![]() Dieison André Moi1
Dieison André Moi1 ![]() ,
, ![]() Alejandro D’Anatro2,
Alejandro D’Anatro2, ![]() Ivan González-Bergonzoni3 and
Ivan González-Bergonzoni3 and ![]() Franco Teixeira-de-Mello4
Franco Teixeira-de-Mello4
PDF: Download Here | Supplementary: Sup | Cite this article
Associate Editor: ![]() Evelyn Habit
Evelyn Habit
Section Editor: ![]() Fernando Pelicice
Fernando Pelicice
Editor-in-chief: ![]() José Birindelli
José Birindelli
Abstract
La diversidad de peces neotropicales ha disminuido en las últimas décadas, impulsada por el aumento de las presiones humanas. Esta pérdida acelerada destaca la urgente necesidad de métodos estandarizados para monitorear estas comunidades de manera confiable. En este estudio, evaluamos la eficacia de las redes agalleras tipo nórdicas, de malla múltiple, para el muestreo de comunidades de peces en el Río Uruguay. Las especies fueron capturadas en todos los tipos de mallas, las más pequeñas hasta las más grandes (5–55 mm, de nudo a nudo). El desempeño del modelo varió entre especies, con clara evidencia de selectividad por talla. Se observó un sesgo de tamaño, principalmente en las clases de longitud más pequeñas de especies de Siluriformes atrapadas en las mallas grandes. Las redes nórdicas de malla múltiple fueron eficientes para capturar especies de tamaño corporal pequeño, mientras que su eficiencia disminuyó para especies de grandes. Estos hallazgos indican que este tipo de redes son útiles para monitorear comunidades de peces en la región Neotropical, pero requieren correcciones por sesgo de tamaño, particularmente para los Siluriformes. Aunque este estudio respalda el uso de estas redes, demuestra que deberían complementarse con mallas de mayor tamaño si se requiere muestrear peces de mayores dimensiones en ríos neotropicales.
Palabras clave: Biodiversidad, Characiformes, Ecosistema fluvial, Neotrópico, Siluriformes.
Introduction
Neotropical freshwater fish are increasingly threatened by anthropogenic pressures, including pollution, biological invasions and overfishing (Shantz et al., 2020; Pelicice et al., 2021; Moi et al., 2025). Hence, standardized long-term monitoring of neotropical freshwater fish is essential (Magurran et al., 2010). In this context, multi-mesh gillnets are among the most widely used standardized sampling tools in certain regions, such as Europe (Millar, Holst, 1997; Carol, García-Berthou, 2007). This method allows simultaneous fishing with multiple mesh sizes, thereby capturing a broad range of body sizes. Although the use of multi-mesh gillnets has been standardized across Europe (Appelberg et al.,1995), no equivalent method exists in the Neotropical region.
Neotropical freshwaters harbor more than 4,200 fish species (Tonella et al., 2023). Neotropical fish assemblages are highly diverse and are dominated by Characiformes and Siluriformes, which exhibit a broad range of body sizes and morphological traits (Albert et al., 2020). Given this highly diverse taxonomic and functional structure, combined with the intensification of anthropogenic pressures, it is necessary to establish standardized gillnet-based methods capable of assessing catch-length distributions, population size structure, and species abundance in Neotropical fish assemblages, while also enabling evaluations of anthropogenic impacts (Moi et al., 2024, 2025). Although multi-mesh gillnets have been adopted in recent Neotropical studies (e.g., González-Bergonzoni et al., 2019, 2020; López-Rodriguez et al., 2019; Moi et al., 2024), their selectivity for these diverse fish communities remains untested.
Gillnets are highly selective and passive fishing gears, and their efficiency is directly related to the probability that fish will encounter them (Millar, Holst, 1997). Therefore, gillnets are ineffective for catching sedentary fish, and seasonal differences in individual movement can alter the encounter probability (Neumann, Willis, 1995). The probability of capture also depends on the fish being successfully entangled in the net. Considering that when a fish is entangled in a net, it involves several parts of its body, such as the operculum, spines, maxilla, and teeth, the probability of capture increases when the organism has more external structures (Shoup, Ryswyk, 2016). Hence, mesh size is a key factor influencing the size of individuals caught in gillnets (Hubert et al., 2012; Tesfaye et al., 2016). It is also strongly size-selective, because the length of an individual caught in a given mesh size usually differs by less than 20% of the length most efficiently retained by the mesh (Hamely, 1975, 1980).
To minimize size bias in fish sampling, gillnets with multiple mesh sizes are deployed simultaneously. Although this approach minimizes bias, it does not eliminate selectivity entirely. The length-frequency distributions obtained from gillnet catches may not accurately reflect the true proportions of each species in the environment (Shoup, Ryswyk, 2016). Previous studies have quantified the gillnet selectivity and proposed correction methods for length-frequency data (e.g., Shoup, Ryswyk, 2016; Smith et al., 2017; Klein et al., 2019). However, these corrections have been developed primarily for the Holarctic region, and no analogous corrections exist for Neotropical systems. Regional differences are important, as fish communities vary markedly across biogeographic realms. For example, in the Holarctic, orders such as Cypriniformes, Perciformes, and Salmoniformes predominate. These fish typically lack prominent external structures such as pectoral spines, and plates being mostly wrapped around their opercles and fins. In Neotropical fish communities, Siluriformes and Characiformes are highly diverse and abundant. In particular, the family Loricariidae (“armored catfish”) can reach high richness and abundance in these rivers (Albert et al., 2020). This family includes several fish with large pectoral and dorsal fin spines, often time serrated, bony plates along the body and even ossified barbels, all of them being structures easily wrapped in gillnets. Consequently, gillnet selectivity may differ substantially between regions dominated by morphologically distinct taxa.
Gillnet selectivity may be analyzed directly or indirectly (Millar, Fryer, 1999). The direct method is implemented if fish lengths distributions are known, but as this knowledge is rare, this method is generally not feasible in practice (Millar, Holst, 1997). As such, gillnet selectivities are evaluated by the indirect methods, in which different mesh sizes are employed at the same time, and size-specific selectivities are assessed through comparison of the capture of each individual length class between the meshes (Millar, Holst, 1997; Vandergoot et al., 2011). The indirect method is quantified with selectivity curves where the probability of capture for each fish length class by each mesh size is calculated as a proportion of the total catch of the length class across all mesh sizes (Shoup, Ryswyk, 2016).
The probability of capture indicates how efficiently that mesh size catches fish of the optimum length. Millar, Holst (1997) provide a statistical approach for indirectly estimating gillnet retention curves through generalized linear modelling (the SELECT method). This method offers a critical approach to size selectivity analysis and has been used in gillnet selectivity studies (e.g., Rodríguez-Climent et al., 2012; Radomski et al., 2020). Despite its importance, the indirect method presents a weakness because it evaluates gillnet selectivity only for fish that are collected by the net and not for the overall fish population (Shoup, Ryswyk, 2016). This results in gillnet size bias being related to the morphology of the retained fish, rather than to differences in size-specific encounter rates (Booth, Potts, 2006). To correct the size bias of the indirect method, Shoup, Ryswyk (2016) developed a correction factor derived from the selectivity curves, which allows for an accurate examination of the contact selectivity of the fish population sampled.
In this study, we monitored the fish community in a neotropical river (the Río Uruguay) over seven years. We used benthic nordic multi-mesh gillnet (Appelberg et al., 1995; Olin et al., 2004). This multi-mesh gillnet has proven useful for managers in European countries, as it provides comprehensive estimates of fish size distributions (Appelberg et al., 1995). The use of this multi-mesh net could also be useful for managers in the neotropical region. To our knowledge, no study has assessed the selectivity of the nordic multi-mesh gillnet for Neotropical fish, which could inform the implementation of this method in these hyperdiverse rivers. Hence, we aimed to identify the best-fit selectivity model for gillnet catches in the Río Uruguay.
Material and methods
Study area. The study was performed in the Río Uruguay, which has an approximate length of 1,838 km and a drainage area of 365,000 km². In this river, the flood pulse is driven mainly by precipitation in the upper portion of its catchment, which increases during El Niño events, producing irregular flood peaks approximately every 3.5–6 years (Krepper et al., 2003). We sampled fish communities at three sites distributed along a 40-km stretch of the lower Río Uruguay, from upstream to downstream, respectively: i) Nuevo Berlín, ii) Fray Bentos, and iii) Las Cañas (Fig. 1).
Fish sampling. The fish community was sampled twice a year in April (fall) and December (spring) at the three river sites mentioned above. We used standard nordic multi-mesh gillnets, consisting of 30 m-long nets with 12 mesh panels of varying sizes, with measures knot to knot of: 5, 6.25, 8, 10, 12.5, 15.5, 19.5, 24, 29, 33, 43, and 55 mm, respectively. To provide a comprehensive spatial monitoring assessment, fish samples were collected at the exact same locations in autumn and spring. In total, we used eight individual nordic multi-mesh gillnets, joining two nets together at each location, hence, resulting in four sets composed by two nordic multi-mesh gillnet each one. Thus, each set measured a total length of 60 m length and 1.5 m height, resulting in an area of 90 m². We positioned two of these 60 m-long sets parallel to each other in the littoral zone, at a maximum distance of 100 m from the coast, with 300 m between them and a depth of 1–2 m. The other two multi-mesh gillnet sets were located at 500 m from the coast in line with the littoral ones, with 300 m between them and at a depth of 2–5 m. Nordic multi-mesh gillnets were deployed from 7 p.m. to 7 a.m., covering approximately 12 h during the twilight and night period. For this study, fish caught in 46 nordic multi-mesh gillnets were analyzed. This number corresponds to selecting one Nordic multi-mesh gillnet per sampling event. During the 8-year monitoring period (2009–2016), 48 sampling events were planned (two per year at three sites), with one Nordic multi-mesh gillnet randomly selected in each event for the selectivity analysis. Two events were lost, resulting in 46 gillnets, which together represent a total sampled gillnet area of 2,070 m² across autumn and spring campaigns. Fish were identified based on the twelve different mesh sizes in which the fish were caught. The collected fish were identified at the species level, counted, and measured (standard length, cm). All fish vouchers are in the fish collections of the Facultad de Ciencias, Universidad de la República, Montevideo (ZVCP) and Centro Universitario Regional del Este (CURE-Ict).
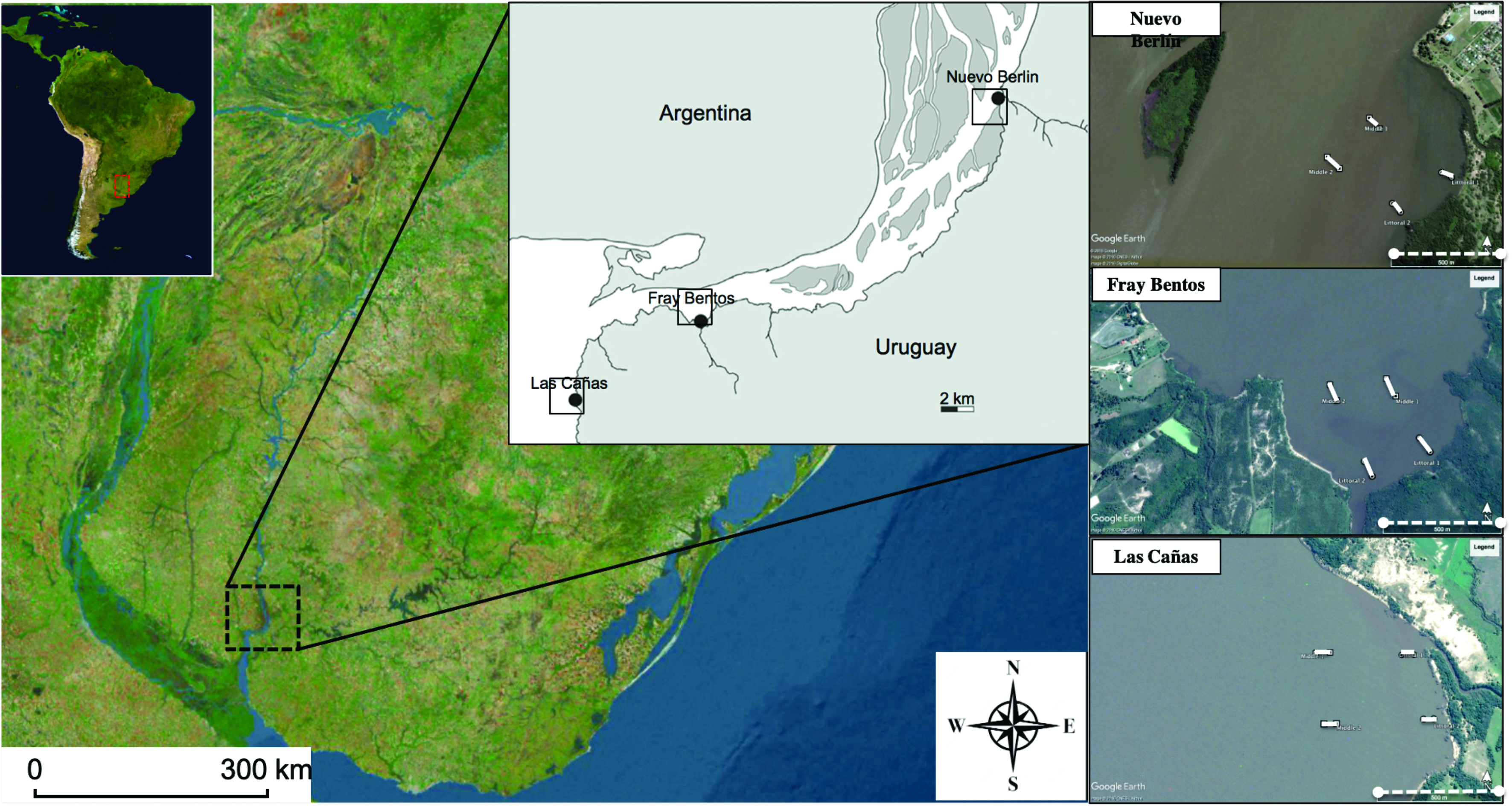
FIGURE 1| Map of the Uruguay River, highlighting the three distinct sites within the river that have been monitored from 2009 to 2016. The dots at each site indicate the locations where the fish collection nordic multi-mesh gillnets were deployed. The rectangles represent the exact sites where the full set of two nordic multi-mesh gillnets was deployed, totaling eight Nordic multi-mesh gillnets.
Data analysis. We selected species with more than 60 individuals caught during monitoring for the gillnet selectivity analysis (Tab. S1). A total of 14 species matched this criterion: Apareiodon affinis, Psalidodon spp., Piabarchus stramineus, Cynopotamus argenteus, Cyphocharax platanus, Iheringichthys labrosus, Megaleporinus obtusidens, Loricariichthys melanocheilus, Pachyurus bonariensis, Parapimelodus valenciennis, Pimelodella gracilis, Pimelodus maculatus, Prochilodus lineatus and Ricola macrops (see Tab. S1).
We estimated gillnet selectivity for 14 species using Millar’s (1992) SELECT approach, implemented in the R package TropFishR (Mildenberger et al.,2017). The SELECT uses a generalized linear model, assuming Poisson distribution of the gillnet catch data to fit them to a specified model using Maximum Likelihood (Millar, Holst, 1997). The selectivity’s curves are fitted as the relative probability of a fish of a given length to be captured when contacting a mesh of a determinate size (Millar, Holst, 1997). We tested four selectivity models, Normal location, Normal scale, Lognormal, and Gamma (Fig. 2), which correspond to different types of statistical distributions used to describe the probability of capture across fish lengths. The Normal scale, Gamma, and Lognormal models follow the principle of geometric similarity, meaning that both the mean and the standard deviation of the retention curve shift proportionally with mesh size (Millar, Holst, 1997). In contrast, the Normal location model assumes a proportional increase of the mean with mesh size but a fixed standard deviation, so all gillnet variants have the same variability (Millar, Holst, 1997). The Normal location and Normal scale models assume a symmetric (Gaussian) distribution, whereas the Gamma and Log-normal models assume a skewed distribution (Millar, Holst, 1997). The choice of these distribution affects the shape of the selectivity curve and how well it fits the observed catch data. Catch rates were summarized into 42 length classes ranging from 30 to 439 mm, with all classes spanning 10 mm to cover the full-size range of the sampled fish. The best-fit model for each species was determined based on the lowest model deviance and most randomly distributed residuals (Millar, Holst, 1997). We also used overdispersion to determine the best model as suggested by Holst et al. (1998). The dispersion parameter is estimated by the ratio of deviations vs. degrees of freedom. Specifically, a dispersion parameter > 1 indicates excessive dispersion.
Previous assessments have found bias in gillnet selectivity’s to certain fish length classes, leading to underestimating selectivity to some fish length classes (Hansen et al., 1997; Shoup, Ryswyk, 2016). To minimize bias, we used the method of Shoup and Ryswyk (2016) to estimate the magnitude of the size bias and to correct the length frequency of individuals that contacted the net. The method uses selectivity curves to adjust the length-frequency distributions of each fish species (Hansen et al., 1997). Selectivity-adjusted data are compared with unadjusted (Shoup, Ryswyk, 2016). The relative selectivity values (S) for employed nordic multi-mesh gillnets, i.e., normalizing selectivity across the mesh sizes, were calculated as:
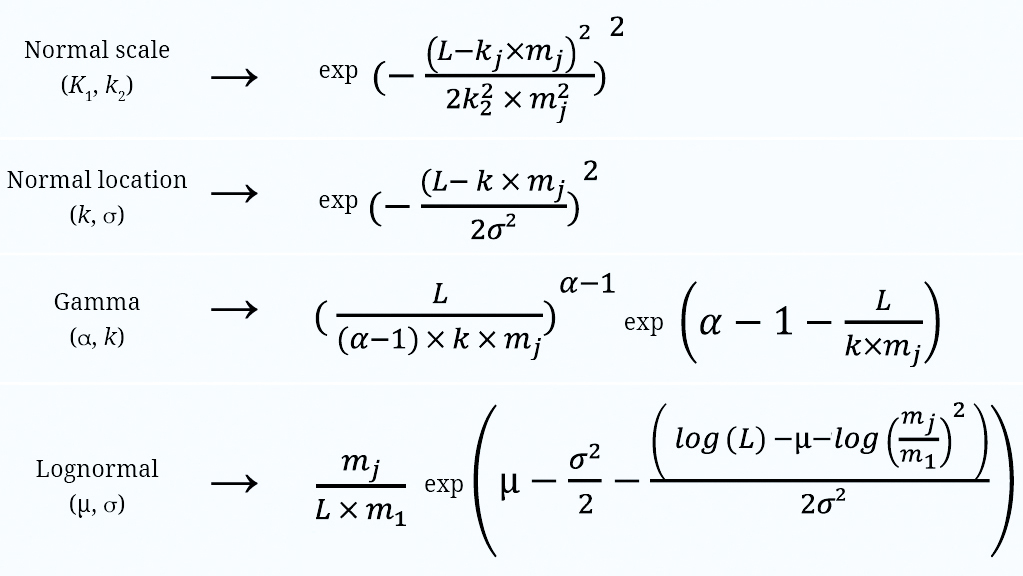
FIGURE 2| Equations and parameters for the four SELECT models. Equations are related to the mesh size j (mj) with number of fish of length L captured in that mesh size. Other symbols used in equations are constant in models. Fitted constants describing contact selectivity for the fish community in Uruguay. We fitted models for all 14 species (occurrence > 60 individuals). Models were fitted with Normal scale, Normal location, Gamma e Lognormal distributions.
Sl = Σ(Sj(l)/maxl)
Where sj(l) is the selectivity of size class l in mesh size j, and maxl is the maximum selectivity value [sj(l)] observed among all length classes (Shoup, Ryswyk, 2016). We adjusted the abundance of each length class (i.e., adjusted fish length-frequency) by dividing the total number of fish captured in a given length class (catch from all mesh sizes) by the overall net selectivity value (Sl) for that length class (Shoup, Ryswyk, 2016). We then compared the unadjusted and Sl–adjusted length-frequency data for each species using a variance analysis (ANOVA). Our data met the assumptions of normality and equal variance, as indicated by the graphical analysis of residuals.
Results
We caught 75 fish species represented by 4,458 individuals, where 1,170 individuals belonged to the Characiformes and 2,008 individuals to the Siluriformes (Tab. S1). These were the two orders most commonly caught, accounting for 85% of fish captured. The overall catches decreased with the increase in mesh sizes, where smaller mesh caught more fish than larger mesh (Fig. 3). All mesh sizes caught a wide size range of several fish species. Nevertheless, the mean standard deviation in length of sampled fish for all 14 species increased with mesh size (Tab. S2).
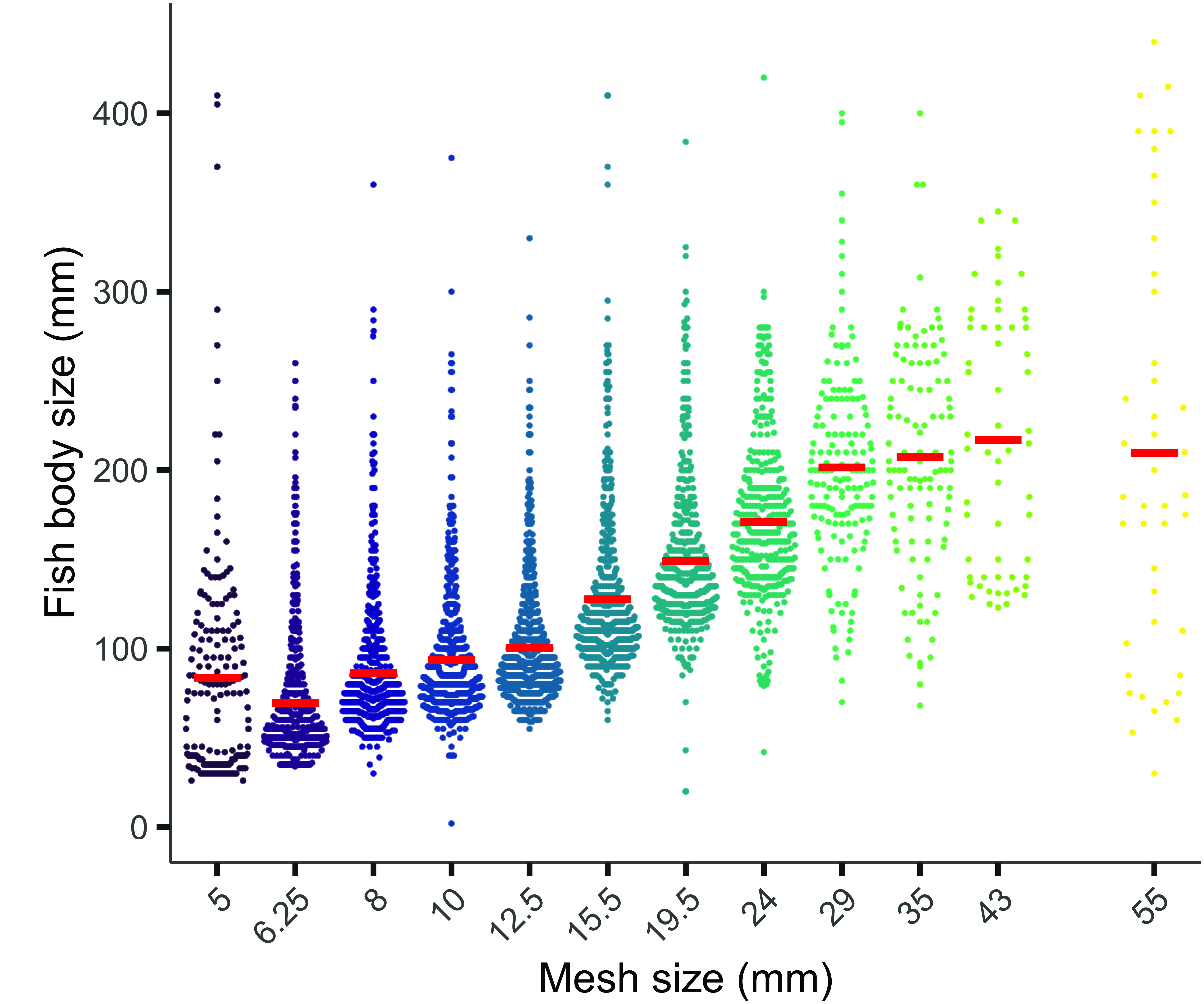
FIGURE 3| Length class of fish (mm) caught in each mesh size (5, 6.25, 8, 10, 12.5, 15.5, 19.5, 24, 29, 35, 43, and 55 mm) used by “nordic multimesh gillnet”. The horizontal red line represents the average body size of fish caught in each mesh size.
The best-fitting model estimated using the SELECT method, varied considerably among the 14 species (Tab. 1). Assuming equal fishing power for all meshes, the normal scale model had the lowest deviance value for the A. affinis, Psalidodon spp., P. stramineus,and P. gracilis (Fig. 4). The normal location model had the lowest deviance value for C. platanus, P. lineatus, Pimelodus sp., and R. macrops (Fig. 4). The lognormal model had the lowest deviance value for C. argenteus, M. obtusidens, I. labrosus, L. melanocheilus, P. valenciennis and P. bonariensis (Fig. 4). We found that small mesh sizes had the highest selectivity for smaller length classes, whereas the large mesh size had the highest selectivity for the largest length classes. In addition, the smallest mesh sizes (i.e., 5, 6.25, 8 and 10 mm) showed very similar selectivity curves for the length of the fish caught (Fig. 4), especially for the Siluriformes species, which suggests that these mesh sizes overlap.
TABLE 1 | Model parameters, residual deviance, degrees of freedom (df), for four gillnet selectivity models (i.e., Normal scale, Normal location, Log-normal, and Gamma) estimate using SELECT method. Parameters 1 and 2 are k and σ for normal location model; k1 and k2for normal scale model (spread proportional to mesh size); α and k for gamma model; µ1 and σ for lognormal model. The model with lowest deviance for each of 14 species is indicated in bold text. Abundance is the number of individuals of each species that were included in the analyses.
| Fish model | Abundance | Model | Fishing power relative mesh size | |||
| Par. 1 | Par. 2 | Deviance | df | |||
| Apareiodon affinis | 264 | Normal location | 4.79 | 7461.93 | 850.21 | 419 |
| Normal scale | 41.42 | 13.39 | 389.76 | 419 | ||
| Gamma | 5.31 | 1.72 | 745.512 | 419 | ||
| Lognormal | 3.91 | 0.35 | 806.51 | 419 | ||
| Psalidodon spp. | 526 | Normal location | 5.93 | 158.66 | 502.08 | 262 |
| Normal scale | 6.25 | 0.83 | 258.16 | 262 | ||
| Gamma | 45.58 | 0.13 | 381.41 | 262 | ||
| Lognormal | 3.43 | 0.02 | 363.01 | 262 | ||
| Piabarchus stramineus | 296 | Normal location | 0.33 | 53.71 | 485.02 | 60 |
| Normal scale | 7.53 | 0.82 | 55.63 | 60 | ||
| Gamma | 48.60 | 0.15 | 116.12 | 60 | ||
| Lognormal | 3.64 | 0.02 | 155.21 | 60 | ||
| Cynopotamus argenteus | 79 | Normal location | 6.87 | 2507.78 | 160.81 | 264 |
| Normal scale | 9.41 | 33.67 | 203.71 | 264 | ||
| Gamma | 6.90 | 1.44 | 171.69 | 264 | ||
| Lognormal | 3.81 | 0.13 | 157.21 | 264 | ||
| Cyphocharax platanus | 254 | Normal location | 6.16 | 485.59 | 187.52 | 198 |
| Normal scale | 7.53 | 6.80 | 615.45 | 198 | ||
| Gamma | 16.12 | 0.44 | 446.63 | 198 | ||
| Lognormal | 3.53 | 0.04 | 376.15 | 198 | ||
| Iheringichthys labrosus | 955 | Normal location | 8.27 | 49.28 | 483.34 | 364 |
| Normal scale | 12.57 | 25.36 | 634.12 | 364 | ||
| Gamma | 5.94 | 2.10 | 378.01 | 364 | ||
| Lognormal | 4.07 | 0.21 | 319.52 | 364 | ||
| Megaleporinus obtusidens | 157 | Normal location | 6.75 | 30.84 | 341.58 | 360 |
| Normal scale | 8.95 | 10.03 | 352.34 | 360 | ||
| Gamma | 9.38 | 0.96 | 328.23 | 360 | ||
| Lognormal | 3.75 | 0.11 | 319.68 | 360 | ||
| Loricariichthys melanocheilus | 168 | Normal location | 11.35 | 60.17 | 220.03 | 324 |
| Normal scale | 17.04 | 55.94 | 297.54 | 324 | ||
| Gamma | 6.72 | 2.48 | 235.83 | 324 | ||
| Lognormal | 4.34 | 0.15 | 216.23 | 324 | ||
| Pachyurus bonariensis | 260 | Normal location | 6.31 | 870.35 | 366.24 | 220 |
| Normal scale | 7.44 | 3.57 | 254.73 | 220 | ||
| Gamma | 17.10 | 0.42 | 270.57 | 220 | ||
| Lognormal | 3.56 | 0.06 | 214.41 | 220 | ||
| Parapimelodus valenciennis | 300 | Normal location | 7.50 | 193.61 | 355.23 | 276 |
| Normal scale | 9.43 | 10.56 | 440.23 | 276 | ||
| Gamma | 11.62 | 0.77 | 285.10 | 276 | ||
| Lognormal | 3.75 | 0.08 | 251.36 | 276 | ||
| Pimelodella gracilis | 146 | Normal location | 9.51 | 366.65 | 182.54 | 276 |
| Normal scale | 8.89 | 3.03 | 122.44 | 276 | ||
| Gamma | 18.10 | 0.50 | 150.52 | 276 | ||
| Lognormal | 3.81 | 0.07 | 176.09 | 276 | ||
| Pimelodus sp. | 105 | Normal location | 7.34 | 144.46 | 112.19 | 324 |
| Normal scale | 10.59 | 16.23 | 180.29 | 324 | ||
| Gamma | 8.02 | 1.29 | 147.43 | 324 | ||
| Lognormal | 3.87 | 0.12 | 134.06 | 324 | ||
| Prochilodus lineatus | 63 | Normal location | 4.95 | 1159.48 | 130.01 | 324 |
| Normal scale | 5.77 | 2.90 | 164.80 | 324 | ||
| Gamma | 17.66 | 0.33 | 145.41 | 324 | ||
| Lognormal | 3.34 | 0.05 | 137.23 | 324 | ||
| Ricola macrops | 107 | Normal location | 10.29 | 82.65 | 152.83 | 288 |
| Normal scale | 20.33 | 138.16 | 199.91 | 288 | ||
| Gamma | 3.95 | 5.29 | 175.02 | 288 | ||
| Lognormal | 4.54 | 0.31 | 163.81 | 288 | ||
We also found some degree of size bias for all 14 fish species monitored. Through overall selectivity curves, we identified length-specific relative selectivity values (Rel Sl) for each length class, which typically corrected these biases For most species (A. affinis, Psalidodon spp., C. argenteus, C. platanus, M. obtusidens, I. labrosus, P. valenciennis, P. gracilis, L. melanocheilus, R. macrops, and P. bonariensis), the highest Rel Sl was observed at intermediate length classes (100–300 mm), whereas the smallest classes had the lowest Rel Sl values (Tabs. S3, S4). In contrast, for P. stramineus, P. lineatus and Pimelodus sp.the peak selectivity occurred in smaller length classes (< 150 mm) (Tab. S2). No fish species had peak selectivity in larger length class (> 400 mm).
Comparing the corrected length frequencies (i.e., adjusted by Rel Sl; Tabs. S3, S4) with unadjusted data, we found similar values for A. affinis, Psalidodon spp., P. stramineus, C. argenteus, C. platanus, M. obtusidens, and P. bonariensis, implying no significant biases for these species (Fig. 5). In contrast, we found significant differences between adjusted and unadjusted data for siluriform species, including I. labrosus, L. melanocheilus, P. valenciennis, P. gracilis, Pimelodus sp.,and R. macrops (Fig. 5). For Siluriformes species biases occurred primarily in the smallest length classes, particularly those below 100 mm, where the adjusted data were substantially higher than the unadjusted values (Fig. 5).
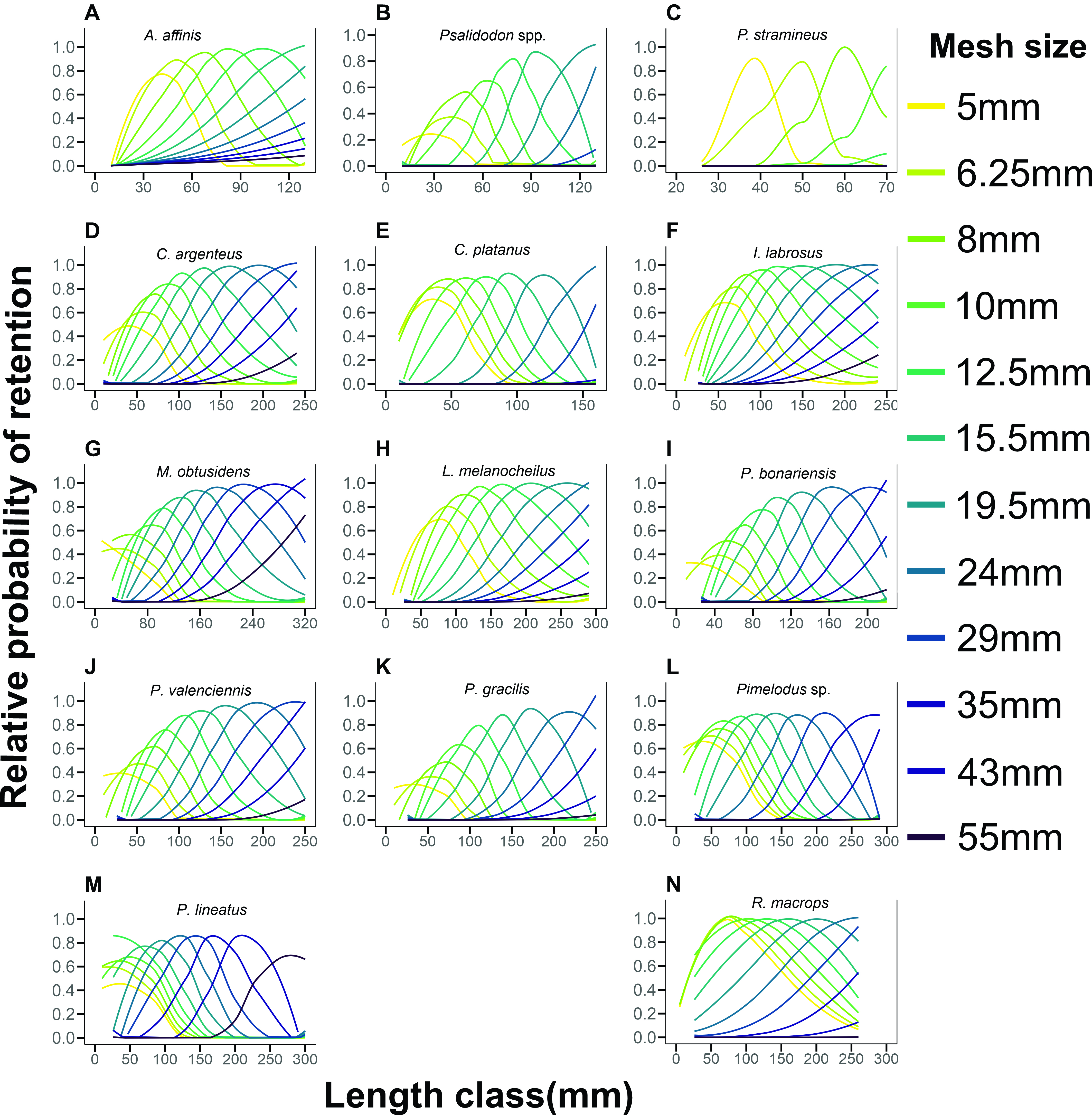
FIGURE 4| Selectivity curves of “Nordic Gillnet multimesh” fitted for 14 species, sampled in Uruguay River, during six years. The retention curves from left to right correspond to 5, 6.25, 8, 10, 12.5, 15.5, 19.5, 24, 29, 35, 43, and 55-mm mesh, respectively.
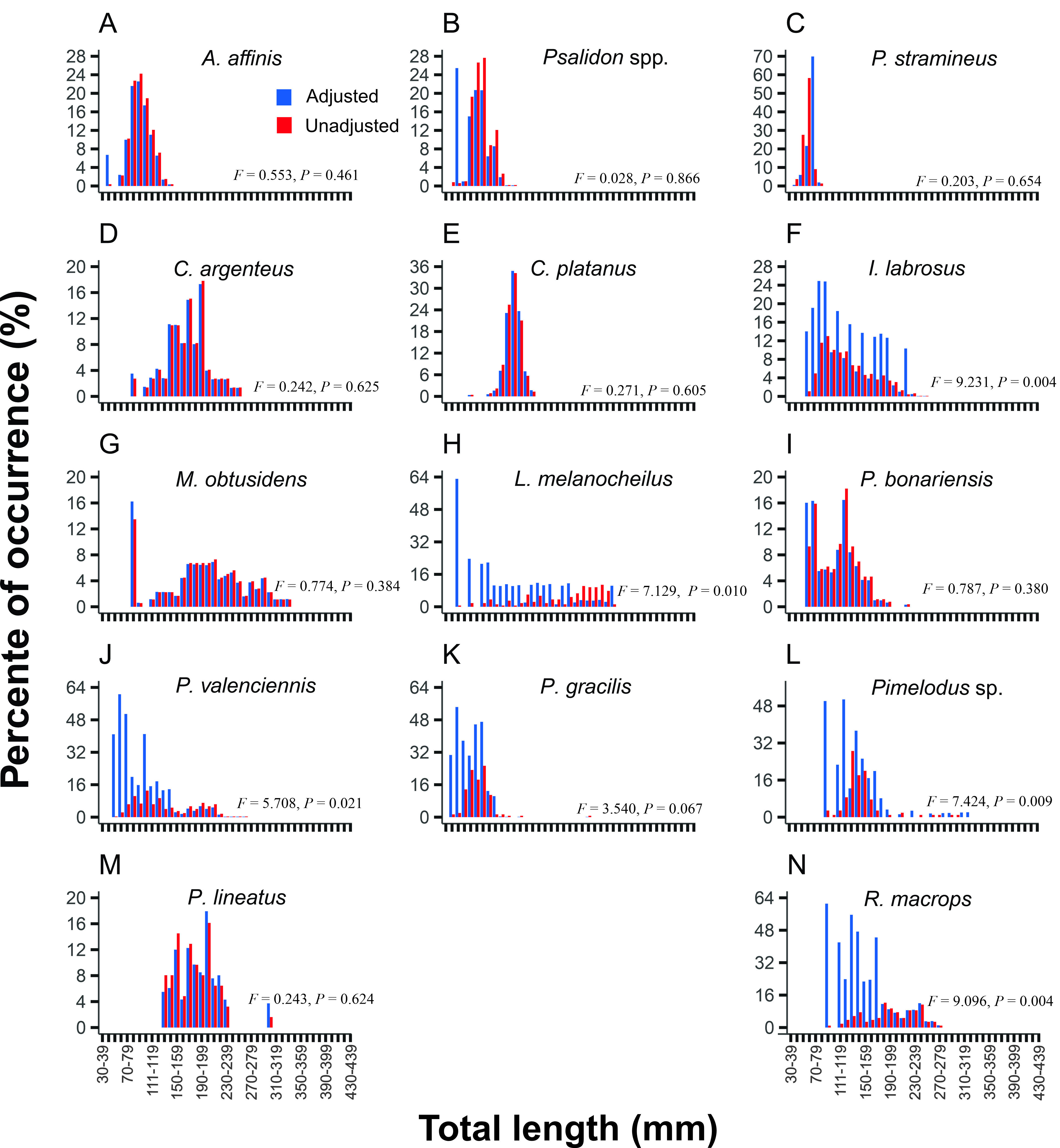
FIGURE 5| Percent occurrence of 14 fish species per length class across three sites in the Río Uruguay, sampled during seven years using nordic multimesh gillnet. Catch rates were summarized into 42 length classes ranging from 30 to 439 mm, with all classes spanning 10 mm to cover the entire size range of the sampled fish. Blue bars indicate adjusted data for contact selectivity and red bars indicate data unadjusted. The differences between the adjusted and unadjusted data were calculated using analysis of variance (ANOVA). F and P values are shown within each graph, where P < 0.05 indicates significant differences.
Discussion
Based on monitoring data, this study evaluated the selectivity of the “nordic multi-mesh gillnet” for 14 fish species along the Río Uruguay. We found that Siluriformes and Characiformes were the most commonly captured orders, matching their high abundance in Neotropical fish assemblages (Albert et al., 2020; Tonella et al., 2023). In general, fish length increased with mesh size (Fig. 3), highlighting the size selectivity of the nordic multi-mesh gillnet. Different selectivity models were more suitable for different species, indicating that model performance is species-specific (Millar, Fryer, 1999; Carol, García-Berthou, 2007; Tesfay et al., 2016). Several Siluriformes species showed evidence of size bias, which was corrected using overall selectivity curves (adjusted by Rel Sl) following Shoup and Ryswyk (2016). These findings indicate that the nordic multi-mesh gillnet may be effectively used for monitoring fish stocks in Neotropical rivers, although size biases may occur for Siluriformes.
Most of the fish species monitored in the Río Uruguay are small-bodied, usually reaching less than 200 mm at the adult stage (e.g., A. affinis, Psalidodon spp., P. stramineus, C. platanus). For these species, we showed that the nordic multi-mesh gillnet was able to catch individuals of a full range of body sizes. However, some species (e.g., M. obtusidens and P. lineatus) are large-bodied, reaching over 700 mm in adulthood. For these species, the nordic multi-mesh gillnets only caught individuals smaller than 300 mm, meaning that they failed to represent the full-size range of large-bodied fishes. This limitation arises not from the gillnet design itself, but from the fact that its mesh sizes were originally developed for European streams, with a maximum of 55 mm (Appelberg et al., 1995). As such, using nordic multi-mesh gillnets for monitoring small-bodied fish is highly effective, whereas if the objective is to monitor large species, this gillnet should include segments with larger meshes. Importantly, the nordic design is based on a geometric progression of mesh sizes, with a fixed ratio of 1.25 between consecutive meshes (Appelberg et al., 1995). Therefore, nordic multi-mesh gillnets could be adapted to catch large fish by increasing the mesh size following a geometric scale with a ratio of 1.25 (Olin et al., 2004; Specziár et al., 2009). By continuing this progression, the new mesh sizes would be: 70 mm, 90 mm, 115 mm, 145 mm, 185 mm, and 240 mm, which are consistent with those used to capture large-bodied fishes in other Neotropical rivers (Moi et al., 2021; Santos et al., 2022). We therefore suggest that further studies adapt the nordic multi-mesh gillnet to also target large-bodied individuals, which have high economic importance (Pelicice et al., 2023).
Our results have important applicability, since researchers and managers may use the selectivity curves to adjust catch data when using the nordic multi-mesh gillnet. Data were adjusted (i.e., corrected length-frequency) by dividing the number of fish captured within a given length class by the overall probability of retention (Sl value) (Tab. S3) for that length class (Hansen et al., 1997). In this method, for instance, if a given net captures ten individuals of Apareidon affinis within the 70–79 mm length class, where Sl = 0.5 for that length class during one twilight, then the catch of A. affinis should be adjusted to 20 individuals per twilight. In the aforementioned example, we divide the number of individuals captured by the Sl value (10/0.5) to adjust for the size bias against this length class. Perhaps more importantly, when we adjusted the number of fish captured, we found twice as many fish as in the unadjusted data. This means that we should double the number of fish captured to account for fish that have contacted the net without being retained.
We showed that Siluriformes species were abundant in length classes with low selectivity. This implies that these species had an Sl value closer to 0 than to 1, which resulted in large adjusted values. By contrast, Characiformes were abundant in length classes with high selectivity; that is, these species showed an Sl closer to 1 than to 0, resulting in lower adjusted values. These results indicate that the selectivity curves produced strong adjustments in populations of Siluriformes but relatively minor differences in Characiformes. Therefore, the standard use of the nordic multi-mesh gillnet should be done cautiously, and relative selectivity should always be applied to correct the data, especially for catfish species.
This different selectivity patterns between orders (e.g., Characiformes vs. Siluriformes) found are related to the species-specific morphologies. For instance, Siluriformes have larger pectoral and dorsal spines, in addition to the presence of barbels (e.g., I. labrosus, Pimelodus sp.), and the Loricariidae have plates with a large number of structures that are very easily tangled in a mesh (e.g., R. macrops) without actually penetrating the fishing gear. Therefore, these species can be considered incidental catches in an unsuitable mesh, which results in size-specific significant differences in the probability of retention. Indeed, we observed that many large-bodied Siluriformes individuals were tangled in small mesh sizes with their spines and barbels. This could explain the low selectivity and significant difference between adjusted and unadjusted length-frequency distributions for only catfish species. In contrast, we did not observe a significant difference between adjusted and unadjusted data for Characiformes species. This indicates that Nordic multi-mesh gillnets are highly selective for this order, probably because Characiformes species do not have many spines that tangle them in smaller meshes.
Neotropical fish stocks have been highly overexploited by human-induced pressures (Agostinho et al., 2004; Link, Watson, 2019; Moi et al., 2021). In several cases, the fish population’s deterioration is not detectable until it is irreversible. Therefore, it is crucial to establish long-term monitoring and control of fish stocks in the Neotropical realm. This is the first study analyzing gillnet selectivity for the Río de la Plata basin, one of the largest estuaries on the planet. Specifically, we provide evidence that the nordic multi-mesh gillnet has a high length selectivity for Characiformes species. Despite this, an important bias for length-based population analyses was evident for Siluriformes species, which is minimized using selectivity corrections. We recommend that researchers and management agencies in Neotropical countries adopt the nordic multi-mesh gillnet for monitoring fish stocks. To increase gillnet accuracy, studies should incorporate adjustments for selectivity, preferably when monitoring Siluriformes species.
Acknowledgments
We are grateful to Universidad de la República for providing access to laboratory infrastructure. We thank Gervasio Gonzalez for logistics and data accessibility.
References
Agostinho AA, Gomes LC, Veríssimo S, Okada EK. Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Rev Fish Biol Fish. 2004; 14:11–19. https://doi.org/10.1007/s11160-004-3551-y
Albert JS, Tagliacollo VA, Dagosta F. Diversification of neotropical freshwater fishes. Annu. Rev Ecol Evol Syst. 2020; 51:27–53. https://doi.org/10.1146/annurev-ecolsys-011620-031032
Appelberg M, Berger H-M, Hesthagen T, Kleiven E, Kurkilahti M, Raitaniemi J et al. Development and intercalibration of methods in Nordic freshwater fish monitoring. Wat Air Soil Poll. 1995; 85:401–06. https://doi.org/10.1007/BF00476862
Booth AJ, Potts WM. Estimating gill-net selectivity for Labeo umbratus (Pisces: Cyprinidae), and an evaluation of using fyke-nets as a non-destructive sampling gear in small reservoirs. Fish Res. 2006; 79(1–2):202–09. https://doi.org/10.1016/j.fishres.2006.02.015
Carol BJ, García-Berthou E. Gillnet selectivity and its relationship with body shape for eight freshwater fish species. J Appl Ichthyol. 2007; 23:654–60. https://doi.org/10.1111/j.1439-0426.2007.00871.x
González-Bergonzoni I, D´Anatro A, Vidal N, Stebniki S, Tesitore G, Silva I et al. Origin of fish biomass in a diverse subtropical river: an allochthonic-supported biomass increase following flood pulses. Ecosystems. 2019; 22:1736–53. https://doi.org/10.1007/s10021-019-00370-0
González-Bergonzoni I, Silva I, Teixeira-de-Mello F, D´Anatro A, Boccardi L, Stebniki S et al. Evaluating the role of predatory fish controlling the invasion of the Asian golden mussel Limnoperna fortunei in a subtropical river. J App Ecol. 2020; 57(4):717–28. https://doi.org/10.1111/1365-2664.13573
Hamely JM. Review of gillnet selectivity. J Fish Res Board Can. 1975; 32(11):1943–69. https://doi.org/10.1139/f75-233
Hamely JM. Sampling with gill nets. Food and Agriculture Organization of the United Nations. FAO Fish Tech Pap. 1980; 33:37–53.
Hansen MJ, Madenjian CP, Selgeby JH, Helser TE. Gillnet selectivity for lake trout (Salvelinus namaycush) in Lake Superior. Can J Fish Aquat Sci. 1997; 54(11):2483–90. https://doi.org/10.1139/f97-156
Holst R, Madsen N, Moth-Poulsen T, Fonseca PA, Campos A. Manual for gillnet selectivity. European Commission. 1998; 43.
Hubert WA, Pope KL, Dettmers JM. Passive capture techniques. In: Zale AV, Parrish DL, Sutton TM, editors. Fisheries techniques, 3rd edition. American Fisheries Society, Bethesda, Maryland; 2012. p.223–65.
Klein ZB, Quist MC, Dux AM, Corsi MP. Size selectivity of sampling gears used to sample kokanee. N Am J Fish Manag. 2019; 39(2):343–52. https://doi.org/10.1002/nafm.10272
Krepper CM, García NO, Jones PD. Interannual variability in the Uruguay river basin. Int J Climatol. 2003; 23(1):103–15. https://doi.org/10.1002/joc.853
Link JS, Watson RA. Global ecosystem overfishing: Clear delineation within real limits to production. Sci Adv. 2019; 5:eaav0474. https://doi.org/10.1126/sciadv.aav0474
López-Rodriguez A, Silva I, Ávila-Simas S, Stebniki S, Bastian R, Massaro MV et al. Diets and trophic structure of fish assemblages in a large and unexplored subtropical river: the Uruguay River. Water. 2019; 11(7):1374. https://doi.org/10.3390/w11071374
Magurran AE, Baillie SR, Buckland ST, Dick JM, Elston DA, Scott EM et al. Long-term datasets in biodiversity research and monitoring: assessing change in ecological communities through time. Trends Ecol Evol. 2010; 25(10):574–82. https://doi.org/10.1016/j.tree.2010.06.016
Mildenberger T, McBride R, Wolff M. TropFishR: Fisheries stock assessment using length-frequency data (R package version 1.42). 2017. Available from: https://CRAN.Rproject.org/package=TropFishR
Millar RB. Estimating the size-selectivity of fishing gear by conditioning on the total catch. J Am Stat Assoc. 1992; 87(420):962–68. https://doi.org/10.2307/2290632
Millar RB, Holst R. Estimation of gillnet and hook selectivity using log-linear models. J Mar Sci. 1997; 54(3):471–77. https://doi.org/10.1006/jmsc.1996.0196
Millar RB, Fryer RJ. Estimating the size-selection curves of towed gears, traps, nets, and hooks. Rev Fish Biol Fish. 1999; 9:89–116. https://doi.org/10.1023/A:1008838220001
Moi D, Alves DC, Figueiredo BRS, Antiqueira PAP, Teixeira-de-Mello F, Jeppesen E et al. Non-native fishes homogenize native fish communities and reduce ecosystem multifunctionality in tropical lakes over 16 years. Sci Total Environ. 2021; 769:144524. https://doi.org/10.1016/j.scitotenv.2020.144524
Moi DA, D´Anatro A, González-Bergonzoni I, Vidal N, Silva I, Gauzens B et al. Long-term changes in multi-trophic diversity alter the functioning of river food webs. Funct Ecol. 2024. 38(8):1739–50. https://doi.org/10.1111/1365-2435.14593
Moi DA, Saito VS, Quirino BA, Alves DC, Agostinho AA, Schmitz MH et al. Human land use and non-native fish species erode ecosystem services by changing community size structure. Nat Ecol Evol. 2025; 9:801–09. https://doi.org/10.1038/s41559-025-02696-6
Neumann RM, Willis DW. Seasonal variation in gill-net sample indexes for northern pike collected from a glacial prairie lake. N Am J Fish Manag. 1995; 15(4):838–44. https://doi.org/10.1577/1548-8675(1995)015%3C0838:SVIGSI%3E2.3.CO;2
Olin M, Kurkilahti M, Peitola P, Ruuhijärvi J. The effect of fish accumulation on the catchability of multimesh gillnet. Fish Res. 2004; 68(1–3):135–47. https://doi.org/10.1016/j.fishres.2004.01.005
Pelicice FM, Bialetzki A, Camelier P, Carvalho FR, García-Berthou E, Pompeu PS et al. Human impacts and the loss of Neotropical freshwater fish diversity. Neotrop Ichthyol. 2021; 19(3):e210134. https://doi.org/10.1590/1982-0224-2021-0134
Pelicice FM, Agostinho AA, Azevedo-Santos VM, Bessa E, Casatti L, Garrone-Neto D et al. Ecosystem services generated by Neotropical freshwater fishes. Hydrobiologia. 2023; 850:2903–26. https://doi.org/10.1007/s10750-022-04986-7
Radomski P, Anderson CS, Bruesewitz RE, Carlson AJ, Borkholder BD. An assessment model for a standard gill net incorporating direct and indirect selectivity applied to Walleye. N Am J Fish Manag. 2020; 40(1):105–24. https://doi.org/10.1002/nafm.10384
Rodríguez-Climent S, Alcaraz C, Caiola N, Ibañez C, Nebra A, Muñoz-Camarillo G et al. Gillnet selectivity in the Ebro Delta coastal lagoons and its implication for the fishery management of the sand smelt, Atherina boyeri (Actinopterygii: Atherinidae). Estuar Coast Shelf Sci. 2012; 114:41–49. https://doi.org/10.1016/j.ecss.2011.09.008
Santos JA, Silva CB, Santana HS, Cano-Barbacil C, Agostinho AA, Normando FT et al. Assessing the short-term response of fish assemblages to damming of an Amazonian river. J Environ Manage. 2022; 307:114571. https://doi.org/10.1016/j.jenvman.2022.114571
Shantz AA, Ladd MC, Burkepile DE. Overfishing and ecological impacts of extirpating large parrotfish from Caribbean coral reefs. Ecol Monogr. 2020; 90(2):e01403. https://doi.org/10.1002/ecm.1403
Shoup DE, Ryswyk RG. Length selectivity and size-bias correction for the North American Standard gill net. N Am J Fish Manag. 2016; 36(3):485–96. https://doi.org/10.1080/02755947.2016.1141809
Smith BJ, Blackwell BG, Wuellner MR, Graeb BDS, Willis DW. Contact selectivity for four fish species sampled with North American Standard Gill Nets. N Am J Fish Manag. 2017; 37(1):149–61. https://doi.org/10.1080/02755947.2016.1254129
Specziár A, Erős T, György ÁI, Tátrai I, Bíro PA. Comparation between the benthic Nordic gillnet and whole water column gillnet for characterizing fish assemblages in the shallow lake Balaton. Int J Limnol. 2009; 45(3):171–80. https://doi.org/10.1051/limn/2009016
Tesfaye G, Wolff M, Taylor M. Gear selectivity of fishery target resources in Lake Koka, Ethiopia: evaluation and management implications. Hydrobiologia. 2016; 765:277–95. https://doi.org/10.1007/s10750-015-2420-0
Tonella LH, Ruaru R, Daga VS, Garcia DAZ, Júnior OBV, Lobato-de-Magalhães T et al. Neotropical freshwater fishes: a dataset of occurrence and abundance of freshwater fishes in the Neotropics. Ecology. 2023; 104(4):e3713. https://doi.org/10.1002/ecy.3713
Vandergoot CS, Kocovsky PM, Brenden TO, Liu W. Selectivity evaluation for two experimental gill-net configurations used to sample Lake Erie Walleyes. N Am J Fish Manag. 2011; 31(5):832–42. https://doi.org/10.1080/02755947.2011.623758
Authors

![]() Dieison André Moi1
Dieison André Moi1 ![]() ,
, ![]() Alejandro D’Anatro2,
Alejandro D’Anatro2, ![]() Ivan González-Bergonzoni3 and
Ivan González-Bergonzoni3 and ![]() Franco Teixeira-de-Mello4
Franco Teixeira-de-Mello4
[1] Laboratório de Interações Multitróficas e Biodiversidade, Departamento de Biologia Animal, Instituto de Biologia, Universidade Estadual of Campinas (UNICAMP), 13083-862, Campinas, SP, Brazil. dieisonparoquial@gmail.com (corresponding author).
[2] Laboratorio de Evolución, Facultad de Ciencias, Universidad de La República, Montevideo, Uruguay. passer@fcien.edu.uy.
[3] Laboratorio de Ecología Fluvial, Departamento de Ciencias Biológicas, CENUR Litoral Norte, Universidad de La República, Paysandú, Uruguay. ivan2012uy@gmail.com.
[4] Departamento de Ecología y Gestión Ambiental CURE, Universidad de la República, Maldonado, Uruguay. frantei@fcien.edu.uy
Authors’ Contribution 

Dieison André Moi: Conceptualization, Data curation, Formal analysis, Investigation, Validation, Visualization, Writing-original draft, Writing-review and editing.
Alejandro D’Anatro: Conceptualization, Data curation, Formal analysis, Supervision, Visualization, Writing-review and editing.
Ivan González-Bergonzoni: Investigation, Methodology, Resources, Validation, Visualization, Writing-review and editing.
Franco Teixeira-de-Mello: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Visualization, Writing-review and editing.
Ethical Statement
Fish handling was carried out following the CHEA (Comisión Honoraria de Experimentación Animal) protocol: “Fish fishing with multi-mesh nets” (form number 1185), associated with the project entitled “Monitoring of fish from the Uruguay and Negro rivers” approved by the CEUA (Comisión de Ética en el Uso de Animales) of the Centro Universitario Regional del Este, Universidad de la República, Uruguay.
Competing Interests
The author declares no competing interests.
Data availability statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
AI statement
The authors did not use any AI-assisted technologies in the creation of this manuscript or its figures.
Funding
DAM received a scholarship from the Fundação de Amparo à Pesquisa do Estado de São Paulo (PAPESP proc. #2022/13301–8). Sampling campaigns were financed by the UPM pulp mill environmental monitoring program. AD, IGB, and FTM received financial support by the ANII National System of Researchers (SNI) and PEDECIBA.
Supplementary Material
Supplementary material SUP
Peer Review
How to cite this article
Moi DA, D’Anatro A, González-Bergonzoni I, Teixeira-de-Mello F. Applying multi-mesh gillnet to monitor fish communities in neotropical rivers. Neotrop Ichthyol. 2026; 24(1):e250086. https://doi.org/10.1590/1982-0224-2025-0086
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2025 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted January 21, 2026
Accepted January 21, 2026
![]() Submitted May 14, 2025
Submitted May 14, 2025
![]() Epub June 05, 2026
Epub June 05, 2026