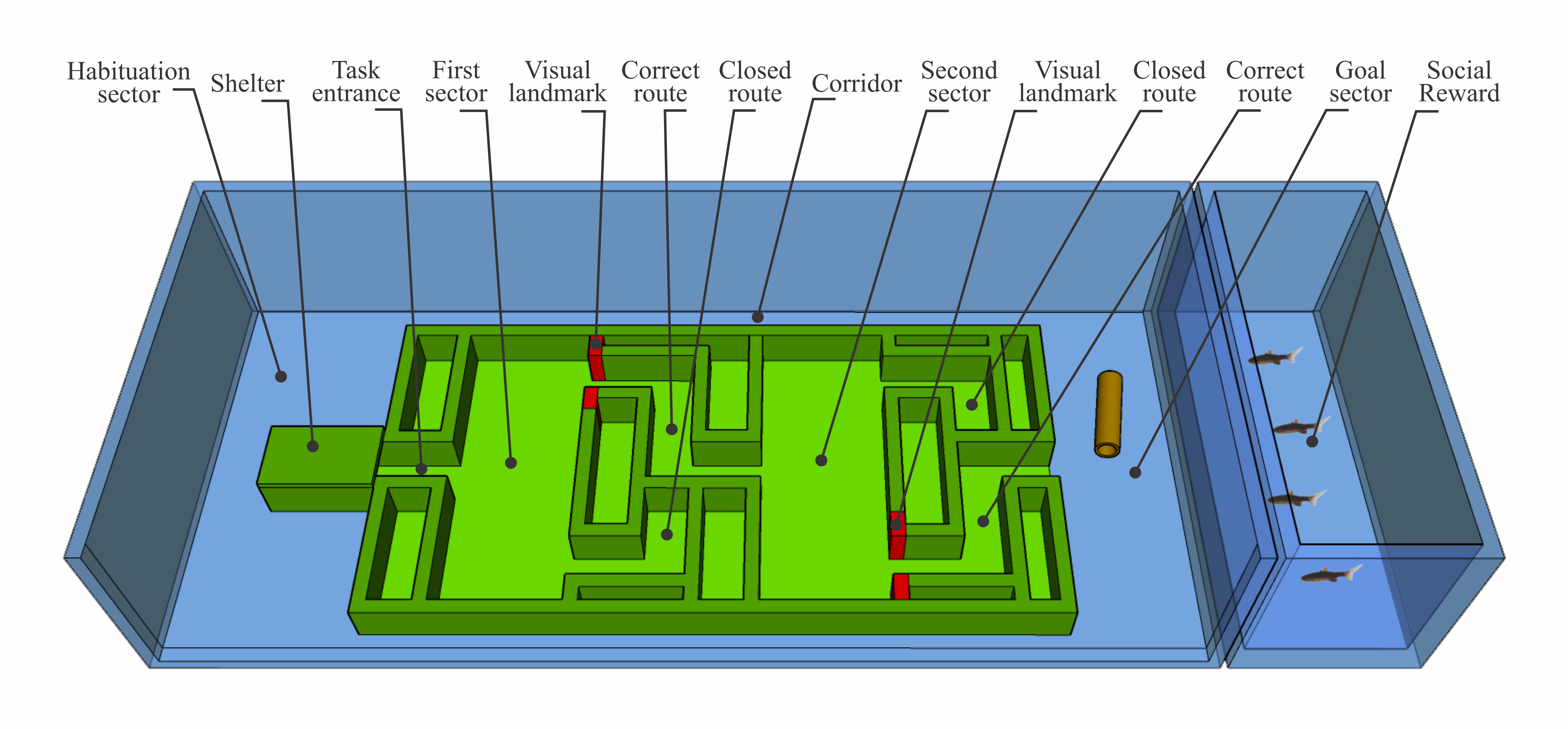
FIGURE 2| 3D schematic of the task used in the spatial learning experiment.
Procedure. Initially, an individual was transferred from the holding aquarium to the initial sector of the apparatus 24 h before testing for habituation. Access to the test area was blocked by an opaque sliding panel, which also prevented visual access to stimuli. After the habituation period, with the individual inside the shelter leading to the test area, the opaque panel was removed, and a plastic brick was gently inserted at the opposite end of the shelter, encouraging the fish to slowly swim into the maze. At this point, with the fish free to decide when to advance to the next sectors, recording began. The experimenter observed the test remotely via a monitor connected to the camera. After the individual reached the goal sector, it remained there for 5 min close to the social reward in the adjacent aquarium. Individuals who took longer than 20 min to complete the test (two males and three females) were removed from the experiment and replaced by naïve individuals from the holding aquarium (see Video S1 for a representative example of the task). The sample size was 25 males (Standard length, SL 3.92 ± 0.18 cm) and 25 females (SL 3.67 ± 0.14 cm). At the end of each trial, the tested individual was gently guided back to the initial sector through the lateral corridor while remaining underwater. Then, the glass plate covering the top of the apparatus was removed, and with the aid of a brush, the water inside the apparatus was gently mixed to homogenize potential chemical cues. The spatial task was then reconfigured. First, the corridors were switched in both sectors, but the correct paths were always maintained on opposite sides of the two sectors. The visual cue was also repositioned, always indicating the correct corridor. When correct choices have remained fixed in the environment, it becomes difficult to determine whether animals are using egocentric learning (repeating a directional turn) or allocentric learning (using environmental landmarks) (Tolman, 1948). Therefore, by alternating the correct side in each training session, learning could only occur if individuals relied on the relative position of the visual landmark rather than on previously correct directional patterns. In this framework, route correction refers to the fish’s ability to avoid repeating a direction that had been correct in previous trials and instead adjust its trajectory based on the visual cue presented in the current session. All experimental trials were conducted between 10:00 and 18:00. After these procedures, the experimental session was repeated again after 1 h, and all five trials for each individual were conducted on the same day. Thus, each fish completed five consecutive trials within a single day, after which individuals were photographed and returned to their respective housing aquaria (males or females).
Statistical analyses. From the video recordings, we verified whether the first route chosen by each individual was correct or incorrect to calculate choice accuracy. We also recorded the time to solve the task, defined as the time in seconds from the moment the fish entered the first sector of the test until it reached the goal sector. To analyze task resolution time, we used a linear mixed-effects model (LMM) with log-transformed time as the dependent variable. In this model, trial number (1 to 5) and sex (male or female) were considered fixed effects, and individual identity was included as a random effect. To assess choice accuracy, we fitted a generalized linear mixed-effects model (GLMM) using the R package lme4 (Bates et al., 2015). The dependent variable was the binary choice of each individual in each trial (correct or incorrect). Trial (1 to 5), sex (male or female), and sector (first or second) were included as fixed effects. Individual identity was included as a random effect to control for the repeated-measures structure. We did not include fish standard length in either model because males are generally larger than females in this species, resulting in collinearity between sex and standard length (Pires et al., 2016). Additionally, in a previous study, we found an effect of male ornamentation in spatial learning (Pinto et al., 2021). Since the ornamentation degree is only available for males, we tested its effect on time and accuracy beforehand. As it showed no significant effect (time: F1,23 = 0.32, p = 0.577; accuracy: χ²(1) = 0.156, p = 0.692), ornamentation was excluded from the final models, allowing symmetric comparisons between sexes with the same fixed predictors. Finally, to evaluate route correction, i.e., accuracy specifically in the second sector, we assessed the influence of sex using a generalized linear mixed-effects model (GLMM) with a binomial distribution and logit link function. The dependent variable was the binary response (correct or incorrect) in the second sector, which directly reflects route-correction performance, and sex was included as the explanatory factor. Individual identity was again included as a random effect to control for repeated measures on the same animal. Predicted accuracy was obtained from model coefficients using a logistic transformation. All statistical analyses and graphical representations were conducted in R software v. 4.2.0 (R Development Core Team, 2022). Statistical tests were two-tailed, with significance set at p = 0.05.
Results
Time to solve the task. The interaction between test sequence (1 to 5) and sex was not significant (F4,442 = 0.793, p = 0.529), indicating that males and females showed a similar pattern of improvement across trials. Considering the main effects, resolution time progressively decreased across tests (F4,442 = 5.618, p < 0.001), reflecting improved performance with repetition. Additionally, males took significantly longer to complete the task than females (F1,48 = 9.299, p = 0.003). Together, these results show that although both sexes improved with experience, females consistently solved the task more quickly (Fig. 3A; see also Fig. S2 for individual-level distributions and Tab. 1 for a summary of statistical effects).
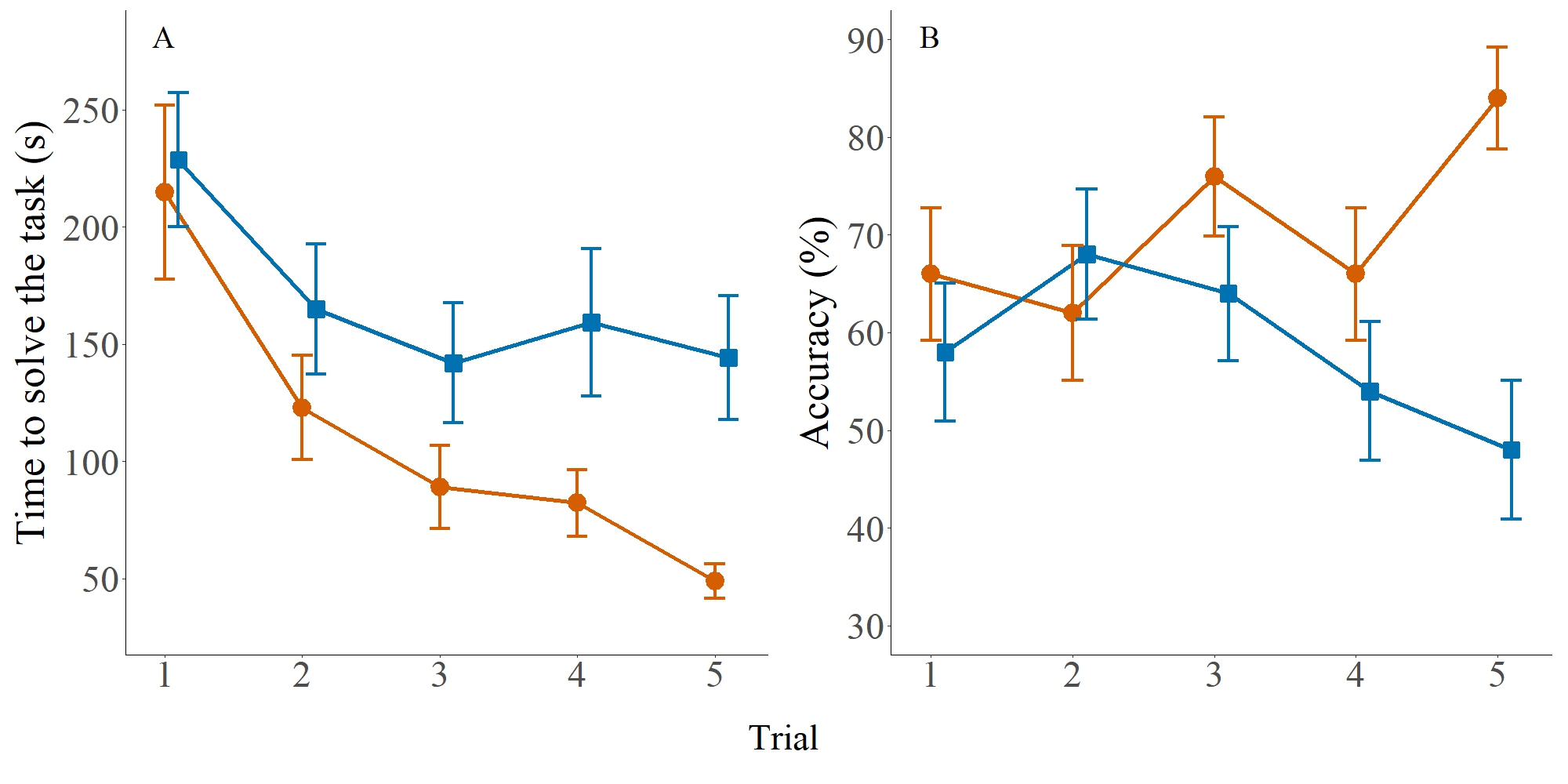
FIGURE 3| Spatial performance of male and female Crenuchus spilurus across five trials. A. Time to solve the task. B. Accuracy (%) of choices across trials. Individual data points are shown for each trial, with horizontal lines representing mean values and vertical bars indicating SEM. Males are shown in blue and females in red. Sample size: N = 25 males and 25 females.
TABLE 1 | Summary of the statistical effects of sex, test sequence, sector, and their interactions on performance measures (resolution time, accuracy, and route correction) during the spatial navigation task. Significant effects are indicated with an asterisk. df = degrees of freedom.
Factors | Statistic | df | p-value |
Resolution time | |||
Test sequence | F = 5.618 | 4.442 | < 0.001 * |
Sex | F = 9.299 | 1.48 | 0.003 * |
Test x Sex | F = 0.793 | 4.442 | 0.529 |
Accuracy (overall) | |||
Test sequence | χ² = 7.725 | 4 | 0.102 |
Sex | χ² = 0.655 | 1 | 0.418 |
Sector | χ² = 0.460 | 1 | 0.497 |
Test x Sex | χ² = 10.417 | 4 | 0.033 * |
Accuracy (males) | |||
Test sequence | χ² = 7.148 | 4 | 0.128 |
Sector | χ² = 0.088 | 1 | 0.766 |
Test x Sector | χ² = 3.714 | 4 | 0.446 |
Accuracy (females) | |||
Test sequence | χ² = 6.374 | 4 | 0.173 |
Sector | χ² = 4.159 | 1 | 0.041 * |
Test x Sector | χ² = 9.241 | 4 | 0.055 |
Route correction | |||
Sex | χ² = 7.86 | 1 | 0.005 * |
Accuracy. The GLMM conducted to evaluate route choice accuracy revealed no significant effect of test sequence (χ²(4) = 7.725, p = 0.102), sex (χ²(1) = 0.655, p = 0.418), or sector (χ²(1) = 0.460, p = 0.497). However, there was a significant interaction between test and sex (χ²(4) = 10.417, p = 0.033), suggesting that the pattern of accuracy across trials differed between males and females (Fig. 3B). Therefore, we performed two separate GLMMs for males and females. In both models, the effect of test was not significant (males: χ²(4) = 7.148, p = 0.128; females: χ²(4) = 6.374, p = 0.173), nor was the interaction between test and sector (males: χ²(4) = 3.714, p = 0.446; females: χ²(4) = 9.241, p = 0.055). For females, the test-by-sector interaction was close to, but did not reach, the significance threshold defined in our statistical procedures. Although this pattern hints at a possible tendency, it does not provide sufficiently clear evidence to support conclusions about learning across trials. The effect of sector was not significant for males (χ²(1) = 0.088, p = 0.766), but was significant for females (χ²(1) = 4.159, p = 0.041), which showed increased accuracy in the second sector (see Tab. 1 for a summary of statistical effects).
Route correction. The GLMM analyzing the influence of sex on accuracy in the second sector revealed a significant effect on the probability of a correct choice (χ²(1) = 7.86, p = 0.005, Fig. 4), indicating that males exhibited lower accuracy compared to females. Females showed an estimated probability of success of approximately 72%, while for males this probability was about 54%. The variance of the random effect by individual was low (σ² = 0.014), suggesting that most of the variation in performance was explained by sex (see Fig. S3 for individual accuracy distributions and Tab. 1 for a summary of statistical effects).
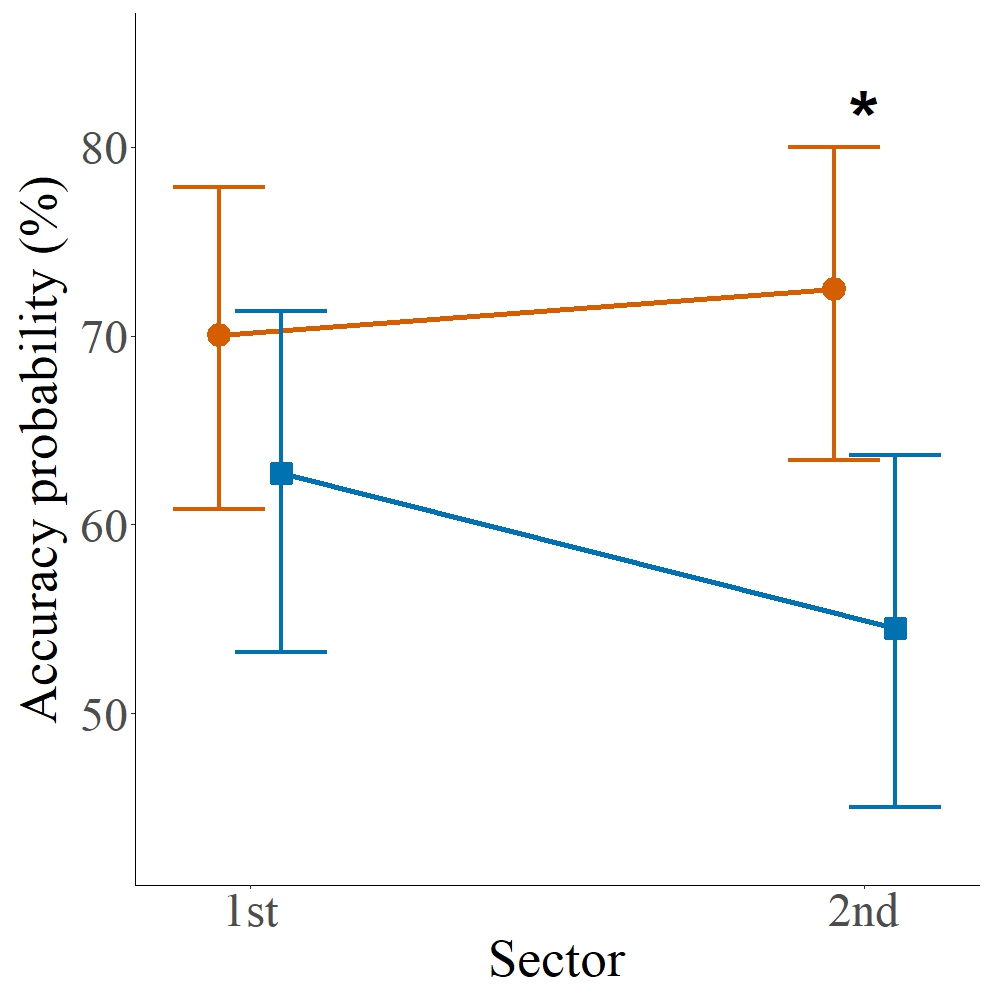
FIGURE 4| Interaction between sex and sector in the accuracy of choices by Crenuchus spilurus individuals. Points and vertical bars represent the mean probability of correct responses (with 95% confidence intervals) for males (blue) and females (red) in the two experimental sectors. Lines connect sectors for each sex, and the asterisk indicates a significant effect.
Discussion
Our results indicate that female Crenuchus spilurus are capable of using visual landmarks to solve spatial navigation tasks, and that males and females differ in how they process and integrate spatial information. While both sexes showed a progressive decrease in resolution time across trials, males consistently took longer to complete the task and did not improve in accuracy across sessions. Accuracy, a most robust measure of learning, as it is less influenced by habituation or motivational changes, revealed clear evidence of landmark-based learning only in females. Males, in contrast, maintained low accuracy even after repeated exposure, indicating that they did not learn the visual cue. This pattern aligns with the species’ ecology, as C. spilurus inhabits clear-water streams rich in vegetation (Pires et al., 2016), environments that favor the use of stable visual references (Odling‐Smee, Braithwaite, 2003; Akre, Johnsen, 2014). Our findings reinforce the broader evidence that spatial cognition is well developed in teleost fishes, especially in ecologically relevant contexts (Brown, Laland, 2003; Salas et al., 2006; Sovrano et al., 2020).
Although both sexes reduced their resolution time across trials, only females showed accuracy patterns consistent with learning the landmark-based task. Contrary to our initial assumption, females completed the task faster and with higher accuracy when correcting their route in the second sector, a pattern suggesting greater cognitive flexibility, potentially linked to broader ecological demands for spatial exploration. In many species, including fishes and birds, females are responsible for locating suitable spawning or foraging sites and are therefore under selective pressure to develop adaptable and efficient navigation strategies (Healy, Hurly, 2004; Guigueno et al., 2014). In this study, our interpretation of “mobility” refers specifically to ecological mobility, broader space uses, and exploratory range in the natural habitat, rather than increased locomotor activity inside the experimental apparatus. Field observations support this distinction: females of C. spilurus exhibit greater ecological mobility in the wild, whereas males show territorial fidelity and remain within small structured areas (Pires et al., 2016). Thus, we do not suggest that females performed better because they physically moved more within the apparatus, but rather that long-term ecological differences in space use may shape sex-specific spatial strategies. Consistent with this interpretation, the test-by-sector interaction in females approached, but did not reach, the predefined statistical threshold; although not conclusive, this trend aligns with the broader pattern observed and may indicate a subtle tendency toward more efficient integration of visual information across task phases, warranting further investigation in future studies with larger samples or refined designs.
Interestingly, males showed lower accuracy, particularly in the phase requiring integration of new visual information after an initial choice (i.e., the second sector of the maze). This limitation may reflect a cognitive bias toward more rigid, possibly egocentric, strategies in which trajectories are based on prior movements rather than on environmental landmarks (Braithwaite, Perera, 2006; Salas et al., 2006). In territorial species such as male C. spilurus, this style may suffice for solving spatial problems in familiar areas but is less effective when the task requires flexibility and constant information updating. This distinction between sexual cognitive styles, with females favoring allocentric navigation and males relying more on fixed-route (egocentric) navigation, is consistent with widely documented patterns in vertebrates, including rodents, birds, and humans (Saucier et al., 2002; Jonasson, 2005; Lucon-Xiccato, Bisazza, 2017c).
On the other hand, previous studies with C. spilurus have shown that more ornamented males, though more cautious, learn quickly in maze contexts (without visual cues), making fewer errors over time (Pinto et al., 2021). This combination of hesitation and efficient learning may reflect the “asset protection” principle (Clark, 1994), according to which individuals that have substantial ongoing reproductive investment, such as male C. spilurus, who defend territories and provide paternal care, tend to adopt more conservative, though not necessarily less effective, strategies in challenging contexts. Thus, the longer resolution times observed in males in this study do not necessarily indicate lower cognitive ability but may reflect a deliberative style that enhances safety. This interpretation reinforces the importance of considering personality traits and individual reproductive value as intra-sexual modulators of cognition, as has been evidenced in several animal groups (Gosling, 2008; Guillette et al., 2017).
An additional consideration concerns the potential difference in the salience of the social reward used in the task. The reward consisted of access to a sheltered PVC tube in an adjacent aquarium containing four conspecific females, a stimulus that likely carries distinct ecological meanings for each sex. For males, the presence of multiple females may represent a direct reproductive opportunity, whereas for females, it more likely provides social companionship and safety, consistent with the shoaling tendencies known for this species (Pires et al., 2016). Although the motivational salience of this reward may therefore differ between sexes, it represents a positive stimulus for both males and females. Importantly, there is no evidence of female-female competition for access to males in C. spilurus, nor indications that females are less motivated by social stimuli. Thus, differences in reward salience alone are unlikely to account for the observed sex differences in accuracy, learning patterns, and route correction. Instead, the results are more parsimoniously explained by sex-specific spatial strategies and differences in cognitive flexibility shaped by ecological and reproductive roles.
Furthermore, our results suggest that males and females adopted distinct problem-solving strategies across trials. Females were more efficient in adapting to spatial changes in the task, particularly when it was necessary to correct choices based on new visual information. This improved performance is consistent with greater behavioral flexibility and more effective inhibition of previously learned but incorrect responses, patterns that have been repeatedly reported in other teleosts (Lucon-Xiccato, Bisazza, 2014; Miletto Petrazzini et al., 2017; Lucon-Xiccato et al., 2020). Such inhibitory control is closely linked to executive functions, including cognitive flexibility and behavioral updating, as emphasized in recent reviews on sex differences in animal cognition (Lucon-Xiccato, 2022). The involvement of working memory and contextual integration mechanisms, cognitive processes essential for navigation in complex or unpredictable environments, may also contribute to female performance (Cauchoix et al., 2017; Vinepinsky, Segev, 2023). Males, in contrast, appear to rely on more fixed and less adaptive (egocentric) spatial rules, which may be advantageous in stable environments but hinder performance in uncertainty. In this context, it is noteworthy that C. spilurus shows higher abundance in partially dammed streams (Pires et al., 2016), where reduced water flow and lower structural rearrangement may create a more stable scenario. Under such conditions, the advantages associated with more flexible navigation may be attenuated, reducing the functional consequences of sexual differences in navigation and potentially favoring population-level reproductive success. Although speculative, this hypothesis underscores the importance of investigating how environmental stability modulates the expression and adaptive relevance of sex-specific cognitive strategies.
From an ecological and evolutionary perspective, the sex differences observed in this study may result from distinct pressures related to space use and social organization. In C. spilurus, male courtship and nest defense behavior depend strongly on maintaining fixed territories, often associated with specific structures such as submerged palm leaves or tubular shelters (Pires et al., 2021). Females, in contrast, need to locate and evaluate multiple territories and potential mates, which may favor greater versatility in navigation skills. This pattern of sexual specialization aligns with theoretical frameworks proposing that cognitive dimorphism can emerge when males and females consistently occupy different spatial or ecological niches (Gaulin, 1992; Watson, Platt, 2008).
This aligns with the “sexually dimorphic adaptive cognition hypothesis”, according to which distinct selective pressures shape cognitive specializations between sexes when their ecological demands consistently diverge. Geary (1996) argued that sex differences in cognitive skills arise from sexual selection and contrasting reproductive strategies between males and females, favoring, for example, greater efficiency in navigation/space use in the more mobile or competitive sex. Complementarily, Jacobs (1996) proposed the “sex-based-dispersal hypothesis”, suggesting that differences in spatial mobility between sexes, such as greater male dispersal or the need for females to return to the nest, may impose distinct pressures on navigation and spatial memory. In the case of C. spilurus, while males are territorial, site-faithful, and perform exclusive paternal care, females must explore the environment in search of feeding grounds and mates, fitting the model predicted by these theories. Accordingly, the differences observed here may reflect the action of natural selection shaping sex-specific cognitive profiles that are potentially adapted to the ecological challenges experienced by each sex in its respective niche.
The results presented here contribute to a broader understanding of how cognition, behavior, and ecology interact in the evolution of sex differences. They reinforce the role of fishes as valuable models for investigating adaptive cognition, highlighting that even small-bodied and cryptic species such as C. spilurus display sophisticated cognitive repertoires, sensitive to ecological context and differential selective pressures between sexes. Moreover, our data suggest that spatial cognition should not be assessed in a generalized manner but should take into account not only sex and environment but also the functional context of the task, individual cognitive style, and the species’ ecological-evolutionary history.
In conclusion, this study reveals that female C. spilurus exhibits navigation skills based on visual cues, while males did not show accuracy improvement and therefore did not demonstrate learning of the landmark-based task. The results also reveal modulation by individual experience and sex differences in spatial learning. Females demonstrate greater flexibility and accuracy in spatial tasks, a pattern that suggests a predominantly allocentric navigation strategy, whereas the combination of lower accuracy and longer resolution times in males may suggest that they rely more heavily on egocentric navigation and potentially engage in more cautious or conservative exploratory strategies. Such conclusions underscore the value of ecological and comparative approaches in understanding the evolution of animal cognition and position C. spilurus as a promising model for further studies on sexually differentiated cognition. Future studies exposing C. spilurus to reversal learning tasks may reveal with greater detail the sex differences in cognitive flexibility and behavioral updating capacity under changing contingencies, as suggested by the present results.
Acknowledgments
We thank Hélio D. Beltrão dos Anjos for his support in collecting the fish used in the study. We thank CNPq, CAPES and FAPEAM for long-term support to the Igarapés Project and for providing scholarship grants for several graduate students involved in the study. We thank Museu na Floresta (JICA) for financing the acquisition of equipment. We thank INPA for logistical support and access to research facilities.
References
Akre KL, Johnsen S. Psychophysics and the evolution of behavior. Trends Ecol Evol. 2014; 29(5):291–300. https://doi.org/10.1016/j.tree.2014.03.007
Bates D, Mächler M, Bolker B, Walker S. Fitting linear mixed-effects models using lme4. J Stat Softw. 2015; 67:1–48. https://doi.org/10.18637/jss.v067.i01
Borghezan EA, Pinto KS, Zuanon J, Pires THS. Someone like me: Size-assortative pairing and mating in an Amazonian fish, sailfin tetra Crenuchus spilurus. PLoS ONE. 2019; 14(9):e0222880. https://doi.org/10.1371/journal.pone.0222880
Borghezan EA, Pires THS, Zuanon J, Kohshima S. Effect of light bias on male mating signal and female mate choice in a sexually dimorphic Amazon fish. Behav Process. 2023; 213:104958. https://doi.org/10.1016/j.beproc.2023.104958
Braithwaite VA, Perera TB. Short-range orientation in fish: how fish map space. Mar Freshw Behav Physiol. 2006; 39(1):37–47. https://doi.org/10.1080/10236240600562844
Broglio C, Rodríguez F, Salas C. Spatial cognition and its neural basis in teleost fishes. Fish Fish. 2003; 4(3):247–55. https://doi.org/10.1046/j.1467-2979.2003.00128.x
Brown C, Braithwaite VA. Effects of predation pressure on the cognitive ability of the poeciliid Brachyraphis episcopi. Behav Ecol. 2005; 16(2):482–87. https://doi.org/10.1093/beheco/ari016
Brown C, Laland KN. Social learning in fishes: a review. Fish Fish. 2003; 4(3):280–88. https://doi.org/10.1046/j.1467-2979.2003.00122.x
Brydges NM, Heathcote RJ, Braithwaite VA. Habitat stability and predation pressure influence learning and memory in populations of three-spined sticklebacks. Anim Behav. 2008; 75(3):935–42. https://doi.org/10.1016/j.anbehav.2007.08.005
Carazo P, Noble DW, Chandrasoma D, Whiting MJ. Sex and boldness explain individual differences in spatial learning in a lizard. Proc Biol Sci. 2014; 281(1782):20133275. https://doi.org/10.1098/rspb.2013.3275
Cauchoix M, Hermer E, Chaine AS, Morand-Ferron J. Cognition in the field: comparison of reversal learning performance in captive and wild passerines. Sci Rep. 2017; 7(1):12945. https://doi.org/10.1038/s41598-017-13179-5
Cauchoix M, Chaine AS, Barragan-Jason G. Cognition in context: plasticity in cognitive performance in response to ongoing environmental variables. Front Ecol Evol. 2020; 8:106. https://doi.org/10.3389/fevo.2020.00106
Clark CW. Antipredator behavior and the asset-protection principle. Behav Ecol. 1994; 5(2):159–70. https://doi.org/10.1093/beheco/5.2.159
Corral-López A, Romensky M, Kotrschal A, Buechel SD, Kolm N. Brain size affects responsiveness in mating behaviour to variation in predation pressure and sex ratio. J Evol Biol. 2020; 33(2):165–77. https://doi.org/10.1111/jeb.13556
Costa SS, Andrade R, Carneiro LA, Gonçalves EJ, Kotrschal K, Oliveira RF. Sex differences in the dorsolateral telencephalon correlate with home range size in blenniid fish. Brain Behav Evol. 2011; 77(1):55–64. https://doi.org/10.1159/000323668
Fuss T. Mate choice, sex roles and sexual cognition in vertebrates: mate choice turns cognition or cognition turns mate choice? Front Ecol Evol. 2021; 9:749495. https://doi.org/10.3389/fevo.2021.749495
Gaulin SJ. Evolution of sex difference in spatial ability. Am J Biol Anthropol. 1992; 35(S15):125–51. https://doi.org/10.1002/ajpa.1330350606
Geary DC. Sexual selection and sex differences in mathematical abilities. Behav Brain Sci. 1996; 19(2):229–47. https://doi.org/10.1017/S0140525X00042400
Gosling SD. Personality in non‐human animals. Soc Personal Psychol Compass. 2008; 2(2):985–1001. https://doi.org/10.1111/j.1751-9004.2008.00087.x
Green PA, Brandley NC, Nowicki S. Categorical perception in animal communication and decision-making. Behav Ecol. 2020; 31(4):859–67. https://doi.org/10.1093/beheco/araa004
Guigueno MF, Snow DA, MacDougall-Shackleton SA, Sherry DF. Female cowbirds have more accurate spatial memory than males. Biol Lett. 2014; 10(2):20140026. https://doi.org/10.1098/rsbl.2014.0026
Guillette LM, Naguib M, Griffin AS. Individual differences in cognition and personality. Behav Process. 2017; 134:1–03. https://psycnet.apa.org/doi/10.1016/j.beproc.2016.12.001
Healy SD, Hurly TA. Spatial learning and memory in birds. Brain Behav Evol. 2004; 63(4):211–20. https://doi.org/10.1159/000076782
Jacobs LF. Sexual selection and the brain. Trends Ecol Evol. 1996; 11(2):82–86. https://doi.org/10.1016/0169-5347(96)81048-2
Jonasson Z. Meta-analysis of sex differences in rodent models of learning and memory: a review of behavioral and biological data. Neurosci Biobehav Rev. 2005; 28(8):811–25. https://doi.org/10.1016/j.neubiorev.2004.10.006
Jones CM, Braithwaite VA, Healy SD. The evolution of sex differences in spatial ability. Behav Neurosci. 2003; 117(3):403. https://psycnet.apa.org/doi/10.1037/0735-7044.117.3.403
Jozet-Alves C, Modéran J, Dickel L. Sex differences in spatial cognition in an invertebrate: the cuttlefish. Proc Biol Sci. 2008; 275(1646):2049–54. https://doi.org/10.1098/rspb.2008.0501
Keagy J, Minter R, Tinghitella RM. Sex differences in cognition and their relationship to male mate choice. Curr Zool. 2019; 65(3):285–93. https://doi.org/10.1093/cz/zoz014
Kelley JL, Brown C. Predation risk and decision making in poeciliid prey. In Evans JP, Pilastro A, Schlupp I, editors. Ecology and evolution of poeciliid fishes. Chicago: University of Chicago Press; 2011. p.174–84.
Kellogg WN, Gavin J. Maze-learning in the guppy. Psychol Rep. 1960; 6(3):445–46. https://doi.org/10.2466/pr0.1960.6.3.445
Kotrschal A, Taborsky B. Environmental change enhances cognitive abilities in fish. PLoS Biol. 2010; 8(4):e1000351. https://doi.org/10.1371/journal.pbio.1000351
Kotrschal A, Corral-Lopez A, Amcoff M, Kolm N. A larger brain confers a benefit in a spatial mate search learning task in male guppies. Behav Ecol. 2015a; 26(2):527–32. https://doi.org/10.1093/beheco/aru227
Kotrschal A, Buechel SD, Zala SM, Corral-Lopez A, Penn DJ, Kolm N. Brain size affects female but not male survival under predation threat. Ecol Lett. 2015b; 18(7):646–52. https://doi.org/10.1111/ele.12441
Lucon-Xiccato T. The contribution of executive functions to sex differences in animal cognition. Neurosci Biobehav Rev. 2022; 138:104705. https://doi.org/10.1016/j.neubiorev.2022.104705
Lucon-Xiccato T, Bisazza A. Discrimination reversal learning reveals greater female behavioural flexibility in guppies. Biol Lett. 2014; 10(6):20140206. https://doi.org/10.1098/rsbl.2014.0206
Lucon-Xiccato T, Bisazza A. Sex differences in spatial abilities and cognitive flexibility in the guppy. Anim Behav. 2017a; 123:53–60. https://doi.org/10.1016/j.anbehav.2016.10.026
Lucon-Xiccato T, Bisazza A. Complex maze learning by fish. Anim Behav. 2017b; 125:69–75. https://doi.org/10.1016/j.anbehav.2016.12.022
Lucon-Xiccato T, Bisazza A. Individual differences in cognition among teleost fishes. Behav Process. 2017c; 141:184–95. https://doi.org/10.1016/j.beproc.2017.01.015
Lucon-Xiccato T, Montalbano G, Bertolucci C. Personality traits covary with individual differences in inhibitory abilities in 2 species of fish. Curr Zool. 2020; 66:187–95. https://doi.org/10.1093/cz/zoz039
Mettke-Hofmann C. Cognitive ecology: ecological factors, life-styles, and cognition. Wiley Interdiscip Rev Cogn Sci. 2014; 5(3):345–60. https://doi.org/10.1002/wcs.1289
Miletto Petrazzini ME, Bisazza A, Agrillo C, Lucon-Xiccato T. Sex differences in discrimination reversal learning in the guppy. Anim Cogn. 2017; 20(6):1081–91. https://doi.org/10.1007/s10071-017-1124-4
Odling-Smee L, Braithwaite VA. The role of learning in fish orientation. Fish Fish. 2003; 4(3):235–46. https://doi.org/10.1046/j.1467-2979.2003.00127.x
Perdue BM, Snyder RJ, Zhihe Z, Marr MJ, Maple TL. Sex differences in spatial ability: a test of the range size hypothesis in the order Carnivora. Biol Lett. 2011; 7(3):380–83. https://doi.org/10.1098/rsbl.2010.1116
Perera TB, Garcia CM. Amarillo fish (Girardinichthys multiradiatus) use visual landmarks to orient in space. Ethology. 2003; 109(4):341–50. https://doi.org/10.1046/j.1439-0310.2003.00876.x
Perera TB. Fish can encode order in their spatial map. Proc Biol Sci. 2004; 271(1553):2131–34. https://doi.org/10.1098/rspb.2004.2867
Pinto KS, Saenz DE, Borghezan EA, Pires THS. Attractive males are cautious and better learners in the sailfin tetra. Anim Behav. 2021; 172:103–11. https://doi.org/10.1016/j.anbehav.2020.12.005
Pires THS, Farago TB, Campos DF, Cardoso GM, Zuanon J. Traits of a lineage with extraordinary geographical range: ecology, behavior and life-history of the sailfin tetra Crenuchus spilurus. Environ Biol Fishes. 2016; 99:925–37. https://doi.org/10.1007/s10641-016-0534-5
Pires THS, Pinto KS, Borghezan EA, Zuanon J. Dominant males exploit the courtship effort of subordinate males in an Amazonian fish. Anim Behav. 2021; 178:185–93. https://doi.org/10.1016/j.anbehav.2021.05.023
R Development Core Team. R: a language and environment for statistical computing [Internet]. Vienna, Austria: R Foundation for Statistical Computing; 2022. Available from: http://www.rproject.org
Reichert MS, Quinn JL. Cognition in contests: mechanisms, ecology, and evolution. Trends Ecol Evol. 2017; 32(10):773–85. https://doi.org/10.1016/j.tree.2017.07.003
Rodriguez F, Duran E, Vargas JP, Torres B, Salas C. Performance of goldfish trained in allocentric and egocentric maze procedures suggests the presence of a cognitive mapping system in fishes. Anim Learn Behav. 1994; 22(4):409–20. https://doi.org/10.3758/BF03209160
Rodríguez F, Quintero B, Amores L, Madrid D, Salas-Peña C, Salas C. Spatial cognition in teleost fish: strategies and mechanisms. Animals. 2021; 11(8):2271. https://doi.org/10.3390/ani11082271
Rosati AG. Foraging cognition: reviving the ecological intelligence hypothesis. Trends Cogn Sci. 2017; 21(9):691–702. https://doi.org/10.1016/j.tics.2017.05.011
Salas C, Broglio C, Durán E, Gómez A, Ocaña FM, Jiménez-Moya F et al. Neuropsychology of learning and memory in teleost fish. Zebrafish. 2006; 3(2):157–71. https://doi.org/10.1089/zeb.2006.3.157
Salas C, Broglio C, Durán E, Ocaña FM, Martín-Monzón I, Gómez A et al. Spatial learning and its neural basis in fish. In: Byrne JH, editor. Learning and memory: a comprehensive reference. Second edition. Oxford: Academic Press; 2017. p.347–73.
Saucier DM, Green SM, Leason J, MacFadden A, Bell S, Elias LJ. Are sex differences in navigation caused by sexually dimorphic strategies or by differences in the ability to use the strategies? Behav Neurosci. 2002; 116(3):403. https://psycnet.apa.org/doi/10.1037/0735-7044.116.3.403
Saucier DM, Shultz SR, Keller AJ, Cook CM, Binsted G. Sex differences in object location memory and spatial navigation in Long-Evans rats. Anim Cogn. 2008; 11:129–37. https://doi.org/10.1007/s10071-007-0096-1
Sovrano VA, Baratti G, Lee SA. The role of learning and environmental geometry in landmark-based spatial reorientation of fish (Xenotoca eiseni). PLoS ONE. 2020; 15(3):e0229608. https://doi.org/10.1371/journal.pone.0229608
Tolman EC. Cognitive maps in rats and men. Psychol Rev. 1948; 55(4):189. https://psycnet.apa.org/doi/10.1037/h0061626
Van der Bijl W, Thyselius M, Kotrschal A, Kolm N. Brain size affects the behavioural response to predators in female guppies (Poecilia reticulata). Proc Biol Sci. 2015; 282(1812):20151132. https://doi.org/10.1098/rspb.2015.1132
Vinepinsky E, Segev R. Neural mechanisms for spatial cognition across vertebrates. Mol Psychol. 2023; 2:1. https://doi.org/10.12688/molpsychol.17503.2
Wallace KJ, Rausch RT, Ramsey ME, Cummings ME. Sex differences in cognitive performance and style across domains in mosquitofish (Gambusia affinis). Anim Cogn. 2020; 23:655–69. https://doi.org/10.1007/s10071-020-01367-2
Warburton K. Learning of foraging skills by fish. Fish Fish. 2003; 4(3):203–15. https://doi.org/10.1046/j.1467-2979.2003.00125.x
Watson KK, Platt ML. Neuroethology of reward and decision making. Philos Trans R Soc Lond B Biol Sci. 2008; 363(1511):3825–35. https://doi.org/10.1098/rstb.2008.0159
White GE, Brown C. Cue choice and spatial learning ability are affected by habitat complexity in intertidal gobies. Behav Ecol. 2015; 26(1):178–84. https://doi.org/10.1093/beheco/aru178
Authors

![]() Kalebe S. Pinto1
Kalebe S. Pinto1 ![]() ,
, ![]() Jansen Zuanon1,2 and
Jansen Zuanon1,2 and ![]() Tiago H. S. Pires1
Tiago H. S. Pires1
[1] Programa de Pós-Graduação em Biologia de Água Doce e Pesca Interior, Instituto Nacional de Pesquisas da Amazônia, Av. André Araújo, 2936, Petrópolis, 69067-375, Manaus, AM, Brazil. (KSP) kalebedsp@gmail.com (corresponding author), (JZ) jzuanon3@gmail.com, (THSP) thspires@gmail.com.
[2] Universidade Santa Cecília, Rua Oswaldo Cruz, 277, Boqueirão, 11045-907, Santos, SP, Brazil (Senior Visiting Researcher).
Authors’ Contribution 

Kalebe S. Pinto: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing-original draft, Writing-review and editing.
Jansen Zuanon: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Tiago H. S. Pires: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing-original draft, Writing-review and editing.
Ethical Statement
Animal collection was conducted under SISBIO permit 10199–1, issued on October 16, 2007. This study was approved by the Ethics Committee for Animal Use (CEUA-INPA, protocol no 046/2016), in accordance with the guidelines of the Brazilian National Council for Animal Experimentation Control (CONCEA). After the experiments, most fish were released back at their collection site. Fish that were not returned to the habitat were kept in the laboratory for other experiments. No procedure was considered invasive or harmful to the fish, and no individuals died or were injured as a result of the experimental procedures.
Competing Interests
The author declares no competing interests.
Data availability statement
The datasets generated during the current study are available in the GitHub repository (https://github.com/kalebepinto/NI-2025-0160).
AI statement
Artificial intelligence (AI) tools (ChatGPT, OpenAI) were used to translate the manuscript from Portuguese to English and to assist with grammar correction.
Funding
Funding for this project was supported by the Japanese Science and Technology/Japan International Cooperation Agency (JST/JICA) Science and Technology Research Partnership (SATREPS), the Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM; FIXAM: 062.01500/2018), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Supplementary Material
Supplementary material SUP
How to cite this article
Pinto KS, Zuanon J, Pires THS. Sex differences in landmark-based spatial learning in the sailfin tetra Crenuchus spilurus (Characiformes: Crenuchidae). Neotrop Ichthyol. 2026; 24(2):e250160. https://doi.org/10.1590/1982-0224-2025-0160
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2025 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted January 26, 2026
Accepted January 26, 2026
![]() Submitted September 21, 2025
Submitted September 21, 2025
![]() Epub June 19, 2026
Epub June 19, 2026