![]() Antonio Augusto Jardim Jr1,2
Antonio Augusto Jardim Jr1,2 ![]() ,
, ![]() Erival Gonçalves Prata1,
Erival Gonçalves Prata1, ![]() Lais Martins Silva1,
Lais Martins Silva1, ![]() João Pedro da Silva Moraes1,
João Pedro da Silva Moraes1, ![]() Lucas Pires de Oliveira1,2,
Lucas Pires de Oliveira1,2, ![]() Luciano Fogaça de Assis Montag1,2 and
Luciano Fogaça de Assis Montag1,2 and ![]() Tiago Magalhães da Silva Freitas3
Tiago Magalhães da Silva Freitas3
PDF: Download Here | Supplementary: Sup | Cite this article
Associate Editor: ![]() Evelyn Habit
Evelyn Habit
Section Editor: ![]() Fernando Pelicice
Fernando Pelicice
Editor-in-chief: ![]() José Birindelli
José Birindelli
Abstract
Este estudo investiga a dinâmica trófica de Pinirampus pirinampu sob um regime de pulso de inundação alterado, utilizando análise de conteúdo estomacal e isótopos estáveis (δ13C e δ15N). Amostragens mensais foram realizadas no rio Xingu (Bacia Amazônica) de janeiro a novembro de 2021. No total, 135 estômagos (40 com conteúdo identificável) e 23 amostras de tecido muscular foram coletadas, juntamente com amostras representativas de potenciais fontes de alimento autóctones. A análise do conteúdo estomacal revelou uma dieta composta principalmente de insetos aquáticos, sem variação significativa na amplitude do nicho ou na intensidade alimentar ao longo dos períodos hidrológicos. No entanto, dados isotópicos indicaram que as principais fontes de carbono assimilado derivaram de crustáceos, peixes e moluscos, refletindo uma posição trófica consistentemente alta ao longo do ciclo hidrológico. Essas descobertas destacam a importância da integração de métodos complementares de avaliação trófica para obter uma compreensão mais abrangente da ecologia de peixes em condições de fluxo controlado. Além disso, os resultados fornecem insights para orientar estratégias de conservação e manejo que visem mitigar os impactos ecológicos dos empreendimentos hidrelétricos nas assembleias de peixes da Amazônia.
Palavras-chave: Ecologia trófica, Isótopos estáveis, Pimelodidae, Pinirampus pirinampu, Rio Xingu.
Introduction
The flood pulse is a seasonal natural phenomenon essential for the functioning of Neotropical River systems, driving ecological interactions between aquatic and terrestrial environments (Junk et al., 1989; Tockner et al., 2000). For fish, during high-water periods, floodplain inundation enhances connectivity among watercourses, enabling migration routes, providing predator refuges, and creating spawning and nursery areas (Goulding, 1980; Barbosa et al., 2018). This process also expands foraging opportunities by providing allochthonous resources such as terrestrial macroinvertebrates, fruits, and seeds, which mainly benefit frugivorous, insectivorous, omnivorous, and herbivorous species (Freitas, Garcez, 2004; Barbosa et al., 2018; Freitas et al., 2022).
In contrast, during low-water periods, the reduction in wetted areas increases the relative availability of autochthonous resources, including aquatic macroinvertebrates, zooplankton, phytoplankton, crustacean, mollusk, and macrophytes, which may become more accessible and easier to capture. This contraction of the main river channel also raises the density of aquatic organisms, favoring carnivorous species due to the greater aggregation of prey (Luz-Agostinho et al., 2009; Pereira et al., 2017; Barbosa et al., 2018).
Many studies have evaluated the effects of ecological processes such as the flood pulse on fish trophic ecology through direct analysis of stomach contents (Silveira et al., 2020; Freitas et al., 2022; Penha et al., 2024). This method involves the visual inspection of gastrointestinal contents, providing short-term information on diet composition that can be used to infer feeding intensity and trophic niche breadth, for instance (Amundsen, Sánchez-Hernández, 2019; Silveira et al., 2020). A complementary approach involves the analysis of stable carbon (δ13C) and nitrogen (δ15N) isotopes (Peterson, Fry, 1987; Fry, 2006; Davis et al., 2012). The δ13C values help trace the basal sources of organic matter assimilated by consumers, whereas δ15N provides estimates of a species’ trophic position within the food web (Albrecht et al., 2021; Winemiller et al., 2023). The isotopic data from these stable elements not only complement stomach content analyses but also offer integrated, long-term information about dietary assimilation, allowing the assessment of whether the items found in stomach contents correspond to those effectively assimilated (Prata et al., 2025; Seabra et al., 2025a).
Integrating stomach content and stable isotope analyses improves our understanding of how floodplain dynamics, particularly those impacted by anthropogenic disturbances, shape fish feeding behavior (Penha et al., 2024; Prata et al., 2025; Seabra et al., 2025a). Among such disturbances, hydropower development poses a significant threat to Neotropical fish assemblages (Fitzgerald et al., 2018), as it alters natural flow regimes and leads to habitat loss, environmental homogenization, and changes in sediment transport and water quality (Winemiller et al., 2016; Arantes et al., 2019).
A notable example is the Xingu River, a highly biodiverse tributary of the Amazon basin (Camargo et al., 2004; Barbosa et al., 2015). Following the commissioning of the Belo Monte Hydroelectric Dam in 2016 (Zuanon et al., 2019), the river’s natural discharge was reduced by approximately 80% downstream (Zuanon et al., 2019; Penha et al., 2024), potentially compromising ecological interactions with terrestrial environments during flood events (Winemiller et al., 2016). This new hydrological configuration may lead to changes in the composition of macroinvertebrate assemblages, an important food resource for many fish species (Fjellheim, Raddum, 1996; Richardson et al., 2002) and consequently alter fish diets in response to shifts in prey abundance and taxonomic composition. Moreover, interspecific competition and predation rates may increase if available food resources become limited, potentially leading to a higher frequency of empty stomachs (Baumgartner, 2007).
Among the diverse Neotropical fish fauna, the family Pimelodidae stands out, comprising approximately 30 genera and 117 species widely distributed throughout South America (Fricke et al., 2025). One of its representatives in the Xingu River is the catfish Pinirampus pirinampu (Spix & Agassiz, 1829), locally known as “barba-chata” (Dagosta, de Pinna, 2019). Siluriforms possess highly developed chemosensory and tactile systems that allow them to locate and capture prey even under low-light or turbid-water conditions (Barthem, Goulding, 1997; Freitas et al., 2020). These traits are particularly relevant for P. pirinampu, a medium-sized nocturnal piscivore that primarily consumes small fish and forages both in the water column and near the bottom (Barbarino Duque, Winemiller, 2003; Carolsfeld et al., 2003). This species also represents an important food resource for traditional local populations and is highly valued in some riverine communities (Valdelamar-Villegas, Olivero-Verbel, 2020). As a predatory catfish with access to a diverse prey base, P. pirinampu is likely influenced by fluctuations in floodplain connectivity and prey availability across hydrological periods, owing to its short-range lateral migration behavior (Lima et al., 2023).
Given this context, the present study aimed to investigate the trophic dynamics of Pinirampus pirinampu under the influence of the regulated flood pulse in the Xingu River. We hypothesized that (1) the contrast between high- and low-water periods under a regulated flow regime would not lead to major shifts in dietary composition, feeding intensity, or trophic niche breadth of P. pirinampu; (2) the isotopic structure (δ13C and δ15N) of the population would remain relatively stable between hydrological periods due to consistent availability of autochthonous resources; and (3) the contributions of assimilated basal sources, as well as isotopic niche area and overlap, would show little variation between periods, reflecting a stable trophic strategy. By identifying the stability of feeding patterns in a carnivorous predator under a regulated flow regime, this study provides insight into how hydrological control can influence energy pathways and trophic interactions in Amazonian River systems.
Material and methods
Study area. This study was conducted in the Reduced Flow Section (RFS) of the Xingu River, located in the Volta Grande region, Pará State, Brazil. The study area lies between the Pimental and Belo Monte dams and includes sampling sites corresponding represented by collection dates (Fig. 1). Since the construction of the Belo Monte Hydroelectric Complex in 2016, this section of the river has been subject to hydrological regulation, with flow regimes controlled according to environmental guidelines established by IBAMA and ANA, which define fixed limits for minimum and maximum discharge. Consequently, the alteration of the natural seasonal dynamics caused by dam construction has resulted in an approximately 80% reduction in downstream flow, significantly modifying the river’s hydrological and ecological processes (Fitzgerald et al., 2018; Zuanon et al., 2019; Bertassoli et al., 2021).
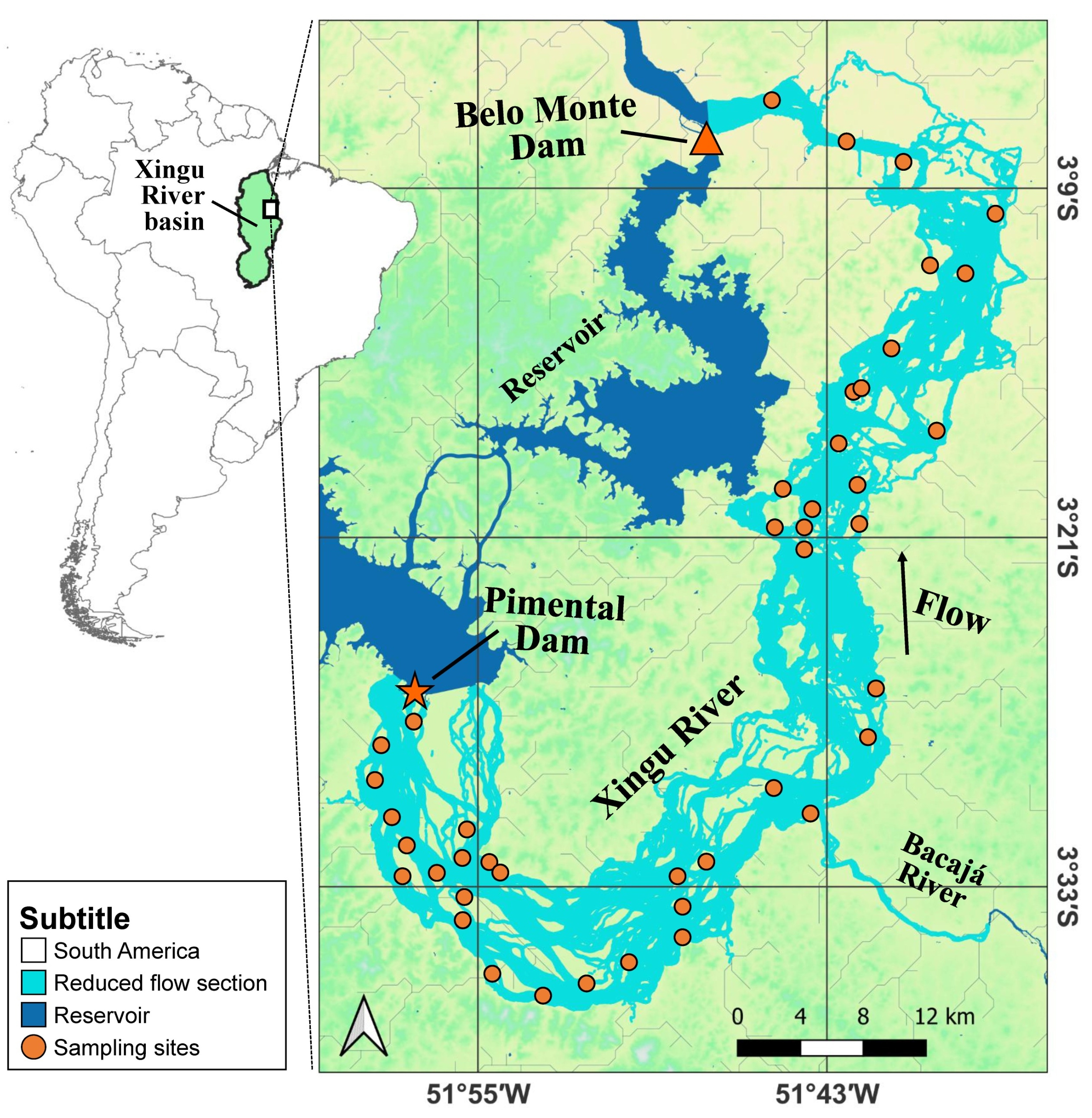
FIGURE 1| Sampling locations for Pinirampus pirinampu in the Volta Grande do Xingu (VGX), middle Xingu River (Pará, Brazil), from January to November 2021.
The region is classified as having a tropical monsoon climate (Am) according to the Köppen-Geiger system and is dominated by dense evergreen ombrophilous forest (Peel et al., 2007; Barbosa et al., 2018; Montag et al., 2025). The river channel in this section features clear waters, heterogeneous depth profiles, and a mosaic of rocky outcrops and sandy substrates, which together contribute to high habitat heterogeneity and complexity (Fitzgerald et al., 2018).
Flow data revealed pronounced seasonal variation, with peak flows of 8,215.9 m3/s in March (high-water season) and 707.8 m3/s in October (low-water season) (Penha et al., 2024). Based on this seasonal pattern, we defined two distinct hydrological phases: the high-water period (December to May) and the low-water period (June to November) (Freitas et al., 2022; Lima et al., 2023; Montag et al., 2025). These phases were used as categorical variables to assess the trophic ecology of Pinirampus pirinampu under regulated flow conditions.
Sampling procedures and processing biological material. Specimens were collected monthly from January to November 2021 using monofilament nylon gillnets with mesh sizes of 2, 8, 10, 12, 16, and 18 cm (measured between opposite knots). Each net measured 20 m in length and 1 m in height, and multiple nets were deployed sequentially to form standardized sampling sets. At each event, three gillnet sets were installed at sites spaced approximately 5 km apart. Captures from the three sets were pooled per sampling event for subsequent analyses. Nets were deployed at 16:00 and retrieved at 21:00 on the same day to ensure consistent sampling effort across sites and months.
After collection, a subset of individuals of Pinirampus pirinampu was selected, from which dorsal muscle tissue samples (without skin, scales, or spines) were collected from the right flank. In addition, tissue samples were obtained from individuals of other fish species, and other sources that could represent potential prey items were also sampled (see Tab. S1). Tissue samples were immediately stored on ice and later processed for stable isotope analysis.
Fish were anesthetized using benzocaine (0.1 g/L) and euthanized following the ethical guidelines of the Brazilian National Council for the Control of Animal Experimentation (CONCEA). Standard length (SL, in cm, to the nearest 0.1 cm) and total weight (TW, in g, to the nearest 0.01 g) were recorded for each specimen. Stomachs were removed via ventral incision, preserved in 70% ethanol, and sent to the Laboratory of Ecology and Conservation (LABECO) at the Universidade Federal do Pará (UFPA, Guamá Campus) for dietary analysis. Voucher specimens were fixed in 10% formalin for 48 h, transferred to 70% ethanol, and deposited in the ichthyological collection of the Zoological Museum at Universidade Federal do Pará (MZUFPA, Belém), under catalog number MZUFPA 891.
Samples of autochthonous sources (crustaceans and mollusks) were collected during each hydrological period. Crustaceans (shrimp and crab) and mollusks (bivalves) were captured through active search on rocks in rapids or rocky areas. Shrimps were kept in aquaria for 24 h to allow gut clearance; afterward, the samples were wrapped in aluminum foil and stored on ice.
All samples were rinsed with distilled water to remove inorganic debris and kept chilled to preserve their physicochemical integrity. Subsequent laboratory processing and stable isotope analysis were conducted at LABECO (UFPA), following standardized protocols to ensure analytical accuracy.
In the laboratory, stomach contents were examined under a stereomicroscope (up to 32× magnification). Food items were identified to the lowest possible taxonomic level using specialized literature (Buckup et al., 2007; Mugnai et al., 2010; Hamada et al., 2019) and with expert assistance. Each item was weighed on a high-precision analytical balance (± 0.0001 g) and assigned to one of six trophic categories: aquatic insects, aquatic crustaceans, aquatic mollusks, fish (autochthonous), plant fragments, and terrestrial insects (allochthonous).
For stable isotope analysis, all tissue and primary source samples were thawed, rinsed with distilled water, and placed in Petri dishes previously sterilized with a 10% diluted hydrochloric acid (HCl) solution for 24 h to remove inorganic carbonates. Samples were then oven-dried at 60 °C, with drying times varying by material type: fish muscle tissues, crustacean, and mollusk were dried for 24 h. Dried samples were ground into a fine powder using a porcelain mortar and pestle, stored in labeled Eppendorf tubes, and sent to the Center for Nuclear Energy in Agriculture (CENA) at the University of São Paulo (USP, Piracicaba) for isotopic analysis. At CENA, approximately 0.5 mg of each sample was encapsulated in ultra-pure tin capsules (4 × 6 mm), and stable isotope ratios of carbon (δ13C) and nitrogen (δ15N) were determined using an isotope ratio mass spectrometer (IRMS). All samples were analyzed in duplicate, and analytical precision, based on repeated measurements of internal laboratory standards, was better than ±0.2‰ for δ13C and ±0.3‰ for δ15N (Pinnegar, Polunin, 1999).
Data analysis. A total of 135 individuals of P. pirinampu were analyzed, with SL ranging from 11.8 to 58.5 cm (mean ± standard deviation = 37.30 ± 9.60 cm) and body weights from 17.85 to 64.60 g (11.13 ± 10.69 g). Of these, 95 had empty stomachs and were excluded. The remaining 40 individuals were analyzed for diet composition, trophic niche breadth, and feeding intensity. Based on the smallest length at first maturity regardless of sex (C50 = 53.6 cm; Peixer et al., 2006), most of the analyzed individuals were classified as juveniles, with only two adults (Fig. S2).
To quantify dietary composition, the weight (g) of each food item was converted to a percentage based on its proportion of total items per stomach (Hynes, 1950). The Frequency of Occurrence (FOi) (Hyslop, 1980) was also calculated to derive the Alimentary Index (Ai), using the modified formula proposed by Kawakami, Vazzoler (1980):
Ai= [(FOi x Mi) / Σ(FOi x Mi)] x 100
Where Ai represents the relative importance of food item i, FOi is its frequency, and Mi is its mass percentage. The Ai was calculated for grouped individuals, both across the entire study and per sampling date.
The Ai values of the trophic categories were square-root transformed and used in two main analyses. First, differences in diet composition between hydrological periods were assessed using Permutational Multivariate Analysis of Variance (PERMANOVA) via adonis2 function from the vegan package (Oksanen et al., 2024) in RStudio (R Development Core Team, 2024). Second, variation in trophic niche breadth was tested using Permutational Analysis of Multivariate Dispersion (PERMDISP), employing the vegdist and betadisper functions, both from the vegan package. This analysis quantifies the average distance of each sample to its group centroid, thereby comparing dispersion across the two hydrological phases. The unit of replication for both analyses was the pooled Ai values from individuals collected per sampling month per hydrological period.
Feeding intensity was evaluated using the Repletion Index (RI), calculated for each specimen according to the formula by Zavala-Camin (1996):
RI = (ME / MT) x 100
Where ME is the stomach content mass (including identifiable items and digested material) and MT is total body mass before dissection. Differences in RI values between phases were tested using a t-test (t.test function, stats package in RStudio). Normality and homogeneity of variances were verified using the Shapiro–Wilk and Levene’s tests. All statistical analyses, including the following tests, adopted α = 0.05 (Zar, 2010) and were conducted in RStudio (R Development Core Team, 2024).
Isotopic ratios of δ13C and δ15N were expressed in parts per thousand (‰) relative to international standards: Pee Dee Belemnite (PDB) for carbon and atmospheric nitrogen (AIR) for nitrogen, following the conventions established by Peterson, Fry (1987):
δ15C(‰) = [(13C / 12C)sample / (13C / 12C)standard -1] x 1000
and
δ15N(‰) = [(15N / 14N)sample / (15N / 14N)standard -1] x 1000
To correct lipid-related bias, we applied mathematical normalization using measured %C and C:N ratios as proxies for lipid content. For fish tissue with C:N ratios > 3.5, we applied the correction following Post et al. (2007):
∆δ13C = δ13Cextracted – δ13Cuntreated
To evaluate differences in isotopic composition between hydrological phases, independent t-tests were applied to δ13C and δ15N values. The isotopic niche area and overlap between hydrological phases were estimated using the Standard Ellipse Area corrected for small sample sizes (SEAc), expressed in ‰2. Twenty thousand iterations were performed, with 1,000 discarded as burn-in, a thinning factor of 10, and two independent chains, resulting in 3,800 posterior samples for each ellipse parameter. This Bayesian-based metric allows for robust estimation of trophic niche breadth and overlap in bivariate isotopic space (Jackson et al., 2011; Syväranta et al., 2013) and was implemented using the SIBER package (Stable Isotope Bayesian Ellipses in R) in RStudio. Isotopic niche overlap (IOv) was also calculated based on SEAc values (Jackson et al., 2011; Syväranta et al., 2013).
Before applying mixing models, we assessed source-consumer compatibility through simulations of the isotopic mixing space (Smith et al., 2013; Phillips et al., 2014). All tissue samples fell within the 95% mixing envelope, indicating no outliers and no need for sample exclusions (Fig. S3). Polygon simulations were performed using the sp (Pebesma et al., 2025) and splancs packages (Bivand et al., 2022) in RStudio.
Finally, to assess variations in the contribution of assimilated basal resources between hydrological phases, Bayesian isotopic mixing models were implemented using the simmr package (Parnell et al., 2010). The models were fitted using Dirichlet priors and Markov Chain Monte Carlo (MCMC) simulations. For each model, 10,000 iterations were run, with 10% discarded as burn-in. We applied the Gelman-Rubin and Geweke diagnostic tests, targeting convergence values equal to 1 as recommended by Parnell (2021). Ninety-five percent of credible intervals were used for result interpretation. To perform the mixing polygon and model analyses, we used the mean ± 1 SD of the trophic discrimination factor (TDFs): 0.47 ± 1.23‰ for δ13C and 2.92 ± 1.78‰ for δ15N, following Zanden and Rasmussen (2001), values previously applied in studies of Amazonian ichthyofauna (Penha et al., 2024, 2025; Prata et al., 2025; Seabra et al., 2025a,b).
Results
Diet composition. The diet of Pinirampus pirinampu (Tab. 1; Fig. 2) comprised 11 distinct food items, with larvae of aquatic insects representing the dominant component, accounting for 91% of the diet (Ai). When categorized by a trophic group, aquatic insects alone contributed 92% to the overall diet, reinforcing the insectivorous feeding habit of the species. All remaining food items combined contributed only 8% of the total diet (Tab. 1).
TABLE 1 | Alimentary index (Ai) of food origin, items, and category for the diet of Pinirampus pirinampu collected monthly and across hydrological periods in the middle Xingu River, Eastern Amazonia, Brazil. n = number of stomachs with food content.
Categories/Food items | 2021 | Total | ||||||||||
High-water | Low-water | |||||||||||
Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | n = 40 | |
n = 2 | n = 1 | n = 8 | n = 5 | n = 10 | n = 7 | n = 1 | n = 2 | n = 1 | n = 2 | n = 1 | ||
AUTOCHTHONOUS | ||||||||||||
Aquatic insect |
|
|
|
|
|
|
|
|
|
|
| 0.92 |
Aquatic insect fragment | 0.11 |
|
|
|
|
|
|
|
| 0.5 |
|
|
Aquatic insect (larva) | 0.89 |
| 1 | 0.87 | 0.98 | 0.56 | 1 | 1 | 1 | 0.53 | 1 |
|
Corydalidae (Immature) |
| 1 | <0.01 |
|
|
|
|
|
| 0.29 |
|
|
Aquatic crustacean |
|
|
|
|
|
|
|
|
|
|
| <0.01 |
Zooplankton (Cladocera) | 1 |
|
|
|
|
|
|
|
|
|
|
|
Aquatic mollusk |
|
|
|
|
|
|
|
|
|
|
| 0.01 |
Bivalve |
|
|
| 0.13 | 0.27 |
|
|
|
|
|
|
|
Fish |
|
|
|
|
|
|
|
|
|
|
| <0.01 |
Scale |
|
|
|
| 0.11 |
|
|
|
|
|
|
|
ALLOCHTHONOUS | ||||||||||||
Plant fragments |
|
|
|
|
|
|
|
|
|
|
| <0.01 |
Plant fragments |
|
|
| 0.93 |
|
|
|
|
|
|
|
|
Terrestrial insect |
|
|
|
|
|
|
|
|
|
|
| 0.07 |
Blattodea (termite) |
|
| 0.28 |
|
| 0.44 |
|
|
| 0.29 |
|
|
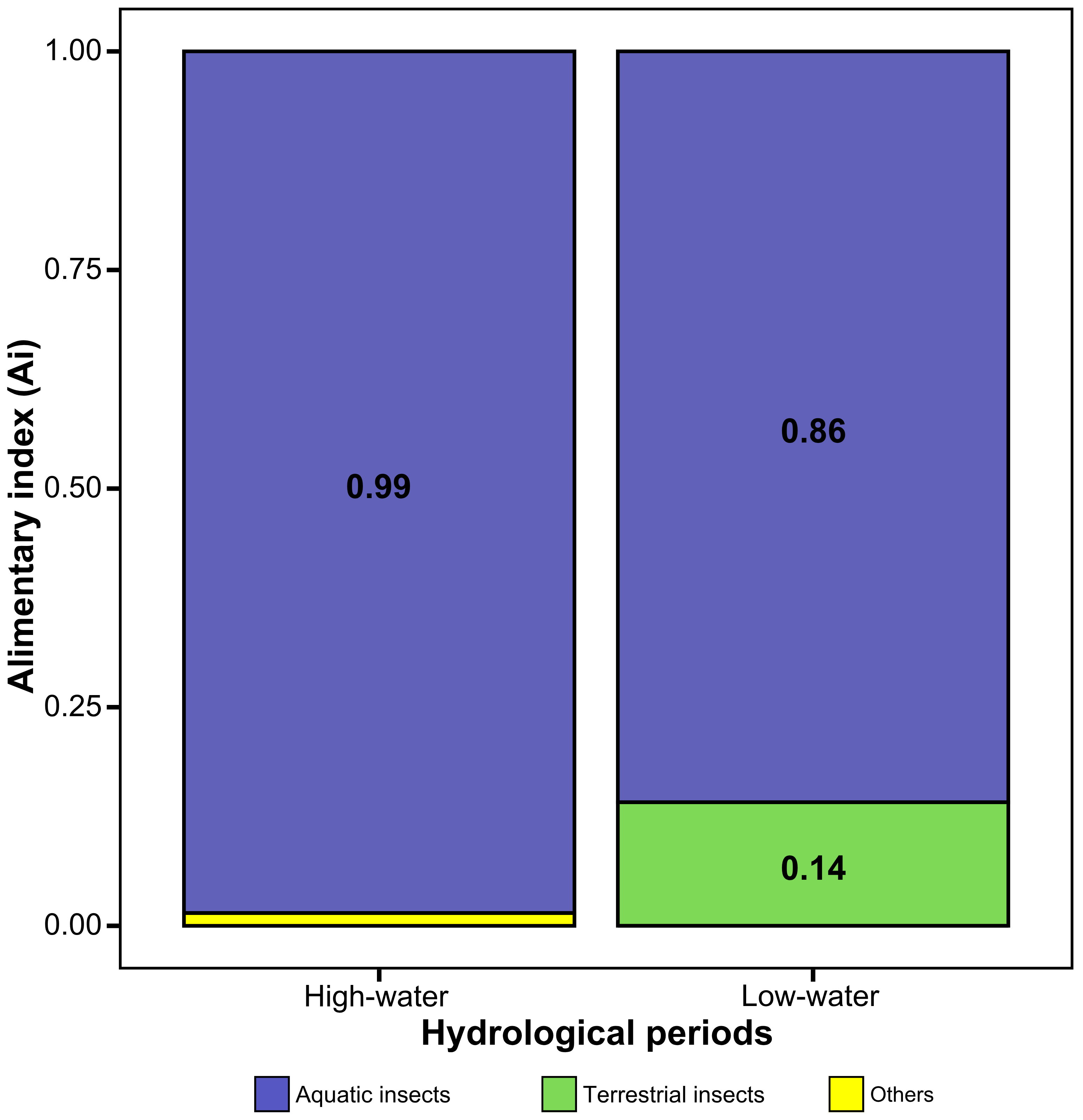
FIGURE 2| Alimentary index (Ai) of the diet of Pinirampus pirinampu collected across hydrological phases in the middle Xingu River, Eastern Amazonia, Brazil.
Food consumption varied among months and between high- and low-water periods. During the high-water period, overall food consumption was higher, with pronounced peaks in May for aquatic insect fragments and bivalves. In the low-water period (June), there was a higher consumption of Blattodea (termites) (Fig. S4).
No significant differences in diet composition were detected between high- and low-water periods (PERMANOVA; Pseudo-F = 1.052, p = 0.35), nor in trophic niche breadth (PERMDISP; F = 0.64, p = 0.43), indicating dietary consistency across hydrological phases. Likewise, feeding intensity, as measured by the Repletion Index (RI), showed no significant variation between periods (t = 0.08, df = 37.363, p = 0.93; n = 26 for the high-water period and n = 14 for the low-water period), suggesting that P. pirinampu maintains a similar feeding activity throughout the hydrological cycle.
Stable isotopes. A total of 824 samples were analyzed for stable isotope composition. Of these, 23 were muscle tissue samples from P. pirinampu, with 11 collected during the high-water period and 12 during the low-water period. The remaining 801 samples represented basal resources, including fish, crustaceans, and mollusks (Tab. 2).
TABLE 2 | Number of samples, means, and standard deviations (±SD) of δ13C (‰) and δ15N (‰) values for Pinirampus pirinampu and basal resources collected across hydrological periods in the middle Xingu River, Eastern Amazonia, Brazil. n = number of tissues sampled.
Species/Resource | High-water | Low-water | ||||||||
Mean | SD | n | Mean | SD | n | |||||
δ13C (‰) | δ15N (‰) | δ13C (‰) | δ15N (‰) | δ13C (‰) | δ15N (‰) | δ13C (‰) | δ15N (‰) | |||
Pinirampus pirinampu | -28 | 12.58 | 1.13 | 0.56 | 11 | -29.1 | 12.5 | 1.12 | 0.35 | 12 |
Fish | -27.3 | 10.59 | 2.91 | 1.36 | 359 | -27.8 | 10.57 | 2.69 | 1.26 | 344 |
Crustacean | -28.8 | 9.9 | 2.11 | 0.46 | 17 | -29.2 | 9.9 | 1.99 | 1.13 | 35 |
Mollusk | -25.8 | 7.36 | 4.66 | 1.04 | 10 | -29 | 7.13 | 3.93 | 1.11 | 36 |
Total |
|
|
|
| 397 |
|
|
|
| 427 |
Mean δ13C values of P. pirinampu differed significantly between hydrological periods (t = 2.210; df = 20.778; p = 0.038; Fig. 3), indicating a shift in the assimilated carbon source. Individuals sampled during the low-water period exhibited more depleted δ13C values (–29.1‰), while those from the high-water period showed less depleted values (–28‰), resulting in an absolute difference of approximately 1‰ (∆δ13C ≈ 1‰). Conversely, δ15N values remained stable across periods (t = 0.363; df = 16.586; p = 0.72), averaging 12.58 ± 0.56‰ during the high-water period and 12.50 ± 0.35‰ during the low-water period (Tab. 2), suggesting no significant change in trophic position.
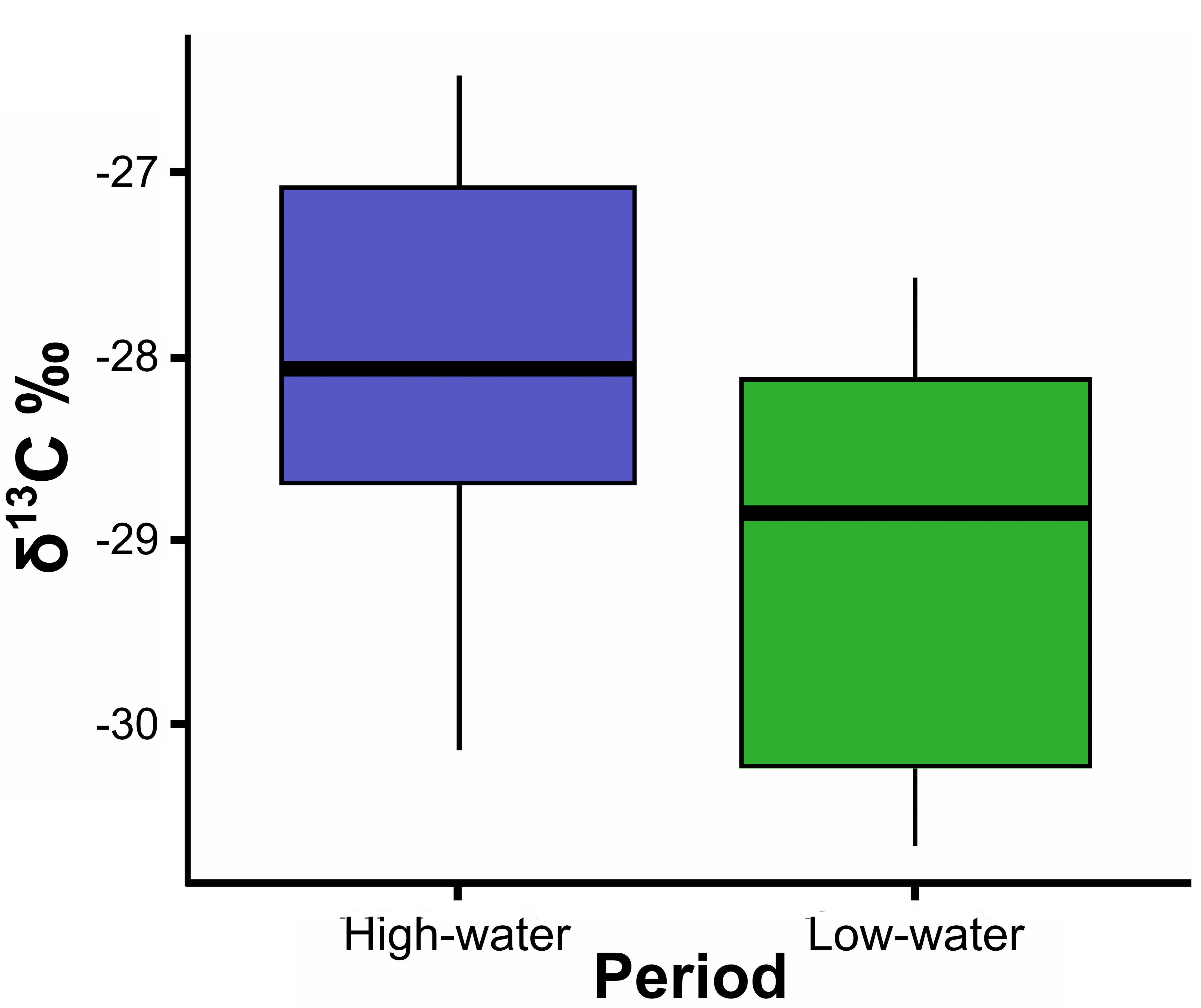
FIGURE 3| Boxplots of δ13C values for Pinirampus pirinampu between hydrological phases in the reduced flow stretch in the middle Xingu River. The median is represented as a horizontal line within the box; box limits represent the first and third quartiles; whiskers indicate minimum and maximum values; dots indicate outliers. Mean δ15N values did not show a statistically significant difference.
The relative contributions of assimilated carbon sources by P. pirinampu varied between hydrological periods, with crustaceans as the primary contributors (Fig. 4). Values represent posterior means from Bayesian mixing models implemented in MixSIAR. During the high-water period, crustaceans accounted for 53%, followed by fish (29%), while mollusk contributed 18%. During the low-water period, the contribution of crustaceans (45%) decreased slightly, fish remained relatively stable (31%), and mollusks increased (24%).
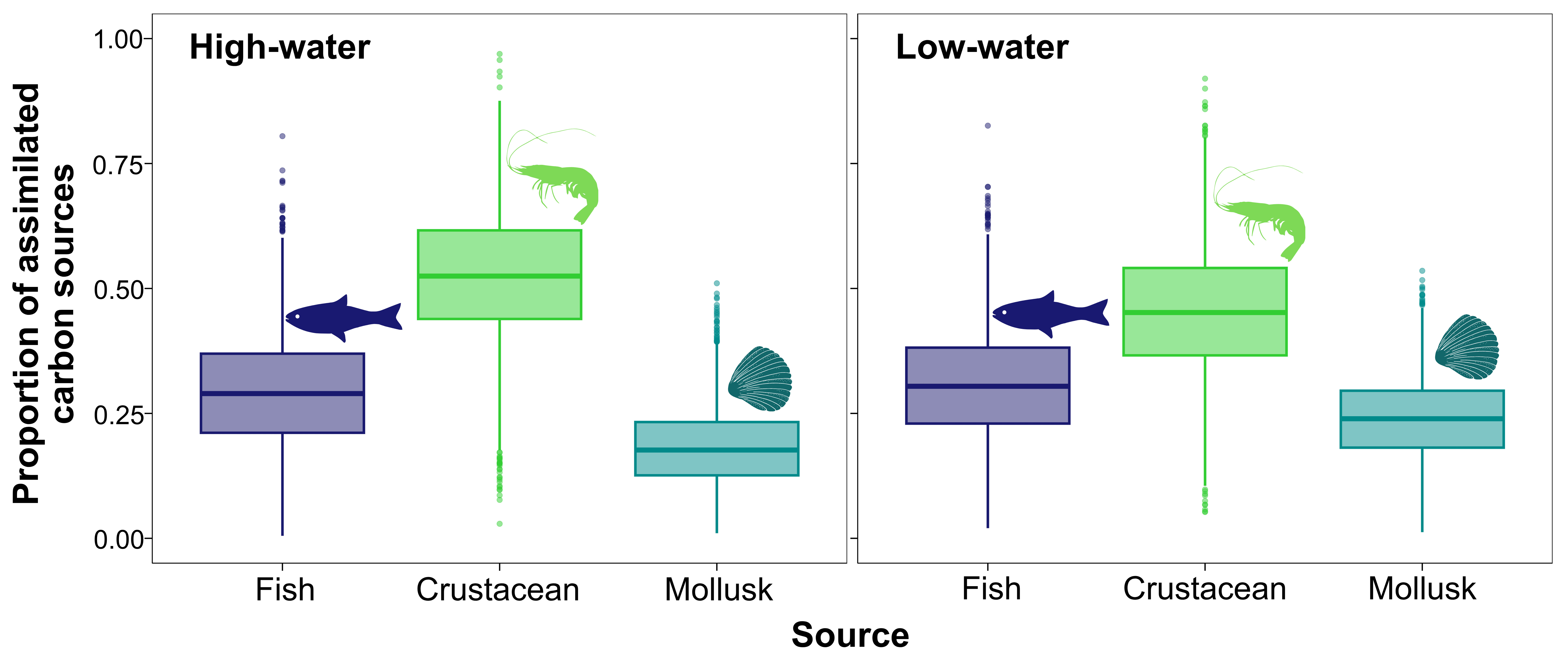
FIGURE 4| Proportional contributions of assimilated basal resources by Pinirampus pirinampu across hydrological phases in the reduced flow stretch of the middle Xingu River, Eastern Amazonia, Brazil.
Mixing model analyses revealed minimal changes in the relative assimilation of carbon sources between hydrological phases (Fig. 5). In both periods, crustaceans were the dominant sources, followed by fish, with mollusks contributing to a lesser extent.
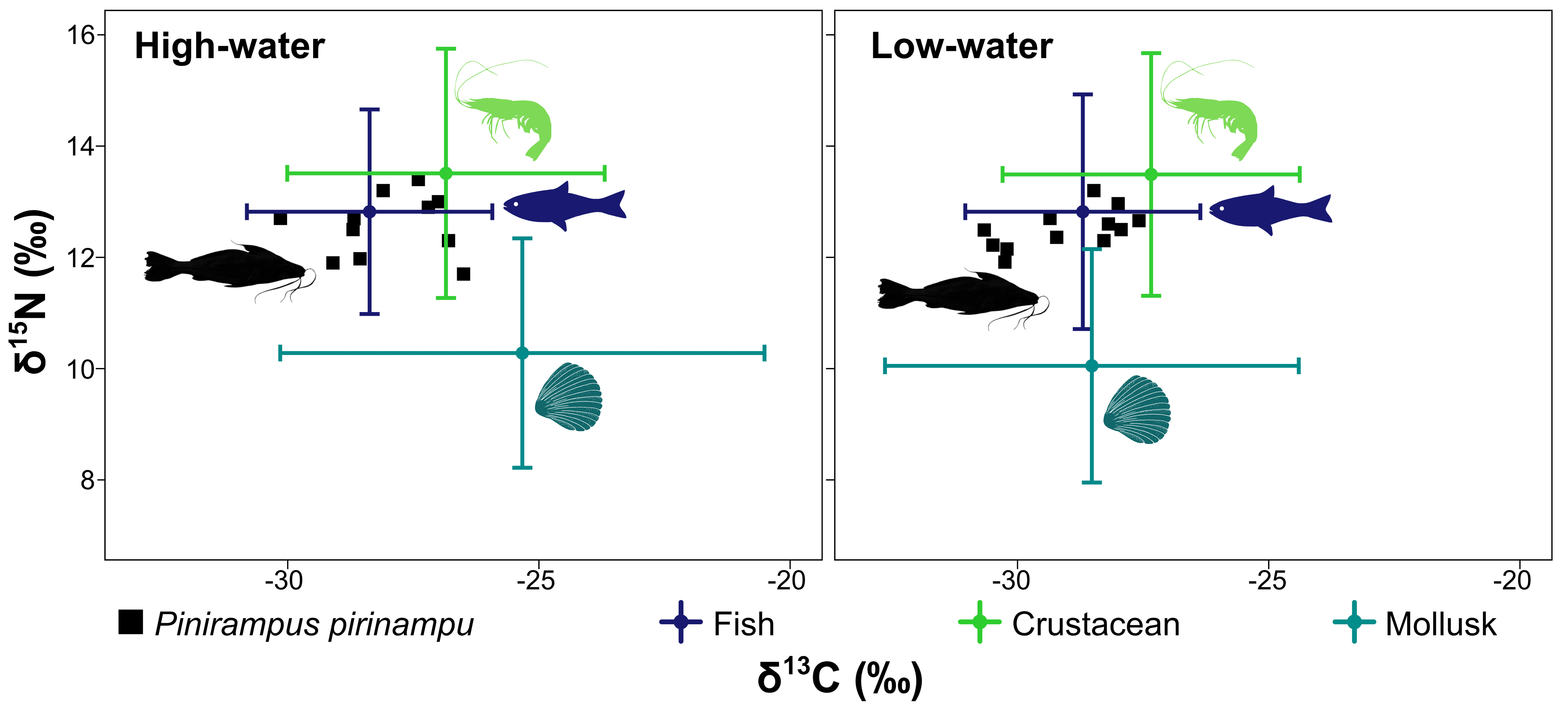
FIGURE 5| Mix model outputs for Pinirampus pirinampu and associated resources collected from January to November 2021 in the Volta Grande do Xingu, Pará, Brazil. Trophic Discrimination Factors (TDF): 0.47 ± 1.23 for δ13C and 2.92 ± 1.78 for δ15N (Zanden, Rasmussen, 2001).
We observed variation in isotopic niche areas between hydrological periods. The largest Standard Ellipse Area corrected for small sample size (SEAc) occurred during the high-water period (2.22‰2), while the low-water period exhibited a reduced isotopic niche area (1.17‰2) (Fig. 6). The overlap between periods, calculated using SEAc-based ellipses in SIBER, was 0.65‰2, representing 19.13% of the total combined isotopic niche area.
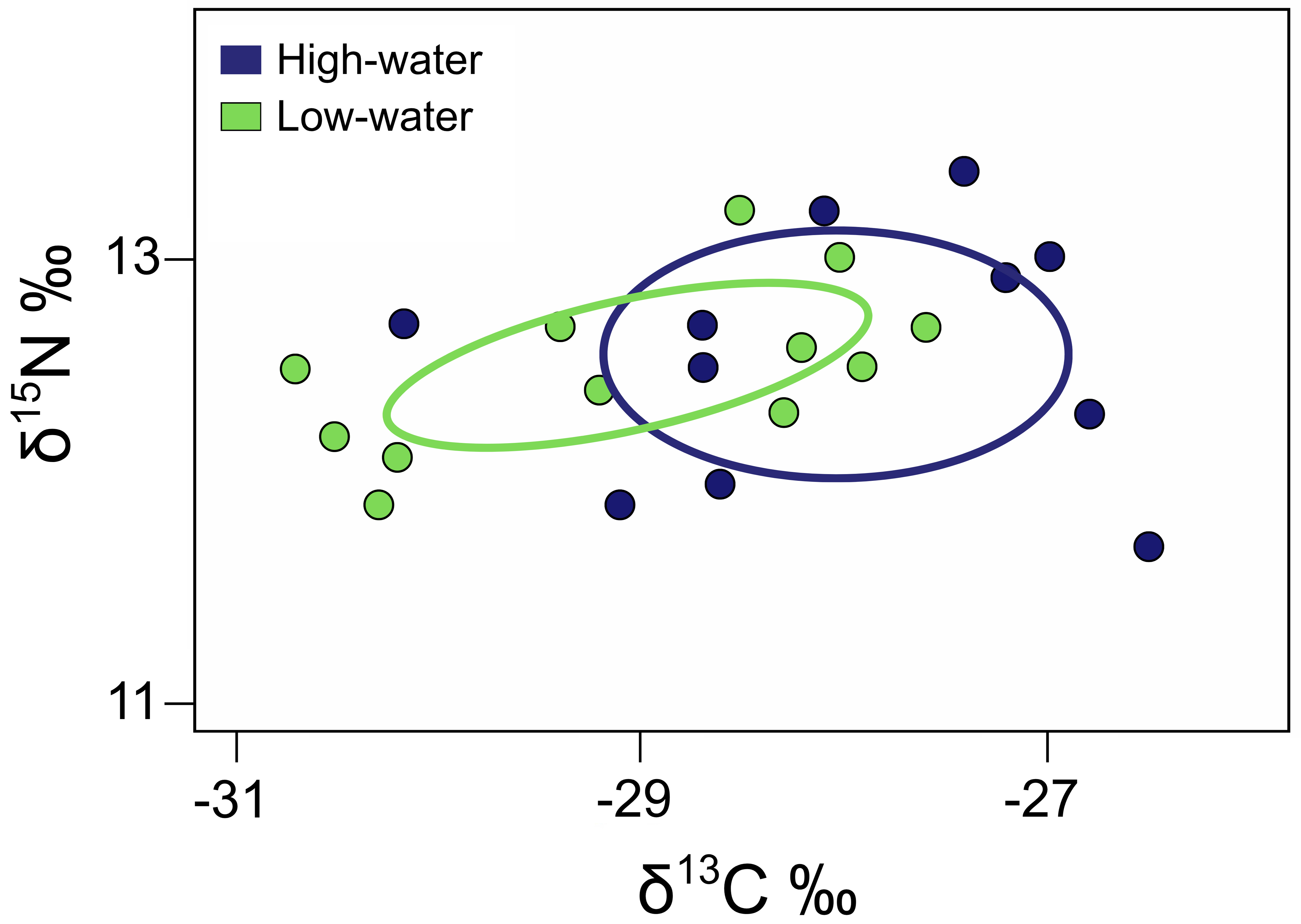
FIGURE 6| Isotopic niche area of Pinirampus pirinampu across hydrological phases in the reduced stretch flow in the middle Xingu River. Solid lines represent SEAc (40% credibility interval). The overlap between periods representing 19.13% of the total combined isotopic niche area.
Discussion
Our results demonstrate that Pinirampus pirinampu maintains a consistent insectivorous feeding strategy throughout the high- and low-water periods. No significant differences were observed in diet composition, trophic niche breadth, or feeding intensity between periods, supporting the initial hypothesis that variations in water volume would not markedly alter the species’ feeding parameters. Despite the relatively small number of stomachs analyzed, the patterns observed were statistically coherent across individuals and periods, which could be further confirmed by future studies with larger sample sizes.
In contrast, the hypothesis regarding isotopic stability was only partially supported. While δ15N values remained stable between the high- and low-water seasons, indicating a consistent trophic position and sustained assimilation of prey from higher levels of the food web, δ13C values varied significantly between periods. Although the absolute difference in δ13C values was modest (≈1‰), such variation can still be ecologically meaningful in Amazonian floodplain systems, where even small isotopic shifts may reflect changes in the balance between allochthonous and autochthonous carbon inputs. In this context, the observed pattern likely reflects seasonal adjustments in assimilated carbon sources, driven by changes in prey use in response to fluctuations in the relative availability of fish, crustaceans, and mollusks. Additionally, the broader trophic niche space (inferred from the SEAc) observed during the high-water period, together with moderate isotopic niche overlap (19.13%), suggests that the species can exploit a wider range of carbon sources when connectivity and habitat diversity increase during floods.
Carnivorous fish are known to adjust their diets according to prey availability, as documented in studies across the Amazon basin (Moreira-Hara et al., 2009; Prata et al., 2025). In the study region, crustaceans, fish (likely juveniles), and aquatic insects represented the most abundant and accessible prey items. Supporting this, Prata et al. (2025), working concurrently in the same area, also reported that these groups dominated the diet of another carnivorous fish species. Similar to our findings for P. pirinampu, that species exhibited limited seasonal dietary variation, suggesting that high prey availability and broad foraging flexibility contribute to the trophic stability of carnivorous fishes in floodplain environments.
Despite the relatively high number of individuals captured, the high proportion of empty stomachs observed in P. pirinampu can be considered a common feature among carnivorous fishes (Freitas et al., 2020). This pattern is often attributed to intermittent feeding behavior and the rapid digestion of soft-bodied prey, both of which reduce the likelihood of detecting food remains (Manko, 2016; Jiao et al., 2023; Skjoldal et al., 2025). In addition, factors such as capture-related stress, seasonal variation in prey availability, and the low feeding frequency of larger individuals may also contribute to this outcome (Almeida et al., 1997; Vignon, Dierking, 2011; Gupta, Banerjee, 2014; Ferreira et al., 2024).
Although P. pirinampu has traditionally been described as a piscivorous species (Barbarino Duque, Winemiller, 2003; Bennemann et al., 2011; Röpke et al., 2025), our results revealed a predominance of aquatic insects in its diet, with fish and other prey groups occurring less frequently. Given that most of the analyzed individuals were juveniles (Peixer et al., 2006), this feeding pattern likely reflects ontogenetic effects, in which greater trophic plasticity and the consumption of small, abundant prey constitute an energetically efficient foraging strategy (Almeida et al., 1997; Qin et al., 2024; Skjoldal et al., 2025). Thus, the predominance of autochthonous benthic resources such as aquatic insects suggests that juvenile P. pirinampu intensively exploit shallow margins and substrates during the high-water period, when prey availability increases substantially (Duarte et al., 2019; Brito et al., 2021; Röpke et al., 2025).
Our findings suggest that the trophic strategy of P. pirinampu aligns with the Optimal Foraging Theory, which posits that predators maximize their net energy intake by targeting prey that are both abundant and energetically efficient to capture and handle (MacArthur, Pianka, 1966; Stephens, Krebs, 1986). The predominance of crustaceans in assimilated resources, which are generally easier to capture than juvenile fish (Carvalho et al., 2006; Carvalho et al., 2020), and the significant consumption of aquatic insects during the high-water period, as discussed above, support this interpretation. Therefore, P. pirinampu exhibits a foraging strategy adapted to local environmental conditions, prioritizing prey that are both abundant and easily captured.
Discrepancies between stomach content analysis and stable isotope data observed in this study are not uncommon and have been documented in other predatory fishes worldwide (Beaudoin et al., 1999; Jensen et al., 2012), where ingestion of invertebrates was common but isotopic signatures reflected assimilation of higher-trophic-level prey (e.g., fish). Conversely, an opposite pattern has been reported, where stable isotope analysis of a previously known piscivorous species revealed aquatic invertebrates’ signatures, resulting in a lower trophic position (Gerber et al. 2023). This discrepancy likely reflects the different temporal scales of both methods: stomach content analysis represents recent feeding, whereas isotopes integrate assimilated diet over weeks or months (Pinnegar, Polunin, 1999; Post, 2002). In our study, stomach contents indicated a mainly insectivorous diet, while isotopes suggested greater assimilation of crustacean-derived carbon, especially during high water, highlighting the complementary nature of both approaches.
The broader isotopic niche observed during the high-water period likely reflects increased food availability and access to expanded habitat, including flooded forests. This pattern is consistent with the Flood Pulse Concept (Junk et al., 1989), which predicts greater trophic generalism during floods due to higher resource heterogeneity and increased foraging opportunities (Winemiller, 1996; Azevedo et al., 2022). Even under an altered flood regime, the seasonal rise in water levels still appears sufficient to expand the diversity of assimilated carbon sources.
Moreover, the moderate overlap observed between isotopic niches, together with the broader niche area during the high-water period, supports the interpretation of dietary plasticity, allowing P. pirinampu to adjust to seasonal shifts while maintaining reliance on core prey groups. Such trophic flexibility is a common adaptive strategy among predatory fishes (Barbosa et al., 2018; Weiss et al., 2025) and may be essential for species persistence in environments subjected to hydrological alterations.
It is also important to note that, following the reduction of flood pulses after the construction of the Belo Monte Hydroelectric Dam in 2016, monitoring in the Volta Grande do Xingu has revealed changes in the spatial dynamics of organic matter deposition (Utsunomiya et al., 2024; Quaresma et al., 2025). Such altered hydrological regimes are known to reduce the biomass of marginal and benthic biomass of macroinvertebrates and to disrupt both the timing and extent of allochthonous and autochthonous prey inputs into adjacent habitats, consequently affecting prey availability for juvenile foragers (Baumgartner, 2007; Pereira et al., 2017). The feeding data obtained here may reflect subtle ecological shifts associated with these hydrological changes, indicating variations in benthic productivity and the use of nearshore resources (Vinson, Angradi, 2011; Baker et al., 2014; Han et al., 2020; Ng et al., 2021; Poiesz et al., 2024). Reservoir impoundment and flow regulation often simplify riparian vegetation and benthic communities, altering the distribution and abundance of insect prey (Fjellheim, Raddum, 1996; Richardson et al., 2002), factors that have been shown to drive changes in the diets and trophic niches of predatory fishes in regulated rivers (Richardson et al., 2002).
In summary, our results revealed a distinct trophic dynamic of P. pirinampu, a carnivorous species influenced by the hydrological regulation imposed by the Belo Monte Dam. Seasonal variations in assimilated carbon sources suggest flexible foraging behavior in response to altered prey availability, reflecting the feeding plasticity that supports the species’ resilience and the maintenance of its functional role within the food web. However, the apparent stability in the feeding dynamics (e.g., stomach contents) may also represent an adaptive response to ongoing environmental changes rather than an absence of impact. Such adjustments underscore the importance of long-term monitoring and the integration of complementary trophic approaches to better assess the environmental consequences of flow regulation in Amazonian rivers.
Acknowledgments
LMS and JPSM thank BRC Norsk Hydro for their undergraduate research scholarship, and the ichthyology team of the Laboratório de Ecologia e Conservação at the Universidade Federal do Pará for their support.
References
Albrecht MP, Reis AS, Neres-Lima V, Zandonà E. Isótopos estáveis e outras ferramentas em estudos tróficos de peixes em riachos tropicais. Oecol Aust. 2021; 25(2):283–300. https://doi.org/10.4257/oeco.2021.2502.05
Almeida VLL, Hahn NS, Vazzoler AEAM. Feeding patterns in five predatory fishes of the high Paraná River floodplain (PR, Brazil). Ecol Freshw Fish. 1997; 6(3):123–33. https://doi.org/10.1111/j.1600-0633.1997.tb00154.x
Amundsen PA, Sánchez-Hernández J. Feeding studies take guts–critical review and recommendations of methods for stomach contents analysis in fish. J Fish Biol. 2019; 95(6):1364–73. https://doi.org/10.1111/jfb.14151
Arantes CC, Fitzgerald DB, Hoeinghaus DJ, Winemiller KO. Impacts of hydroelectric dams on fishes and fisheries in tropical rivers through the lens of functional traits. Curr Opin Environ Sustain. 2019; 37:28–40. https://doi.org/10.1016/j.cosust.2019.04.009
Azevedo LS, Pestana IA, Almeida MG, Bastos WR, Souza CMM. Do fish isotopic niches change in an Amazon floodplain lake over the hydrological regime? Ecol Freshw Fish. 2022; 31(1):72–80. https://doi.org/10.1111/eff.12609
Baker R, Buckland A, Sheaves M. Fish gut content analysis: robust measures of diet composition. Fish Fish. 2014; 15(1):170–77. https://doi.org/10.1111/faf.12026
Barbarino Duque A, Winemiller KO. Dietary segregation among large catfishes of the Apure and Arauca Rivers, Venezuela. J Fish Biol. 2003; 63(2):410–27. http://dx.doi.org/10.1046/j.1095-8649.2003.00163.x
Barbosa TAP, Barthem RB, Montag LFA. Feeding ecology of immature Lithodoras dorsalis (Valenciennes, 1840) (Siluriformes: Doradidae) in a tidal environment, estuary of the rio Amazonas. Neotrop Ichthyol. 2015; 13(2):341–48. https://doi.org/10.1590/1982-0224-20130014
Barbosa TAP, Rosa DC, Soares BE, Costa CH, Esposito MC, Montag LFA. Effect of flood pulses on the trophic ecology of four piscivorous fishes from the eastern Amazon. J Fish Biol. 2018; 93(1):30–39. https://doi.org/10.1111/jfb.13669
Barthem R, Goulding M. The catfish connection: ecology, migration, and conservation of amazon predators. New York. Columbia University Press; 1997.
Baumgartner LJ. Diet and feeding habits of predatory fishes upstream and downstream of a low-level weir. J Fish Biol. 2007; 70(3):879–94. https://doi.org/10.1111/j.1095-8649.2007.01352.x
Beaudoin CP, Tonn WM, Prepas EE, Wassenaar LI. Individual specialization and trophic adaptability of northern pike Esox lucius: an isotope and dietary analysis. Oecologia. 1999; 120:386–96. https://doi.org/10.1007/s004420050871
Bennemann ST, Galves W, Capra LG. Recursos alimentares utilizados pelos peixes e estrutura trófica de quatro trechos no reservatório Capivara Rio Paranapanema. Biota Neotrop. 2011; 11(1):63–71. https://doi.org/10.1590/S1676-06032011000100006
Bertassoli Jr. DJ, Sawakuchi HO, Araújo KR, Camargo MG, Alem VA, Pereira TS et al. How green can Amazon hydropower be? Net carbon emission from the largest hydropower plant in Amazonia. Sci Adv. 2021; 7(26):eabe1470. https://doi.org/10.1126/sciadv.abe1470
Bivand R, Rowlingson B, Diggle P, Petris G, Eglen S. Splancs: spatial and space-time point pattern analysis. J Geogr Syst. 2022; 2:307–17. https://doi.org/10.32614/CRAN.package.splancs
Brito JS, Michelan TS, Juen L. Aquatic macrophytes are important substrates for Libellulidae Odonata larvae and adults. Limnology. 2021; 22:139–49. https://doi.org/10.1007/s10201-020-00643-x
Buckup PA. Catálogo das espécies de peixes de água doce do Brasil. In: Buckup PA, Menezes NA, Ghazzi MSA, editors. Rio de Janeiro: Museu Nacional; 2007
Camargo M, Giarrizzo T, Isaac V. Review of the geographic distribution of fish fauna of the Xingu river basin, Brazil. Ecotropica. 2004; 10(2):123–47.
Carolsfeld J, Harvey B, Ross C, Baer A. Migratory fishes of South America. Ottawa: World Fisheries Trust/The World Bank. International Development Research Centre; 2003.
Carvalho FL, Magalhães C, Mantelatto FL. A molecular and morphological approach on the taxonomic status of the Brazilian species of Palaemon Decapoda Palaemonidae. Zool Scr. 2020; 49(1):101–16. https://doi.org/10.1111/zsc.12394
Carvalho LN, Zuanon J, Sazima I. The almost invisible league: crypsis and association between minute fishes and shrimps as a possible defence against visually hunting predators. Neotrop Ichthyol. 2006; 4(2):219–24. https://doi.org/10.1590/S1679-62252006000200008
Dagosta FCP, de Pinna M. The fishes of the Amazon: distribution and biogeographical patterns, with a comprehensive list of species. Bull Am Mus Nat Hist. 2019; 2019(431):1–163. https://doi.org/10.1206/0003-0090.431.1.1
Davis AM, Blanchette ML, Pusey BJ, Jardine TD, Pearson RG. Gut content and stable isotope analyses provide complementary understanding of ontogenetic dietary shifts and trophic relationships among fishes in a tropical river. Freshw Biol. 2012; 57(10):2156–72. https://doi.org/10.1111/j.1365-2427.2012.02858.x
Duarte C, Magurran AE, Zuanon J, Deus CP. Trophic ecology of benthic fish assemblages in a lowland river in the Brazilian Amazon. Aquat Ecol. 2019; 53(4):707–18. https://doi.org/10.1007/s10452-019-09720-5
Ferreira VMB, Soares BE, Leal JS, Albrecht MP. Spatial factors overcome seasonality regulating the consumption of allochthonous food resources by fishes from tropical lotic ecosystems. Freshw Biol. 2024; 69(6):783–91. https://doi.org/10.1111/fwb.14245
Fitzgerald DB, Sabaj Perez MH, Sousa LM, Gonçalves AP, Rapp Py-Daniel L, Lujan NK et al. Diversity and community structure of rapids-dwelling fishes of the Xingu River: implications for conservation amid large-scale hydroelectric development. Biol Conserv. 2018; 222:104–12. https://doi.org/10.1016/j.biocon.2018.04.002
Fjellheim A, Raddum GG. Weir building in a regulated west Norwegian river: long-term dynamics of invertebrates and fish. River Res Applic. 1996; 12(4-5):501–08. https://doi.org/10.1002/(SICI)1099-1646(199607)12:4/5%3C501::AID-RRR414%3E3.0.CO;2-F
Freitas CEC, Garcez RCS. Fish communities of natural channels between floodplain lakes and Solimoes-Amazonas River Amazon-Brazil. Acta Limnol Bras. 2004; 16(3):273–80.
Freitas TMS, Prudente BDS, Montag LFA. Flood pulse influence on the feeding ecology of two Amazonian auchenipterid catfishes. Neotrop Ichthyol. 2022; 20(1):e210103. https://doi.org/10.1590/1982-0224-2021-0103
Freitas TMS, Santos WO, Prudente BDS, Montag LFA. Diet and foraging behavior of Ageneiosus inermis (Teleostei, Auchenipteridae). Neotrop Biol Conserv. 2020; 15(3):209–18. https://doi.org/10.3897/neotropical.15.e53383
Fricke R, Eschmeyer WN, Fong JD. Eschmeyer’s catalog of fishes: genera/species by family/subfamily [Internet]. San Francisco: California Academy of Sciences; 2025. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp
Fry B. Stable isotope ecology. New York: Springer; 2006. https://doi.org/10.1007/0-387-33745-8
Gerber R, Necker L, van Vuren JH, Ikenaka Y, Nakayama SM, Ishizuka M et al. A stable isotope analysis of the dietary patterns of the aquatic apex predator, the African tigerfish (Hydrocynus vittatus). J Fish Biol. 2023; 103(5):1129–43. https://doi.org/10.1111/jfb.15516
Gupta S, Banerjee S. Food and feeding habit of a freshwater catfish, Mystus tengara (Siluriformes: Bagridae). J Ichthyol. 2014; 54(9):742–48. https://doi.org/10.1134/S0032945214060071
Goulding M. The fishes and the forest: explorations in Amazonian natural history. Oakland: University of California Press; 1980.
Hamada N, Nessimian J, Querino R. Insetos aquáticos na Amazônia brasileira: taxonomia, biologia e ecologia. Manaus: Editora INPA; 2019
Han D, Zhang C, Xue Y, Xu B, Ren Y, Chen Y. Impacts of sample size for stomach content analysis on the estimation of ecosystem indices. Acta Oceanol Sin. 2020; 39(8):53–61. https://doi.org/10.1007/s13131-020-1633-x
Hynes HBN. The food of fresh-water sticklebacks Gasterosteus aculeatus and Pygosteus pungitius, with a review of methods used in studies of the food of fishes. J Anim Ecol. 1950; 19(1):36–58. https://doi.org/10.2307/1570
Hyslop EJ. Stomach contents analysis, a review of methods and their application. J Fish Biol. 1980; 17(4):411–29. https://doi.org/10.1111/j.1095-8649.1980.tb02775.x
Jackson AL, Inger R, Parnell AC, Bearhop S. Comparing isotopic niche widths among and within communities: SIBER – Stable Isotope Bayesian Ellipses in R. J Anim Ecol. 2011; 80(3):595–602. https://doi.org/10.1111/j.1365-2656.2011.01806.x
Jensen H, Kiljunen M, Amundsen PA. Dietary ontogeny and niche shift to piscivory in lacustrine brown trout Salmo trutta revealed by stomach content and stable isotope analyses. J Fish Biol. 2012; 80(7):2448–62. https://doi.org/10.1111/j.1095-8649.2012.03294.x
Jiao F, Zhang L, Limbu SM, Yin H, Xie Y, Yang Z et al. A comparison of digestive strategies for fishes with different feeding habits: digestive enzyme activities, intestinal morphology, and gut microbiota. Ecol Evol. 2023; 13(9):e10499. https://doi.org/10.1002/ece3.10499
Junk WJ, Bayley PB, Sparks RE. The flood pulse concept in river-floodplain systems. Can Spec Publ Fish Aquat Sci. 1989; 106(1):110–27.
Kawakami E, Vazzoler G. Método gráfico e estimativa de índice alimentar aplicado no estudo de alimentação de peixes. Bol Inst Oceanogr. 1980; 29(2):205–07. https://doi.org/10.1590/S0373-55241980000200043
Lima MS, Martins SS, Montag LFA, Freitas TMS. Dietary shift of a pimelodid catfish in response to the flood pulse in the Xingu River. Neotrop Ichthyol. 2023; 21(4):e230097. https://doi.org/10.1590/1982-0224-2023-0097
Luz-Agostinho KD, Agostinho AA, Gomes LC, Fugi R. Effects of flooding regime on the feeding activity and body condition of piscivorous fish in the Upper Paraná River floodplain. Braz J Biol. 2009; 69:481–90. https://doi.org/10.1590/S1519-69842009000300004
MacArthur RH, Pianka ER. On optimal use of a patchy environment. Am Nat. 1966; 100(916):603–09. https://doi.org/10.1086/282454
Manko P. Stomach content analysis in freshwater fish feeding ecology. Slovakia: University of Prešov Press; 2016.
Montag LF, Sousa LM, Netto-Ferreira AL, Sawakuchi AO, Hallwass G, Silvano RA et al. Amazonian rivers from the Brazilian Shield: Xingu and Tapajós. In: Rivers of South America. Amsterdam: Elsevier; 2025. p.381–405. https://doi.org/10.1016/B978-0-12-823429-7.00010-0
Moreira-Hara SS, Zuanon JA, Amadio SA. Feeding of Pellona flavipinnis Clupeiformes Pristigasteridae in a Central Amazonian floodplain. Iheringia Sér Zool. 2009; 99(2):153–57. https://doi.org/10.1590/S0073-47212009000200006
Mugnai R. Manual de identificação de macroinvertebrados aquáticos do Estado do Rio de Janeiro: para atividades técnicas, de ensino e treinamento em programas de avaliação da qualidade ecológica dos ecossistemas lóticos. Rio de Janeiro: Technical Books Editora; 2010.
Ng EL, Deroba JJ, Essington TE, Grüss A, Smith BE, Thorson JT. Predator stomach contents can provide accurate indices of prey biomass. ICES J Mar Sci. 2021; 78(3):1146–59. https://doi.org/10.1093/icesjms/fsab026
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara RB et al. Community ecology package. R package version. 2024; 2:321–26. Available from: https://cran.r-project.org/web/packages/vegan/
Parnell AC. SIMMR: Stable Isotope Mixing Model in R. R package version 0.4. 5. 2021. Available from: https://github.com/andrewcparnell/simmr
Parnell AC, Inger R, Bearhop S, Jackson AL. Source partitioning using stable isotopes: coping with too much variation. PLoS ONE. 2010; 5(3):e9672. https://doi.org/10.1371/journal.pone.0009672
Pebesma E, Bivand R, Rowlingson B, Gomez-Rubio V, Hijmans R, Sumner M et al. sp: Classes and methods for spatial data. R package version. 2025. https://doi.org/10.32614/CRAN.package.sp
Peel MC, Finlayson BL, McMahon TA. Updated world map of the Köppen-Geiger climate classification. Hydrol Earth Syst Sci. 2007; 11:1633–44. https://doi.org/10.5194/hess-11-1633-2007
Penha ICS, Jardim Jr. AA, Prata EG, Seabra LB, Gusmão RR, Ferreira GT et al. Feeding ecology of a detritivorous fish in controlled flood pulse area in the Amazon revealed by stomach content and stable isotopes analyses. Aquat Sci. 2025; 87(1):1–13. https://doi.org/10.1007/s00027-024-01147-x
Penha ICS, Seabra LB, Prata EG, Freitas TMS, Montag LFA. Unravelling a specialised diet of an Amazonian catfish in a controlled flood-pulse area by combining stomach-content and stable-isotope analyses. Mar Freshw Res. 2024; 75(6):MF23039. https://doi.org/10.1071/MF23039
Pereira LS, Tencatt LFC, Dias RM, Oliveira AG, Agostinho AA. Effects of long and short flooding years on the feeding ecology of piscivorous fish in floodplain river systems. Hydrobiologia. 2017; 795:65–80. https://doi.org/10.1007/s10750-017-3115-5
Peterson BJ, Fry B. Stable isotopes in ecosystem studies. Annu Rev Ecol Syst. 1987; 18:293–320. https://doi.org/10.1146/annurev.es.18.110187.001453
Phillips DL, Inger R, Bearhop S, Jackson AL, Moore JW, Parnell AC et al. Best practices for use of stable isotope mixing models in food-web studies. Can J Zool. 2014; 92(10):823–35. https://doi.org/10.1139/cjz-2014-0127
Pinnegar JK, Polunin NVC. Differential fractionation of δ13C and δ15N among fish tissues: implications for the study of trophic interactions. Funct Ecol. 1999; 13(2):225–31. https://doi.org/10.1046/j.1365-2435.1999.00301.x
Poiesz SSH, Witte JI, van der Veer HW. Stomach content analysis indicates multi decadal trophic stability in a temperate coastal fish food web, western Dutch Wadden Sea. Estuar Coast Shelf Sci. 2024; 308:108912. https://doi.org/10.1016/j.ecss.2024.108912
Post DM. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology. 2002; 83(3):703–18. https://doi.org/10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2
Post DM, Layman CA, Arrington DA, Takimoto G, Quattrochi J, Montana CG. Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia. 2007; 152:179–89. https://doi.org/10.1007/s00442-006-0630-x
Prata EG, Seabra LB, Neres-Lima V, Montag LFDA, Freitas TMS. Diet composition and isotopic analysis unveil trophic dynamics of a fish in a controlled flood pulse area of the Amazonia. Hydrobiologia. 2025; 852(3):689–703. https://doi.org/10.1007/s10750-024-05716-x
Qin J, Liu F, Schmidt BV, Sun Z, Kong L, Yan Y. Interpopulation trophic niches and ontogenetic shifts of a mangrove fish predator. J Fish Biol. 2024; 104(2):365–73. https://doi.org/10.1111/jfb.15219
Quaresma A, Zuquim G, Demarchi LO, Ribas CC, Wittmann F, Assunção AM et al. Belo Monte Dam impacts: protagonism of local people in research and monitoring reveals ecosystem service decay in Amazonian flooded vegetation. Perspect Ecol Conserv. 2025; 23(1):39–50. https://doi.org/10.1016/j.pecon.2025.02.001
Peixer J, Mateus LAF, Resende EK. First gonadal maturation of Pinirampus pirinampu (Siluriformes: Pimelodidae) in the Pantanal, Mato Grosso do Sul State, Brazil. Braz J Biol. 2006; 66:317–23. https://doi.org/10.1590/S1519-69842006000200014
R Development Core Team. R: a language and environment for statistical computing [Internet]. Vienna: R Foundation for Statistical Computing; 2024. Available from: https://www.r-project.org/
Richardson SM, Hanson JM, Locke A. Effects of impoundment and water-level fluctuations on macrophyte and macroinvertebrate communities of a dammed tidal river. Aquat Ecol. 2002; 36(4):493–510. https://doi.org/10.1023/A:1021137630654
Röpke C, Lonardoni A, Zuanon J, Doria CRC, Lima AL, Sant’Anna IRA et al. Diet seasonality and resource partitioning by large catfishes in the Madeira River, Brazil. Acta Amaz. 2025; 55:e55bc25003. https://doi.org/10.1590/1809-4392202500031
Seabra LB, Huckembeck S, Freitas TMS, Lobato CMC, Penha ICS, Prata EG et al. Variation in basal sources contribution to the diet of a predator fish in an altered flood pulse area in the Amazon. Hydrobiologia. 2025a; 852(4):909–25. https://doi.org/10.1007/s10750-024-05736-7
Seabra LB, Tedesco PA, Oberdorff T, Winemiller KO, Huckembeck S, Freitas TMS et al. Exploring isotopic patterns of fish trophic guilds in the Volta Grande reach of the Xingu River, eastern Amazon, regulated by the operation of a hydroelectric dams. Aquat Sci. 2025b; 87(3):1–13. https://doi.org/10.1007/s00027-025-01187-x
Silveira EL, Semmar N, Cartes JE, Tuset VM, Lombarte A, Ballester ELC et al. Methods for trophic ecology assessment in fishes: a critical review of stomach analyses. Rev Fish Sci Aquac. 2020; 28(1):71–106. https://doi.org/10.1080/23308249.2019.1678013
Skjoldal HR, Eriksen E, Ono K, Dolgov A. Diet and trophic structure of fishes in the Barents Sea: between empty and full stomachs-large individual variability follows a common pattern. J Fish Biol. 2025; 106(6):1734–51. https://doi.org/10.1111/jfb.16058
Smith JA, Mazumder D, Suthers IM, Taylor MD. To fit or not to fit: evaluating stable isotope mixing models using simulated mixing polygons. Methods Ecol Evol. 2013; 4(7):612–18. https://doi.org/10.1111/2041-210X.12048
Stephens DW, Krebs JR. Foraging theory. Princeton: Princeton University Press; 1986.
Syväranta J, Lensu A, Marjomäki TJ, Oksanen S, Jones RI. An empirical evaluation of the utility of convex hull and standard ellipse areas for assessing population niche widths from stable isotope data. PloS ONE. 2013; 8(2):e56094. https://doi.org/10.1371/journal.pone.0056094
Tockner K, Malard F, Ward JV. An extension of the flood pulse concept. Hydrol Process. 2000; 14(16–17): 2861–83. https://doi.org/10.1002/1099-1085(200011/12)14:16/17%2861::AID-HYP124%3.0.CO;2-F
Utsunomiya R, Beveridge C, Lobo G, Assahira C, Moretto EM, Athayde S. Dewatering the Xingu River: hydrological alterations and biocultural connections among the Arara Indigenous People in the Volta Grande region, Brazilian Amazon. Reg Environ Change. 2024; 24(2):85. https://doi.org/10.1007/s10113-024-02230-7
Valdelamar-Villegas J, Olivero-Verbel J. High mercury levels in the indigenous population of the Yaigojé Apaporis National Natural Park, Colombian Amazon. Biol Trace Elem Res. 2020; 194(1):3–12. https://doi.org/10.1007/s12011-019-01760-0
Vignon M, Dierking J. Prey regurgitation and stomach vacuity among groupers and snappers. Environ Biol Fish. 2011; 90(4):361–66. https://doi.org/10.1007/s10641-010-9746-2
Vinson MR, Angradi TR. Stomach emptiness in fishes: sources of variation and study design implications. Rev Fish Sci. 2011; 19(2):63–73. https://doi.org/10.1080/10641262.2010.536856
Weiss B, Costa F, Santana F, Klein VP, Lopes A, Costa G et al. Fish assemblage characteristics and allochthonous food resources shape the topology of trophic networks in blackwater floodplain forests. Wetlands. 2025; 45(5):1–14. https://doi.org/10.1007/s13157-025-01937-5
Winemiller KO. Factors driving temporal and spatial variation in aquatic floodplain food webs. In: Polis GA, Winemiller KO, editors. Food webs: integration of patterns and dynamics. Boston: Springer; 1996. p.298–312. https://doi.org/10.1007/978-1-4615-7007-3_29
Winemiller KO, Andrade MC, Arantes CC, Bokhutlo T, Bower LM, Cunha ER et al. Can spatial food web subsidies associated with river hydrology and lateral connectivity be detected using stable isotopes? Food Webs. 2023; 34:e00264. https://doi.org/10.1016/j.fooweb.2022.e00264
Winemiller KO, McIntyre PB, Castello L, Fluet-Chouinard E, Giarrizzo T, Nam S et al. Balancing hydropower and biodiversity in the Amazon, Congo, and Mekong. Science. 2016; 351(6269):128–29. https://doi.org/10.1126/science.aac7082
Zanden MJV, Rasmussen JB. Variation in δ15N and δ13C trophic fractionation: implications for aquatic food web studies. Limnol Oceanogr. 2001; 46(8):2061–66. https://doi.org/10.4319/lo.2001.46.8.206
ZAR JH. Biostatistical analysis. New Jersey. Prentice-Hall; 2010.
Zavala-Camin LA. Introdução aos estudos sobre alimentação natural em peixes. Maringá: Editora da Universidade Estadual de Maringá; 1996.
Zuanon J, Sawakuchi AO, Camargo M, Wahnfried I, Sousa L, Akama A et al. Condições para a manutenção da dinâmica sazonal de inundação, a conservação do ecossistema aquático e manutenção dos modos de vida dos povos da Volta Grande do Xingu. Papers NAEA. 2019; 1(2):20–62. https://doi.org/10.18542/papersnaea.v28i2.8106
Authors

![]() Antonio Augusto Jardim Jr1,2
Antonio Augusto Jardim Jr1,2 ![]() ,
, ![]() Erival Gonçalves Prata1,
Erival Gonçalves Prata1, ![]() Lais Martins Silva1,
Lais Martins Silva1, ![]() João Pedro da Silva Moraes1,
João Pedro da Silva Moraes1, ![]() Lucas Pires de Oliveira1,2,
Lucas Pires de Oliveira1,2, ![]() Luciano Fogaça de Assis Montag1,2 and
Luciano Fogaça de Assis Montag1,2 and ![]() Tiago Magalhães da Silva Freitas3
Tiago Magalhães da Silva Freitas3
[1] Laboratório de Ecologia e Conservação, Instituto de Ciências Biológicas, Universidade Federal do Pará, Rua Augusto Corrêa, 01, 66075-110, Belém, PA, Brazil. (AAJJ) biosantosjr@gmail.com (corresponding author), (EGP) erival.gprata@gmail.com, (LMS) lais.martins.silva@icb.ufpa.br, (JPSM) jopedromoraes@gmail.com, (LPO) lucspires10@gmail.com, (LFAM) lfamontag@gmail.com.
[2] Programa de Pós-Graduação em Ecologia, Instituto de Ciências Biológicas, Universidade Federal do Pará, Rua Augusto Corrêa, 01, 66075-110, Belém, PA, Brazil.
[3] Laboratório de Zoologia, Faculdade de Ciências Naturais, Campus Universitário do Marajó-Breves, Universidade Federal do Pará, Alameda IV, 3418, 68000-000, Breves, PA, Brazil. (TMSF) freitastms@gmail.com.
Authors’ Contribution 

Antonio Augusto Jardim Jr: Conceptualization, Formal analysis, Writing-original draft, Writing-review and editing.
Erival Gonçalves Prata: Conceptualization, Formal analysis, Writing-original draft, Writing-review and editing.
Lais Martins Silva: Formal analysis, Writing-original draft.
João Pedro da Silva Moraes: Formal analysis, Writing-original draft.
Lucas Pires de Oliveira: Formal analysis, Writing-original draft.
Luciano Fogaça de Assis Montag: Conceptualization, Funding acquisition, Methodology, Project administration, Writing-original draft, Writing-review and editing.
Tiago Magalhães da Silva Freitas: Conceptualization, Data curation, Methodology, Writing-original draft, Writing-review and editing.
Ethical Statement
Experiments were approved by the Ethical Committee for Animal Use in Experiments of the Universidade Federal do Pará (CEUA number 8293020418/2020) and Collection Licenses of SISBIO number 1267/2020.
Competing Interests
The author declares no competing interests.
Data availability statement
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
AI statement
The authors did not use any AI-assisted technologies in the creation of this manuscript or its figures.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) – Financing Code 001. We are grateful for funding from the authors’ grants: AAJJ (Process No 88887.687748/2022–00) and LPO (Process No 88887.939430/2024–00). LFAM thanks the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the research productivity grant (Process No 302881/2022–1).
Supplementary Material
Supplementary material SUP
Peer Review
How to cite this article
Jardim Jr AA, Prata EG, Silva LM, Moraes JPS, Oliveira LP, Montag LFA, Freitas TMS. Trophic dynamics of a carnivorous catfish under dam-regulated flood conditions in the Amazon. Neotrop Ichthyol. 2026; 24(2):e250114. https://doi.org/10.1590/1982-0224-2025-0114
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2025 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted January 12, 2026
Accepted January 12, 2026
![]() Submitted June 27, 2025
Submitted June 27, 2025
![]() Epub June 05, 2026
Epub June 05, 2026