![]() Matheus Azambuja1,
Matheus Azambuja1, ![]() Ezequiel A. de Oliveira2,
Ezequiel A. de Oliveira2, ![]() Francisco de M. C. Sassi2,3,
Francisco de M. C. Sassi2,3, ![]() Viviane Nogaroto4,
Viviane Nogaroto4, ![]() Orlando Moreira-Filho2,
Orlando Moreira-Filho2, ![]() Carla S. Pavanelli5 and
Carla S. Pavanelli5 and ![]() Marcelo R. Vicari1,4
Marcelo R. Vicari1,4 ![]()
PDF: Download Here | Supplementary: Sup | Cite this article
Associate Editor: ![]() Claudio Oliveira
Claudio Oliveira
Section Editor: ![]() Bruno Melo
Bruno Melo
Editor-in-chief: ![]() José Birindelli
José Birindelli
Abstract
Parodontidae é uma família de peixes Neotropicais agrupada nos gêneros Saccodon, Parodon e Apareiodon. Apesar das similaridades morfológicas em alguns pares de espécies, marcadores genéticos e cromossômicos têm sido úteis para resolver incertezas taxonômicas em Parodontidae. Representantes de Parodon da bacia Amazônica estão entre as espécies com características morfológicas sutis que exigem identificação precisa. Assim, este estudo comparou espécies de Parodon usando marcadores citogenéticos e genéticos, com o objetivo de delimitar molecularmente as espécies. Adicionalmente, duas populações de Parodon da bacia Amazônica, morfologicamente identificadas como Parodon sp. (rio do Peixe) e Parodon cf. buckleyi (rio Branco) tiveram caracterização citogenética e molecular. Ambas, Parodon sp. e P. cf. buckleyi apresentaram 2n = 54 e marcadores cromossômicos similares a Parodon nasus, mas com diferenças espécie-específicas. A distância genética, a análise Bayesiana e os métodos moleculares de delimitação de espécies reconheceram P. guyanensis, P. pongoensis, P. hilarii, P. caliensis, P. suborbitalis, P. magdalenensis, P. apolinari, P. nasus, P. cf. buckleyi e Parodon sp. como espécies válidas. A análise integrativa dos dados citogenéticos e moleculares indica Parodon sp. como uma Unidade Taxonômica Operacional molecular. Esses resultados revelam uma diversidade oculta no gênero Parodon e sugerem que o “complexo P. suborbitalis” não é monofilético, com grupos distintos em uma estrutura biogeográfica.
Palavras-chave: Cromossomos, DNA Barcoding, FISH, MOTU, Taxonomia integrativa.
Introduction
The Amazon basin is the largest river basin on Earth, covering approximately 6.3 million km2 (Milliman, Farnsworth, 2011). It supports the highest freshwater biodiversity (Tisseuil et al., 2013), with approximately 2,400 valid species (Jézéquel et al., 2020), accounting for nearly 15% of all known freshwater fish species worldwide (Tedesco et al., 2017). However, the true extent of fish diversity in the Amazon basin is likely underestimated, as numerous new species continue to be described every year (Antonelli et al., 2018; Jézéquel et al., 2020).
Parodontidae is a Neotropical fish family comprising 32 valid species (Fricke et al., 2025), grouped into three genera: Parodon Valenciennes, 1849, Saccodon Kner, 1863, and Apareiodon Eigenmann, 1916 (Pavanelli, 2003). This family has a broad geographic range in South America and Panama, except for some coastal Atlantic basins and Patagonia (Pavanelli, Britski, 2003). Their species are classified into genera based on characters such as jaw lateral teeth and the number of undivided rays in the pectoral fin (Pavanelli, 2003). For species identification, body coloration patterns with a single regular black longitudinal stripe or vertical bars can be used, along with the shape and number of cusps of the symphysial tooth, among others (Pavanelli, 1999).
Among Parodon species, the “P. suborbitalis complex” was initially proposed by Pavanelli (1999) based on morphological similarities, particularly body color patterns featuring a black zigzag lateral band on a light background, despite inhabiting different river basins. The species complex originally comprised five species: Parodon bifasciatus Eigenmann, 1912, P. buckleyi Boulenger, 1887, P. hilarii Reinhardt, 1867, P. nasus Kner, 1859, and P. suborbitalis Valenciennes, 1850. In a taxonomic review of Parodon species from Colombia, Londoño-Burbano et al. (2011) agreed with the previously proposed “P. suborbitalis complex” but suggested a new arrangement. They included one species previously excluded by Pavanelli (1999), Parodon carrikeri Fowler, 1940, with a black lateral band with a diffuse zigzag on a dark background, in addition to P. alfonsoi Londoño-Burbano, Román-Valencia & Taphorn, 2011, P. atratoensis Londoño-Burbano, Román-Valencia & Taphorn, 2011, and P. magdalenensis Londoño-Burbano, Román-Valencia & Taphorn, 2011, which were described after the earlier work. The species complex now encompasses nine of the fourteen valid Parodon species. Despite detailed morphological descriptions, species identification in the complex relies on average character values with some overlap, making geographic origin an important consideration for identification (see Pavanelli, 1999, and Londoño-Burbano et al.,2011 for further details). Most of “P. suborbitalis complex” representatives lack genetic and cytogenetic data, which could contribute to resolving this group’s classification.
Six Parodon species are documented in Brazilian river basins: P. bifasciatus, P. buckleyi, P. hilarii, P. moreirai Ingenito & Buckup, 2005, P. nasus, and P. pongoensis (Allen, 1942) (Pavanelli, 2003; Londoño-Burbano et al., 2011). In the Brazilian Amazon River basin, P. bifasciatus is found in the northern region, while P. buckleyi occurs in the southern part of the basin (Pavanelli, 2003; Londoño-Burbano et al., 2011). However, according to Pavanelli (1999), Brazilian populations of P. buckleyi from the Machado River sub-basin in Rondônia state are considered isolated representatives of the species, due to numerous waterfalls in the Machado River, which likely serve as geographical barriers to the gene flow. The P. buckleyi specimens analyzed by Pavanelli (1999) from the Machado River sub-basin were more robust, with projections of the main black longitudinal band not very evident, in addition to a lower average number of cusps in the premaxillary teeth, in comparison to P. buckleyi from Ecuador and Peru.
Cytogenetically, Parodontidae has a conserved diploid chromosome number (2n) of 54 among the species (Bellafronte et al., 2011). However, their species can be distinguished according to chromosomal characteristics: karyotypic formula, differential accumulation and number of heterochromatic bands, presence or absence of sex heteromorphic chromosomes, number of sites of rDNAs and snDNAs, distribution and sites number of the pPh2004 satellite DNA, and the WAp repetitive fraction distribution on the karyotypes (Bellafronte et al., 2011; Schemberger et al., 2011; Ziemniczak et al., 2014; Traldi et al., 2016, 2020; Santos et al., 2019; Nirchio et al., 2021; Azambuja et al., 2022a,b, 2023). The occurrence of ZW sex chromosome differentiation in some species is a significant feature in the Parodontidae fish’s diversification (Schemberger et al., 2011, 2019; Oliveira et al., 2024, 2025). Species lacking heteromorphic sex chromosomes or with sex chromosomes at different levels of differentiation, such as proto-sex chromosomes, ZW sex chromosomes, and with multiple systems of heteromorphic sex chromosomes (ZZ/ZW1W2) were observed in the group (Schemberger et al., 2011; Bellafronte et al., 2012; Traldi et al., 2016, 2020; Nascimento et al., 2018; Santos et al., 2019; Nirchio et al., 2021; Wolf et al., 2024). For species in “P. suborbitalis complex”, just P. hilarii and P. nasus were assessed by cytogenetic studies (Centofante et al., 2002; Vicente et al., 2003). However, remarkable karyotype differences are found, i.e., P. hilarii, 2n = 54 m/sm, ZZ/ZW sex chromosome system, non-syntenic 45S and 5S rDNA sites, and 14–16 pPh2004 satellite DNA sites; P. nasus, 2n = 54, 48m/sm + 6st, proto-sex chromosome, 8 pPh2004 satellite DNA sites, among others (Bellafronte et al., 2011; Schemberger et al., 2011).
The Molecular Operational Taxonomy Unit (MOTU) represents clusters of DNA sequences that differ from each other by more than an established threshold of similarity and are therefore proposed as operational taxonomic units (Floyd et al., 2002; Blaxter, 2004). For this purpose, molecular analyses using the Cytochrome c Oxidase subunit I (COI) gene can help in the initial delimitation of species (Hebert et al., 2003) and have been widely used in the identification and delimitation of Neotropical fishes (Serrano et al., 2018; Ramirez et al., 2020; Morais-Silva et al., 2023; Souza et al., 2023; Almeida et al., 2024), including Parodontidae (Bellafronte et al., 2013; Nascimento et al., 2018; Santos et al., 2019; Traldi et al., 2020), as well as in fish complex groups (Terán et al., 2020; Limeira Filho et al., 2024; Fernandes et al., 2025). Different modern methods for delimiting species have been proposed for single-locus analyses (Pons et al., 2006; Fujisawa, Barraclough, 2013; Zhang et al., 2013; Puillandre et al., 2021). However, recent analyses have shown that combining other methods with integrative taxonomy yields more representative results for the lineages (Tang et al., 2014; Kapli et al., 2017; Luo et al., 2018; Serrano et al., 2018; Traldi et al., 2020).
The integration of chromosome markers (cytotaxonomy) and COI sequence analysis has been utilized as a tool to address taxonomy uncertainties among parodontids, especially in species with similar morphological and chromosomal traits (Bellafronte et al., 2013; Nascimento et al., 2018; Santos et al., 2019; Traldi et al., 2020; Azambuja et al., 2022a). Regarding species in the Parodon genus, with special attention to those within the “P. suborbitalis complex” a more comprehensive collection of cytogenetic and genetic data is still required. To elucidate the genetic diversity and taxonomic boundaries within Parodon, we cytogenetically and molecularly characterized two Amazonian Parodon species exhibiting morphological traits, particularly body color pattern, consistent with the “P. suborbitalis complex”. These data were integrated with chromosomal and mitochondrial COIinformation from additional congeners, both within and outside the complex, and from different hydrographic regions of the Amazon basin. By applying molecular species delimitation methods within an integrative taxonomic framework, we aimed to assess the occurrence of MOTUs and refine the limits of the “P. suborbitalis complex”.
Material and methods
Biological samples. Fifteen individuals (1 female, 8 males, and 6 sex undetermined) previously identified as Parodon sp. (Fig. 1A) from the Peixe River, Amazon basin, Tapajós River sub-basin, Pará State, Brazil, 08°39’15.5”S 55°09’24.3”W (Fig. 1C), and thirty-two individuals (10 females and 22 males) identified as P. cf. buckleyi (Fig. 1B) from the Branco River, Amazon basin, Madeira River sub-basin, Rondônia State, Brazil, 11°55’51.1”S 62°09’09.4”W (Fig. 1C) were analyzed.
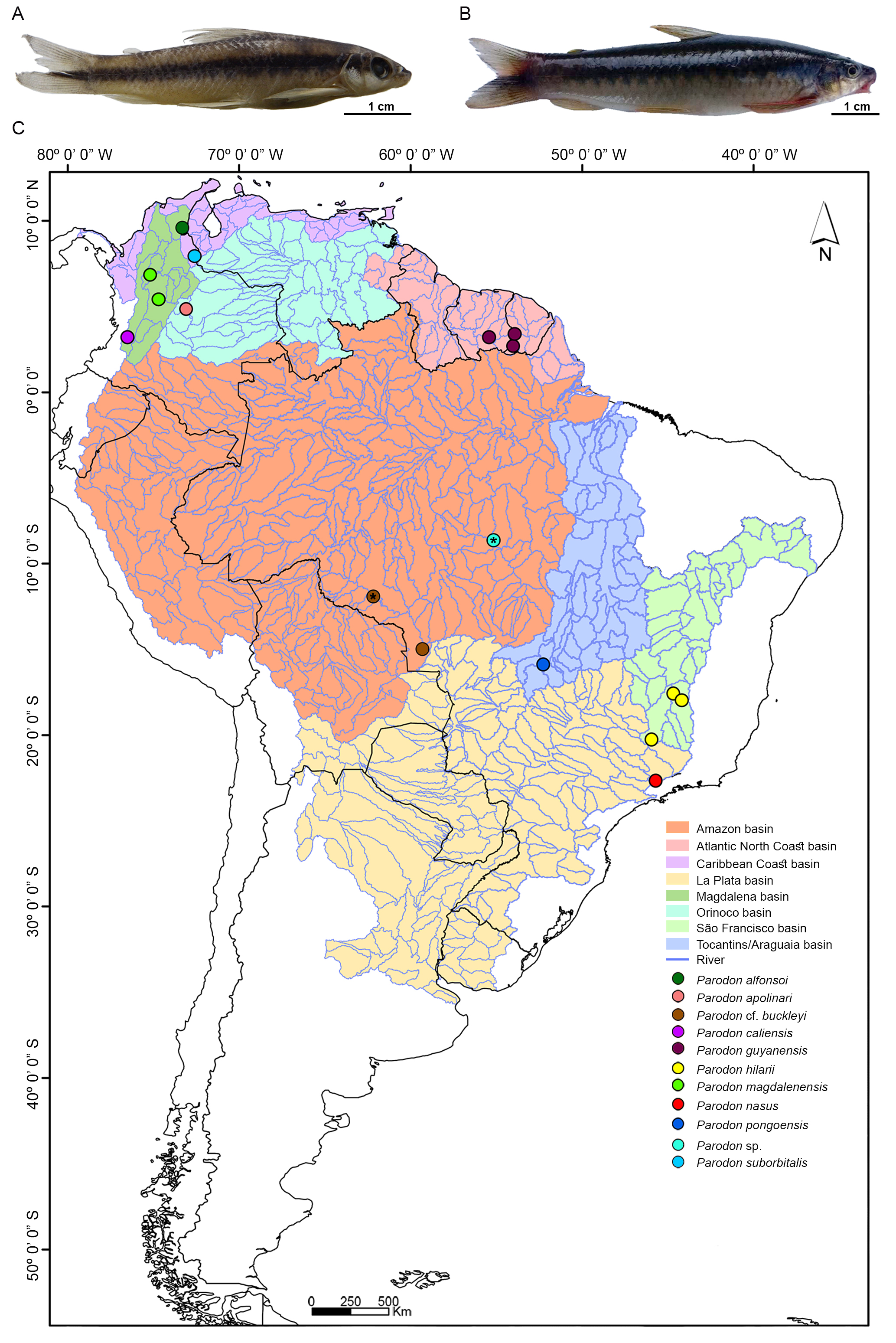
FIGURE 1| A. Parodon sp. and B. Parodon cf. buckleyi. C. Partial map of South America showing collection sites and river basins for analyzed Parodon species. *Correspond to Parodon sp. and P. cf. buckleyi populations analyzed cytogenetically.
Specimen identification was conducted based on geographic origin and color pattern, as this species complex cannot currently be distinguished solely by morphological traits. Specimens were deposited as vouchers in the Ichthyological Collection at Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (Nupelia) of Universidade Estadual de Maringá, Paraná State, Brazil (NUP 22248, 23214, and 23216). Eight liver samples of Parodon pongoensis from the Taquaralzinho River, Tocantins-Araguaia basin, Mato Grosso State, Brazil, 15º53’28”S 52º14’56”W (Fig. 1C) and four of P. hilarii from the Araras River, São Francisco basin, Minas Gerais state, Brazil, 20°16’15”S 45°55’39”W (Fig. 1C) from the tissue bank of the Laboratório de Citogenética Evolutiva of the Universidade Federal de São Carlos were used for the molecular analyses. In addition, one tissue sample of P. buckleyi from the Pindaituba River, Amazon basin, Madeira River sub-basin, Mato Grosso State, Brazil, 15°00’41”S 59°17’18”W (Fig. 1C), from the ichthyological collection of the Museu de Ciências e Tecnologia of the Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre (PUCRS) (Voucher number: MCP 38672) was used for comparisons with our P. cf. buckleyi samples and included in the molecular analyses.
Chromosomal preparations and classical cytogenetics analysis. Mitotic chromosomes were obtained from the animals’ kidneys as described by Bertollo et al. (2015). Chromosomes were stained with 5% Giemsa in phosphate buffer (pH = 6.8) to determine 2n, fundamental number (FN), and karyotype formula. Heterochromatic regions were identified by C-banding, as described by Sumner (1972). Metaphases were analyzed and photographed using a bright-field microscope (Olympus BX43) coupled to a DP72 CCD camera (Olympus). Homologous chromosomes were paired and organized into karyotypes following Levan et al. (1964). At least 30 metaphases of each species were analyzed to confirm the 2n and heterochromatic bands.
Molecular cytogenetics analysis. Fluorescence in situ hybridization (FISH) was performed following Pinkel et al. (1986), and six sequences of repetitive DNAs were physically mapped in Parodon sp. and P. cf. buckleyi: 18S and 5S rDNAs, pPh2004 satellite DNA, (GATA)n, and the transposable elements Helitron and Tc1-Mariner. The 18S probe was obtained according to Hatanaka, Galetti Jr. (2004), and labeled with Biotin-16-dUTP (Biotin-Nick Translation Mix; Roche Applied Science). The 5S rDNA sequence was obtained and labeled with digoxigenin-11-dUTP (DIG-11-dUTP; Jena Bioscience) by polymerase chain reaction (PCR) using genomic DNA from P. nasus and the primers 5SA and 5SB (Martins, Galetti, 1999). The pPh2004 probe was prepared according to Vicente et al. (2003) and labeled with digoxigenin-11-dUTP (Dig Nick Translation Mix; Roche Applied Science) and Biotin-16-dUTP (Biotin-Nick Translation Mix; Roche Applied Science). The (GATA)n probe was obtained according to Traldi et al. (2013), and the Helitron and Tc1-Mariner probes were obtained according to Schemberger et al. (2019) and labeled with digoxigenin-11-dUTP (Dig Nick Translation Mix; Roche Applied Science). FISH was performed under stringent conditions (~80%) (300 ng of each probe, 50% formamide, 10% dextran sulfate, 2xSSC, 37°C for 16 h). Signal detection was carried out using Streptavidin Alexa Fluor 488 (Molecular Probes) and anti-digoxigenin-rhodamine (Roche Applied Science). Chromosomes were stained with DAPI (0.2 µL/mL) present in Vectashield mounting medium (Vector) and analyzed in an epifluorescence microscope (Leica DM 2000) coupled to a DFC3000 G CCD camera (Leica). At least 20 metaphases per probe were analyzed to determine FISH signals.
Molecular analysis. Genomic DNA of five individuals of Parodon sp., six of P. cf. buckleyi, eight of P. pongoensis, and four of P. hilarii were extracted from liver samples following the CTAB (cetyltrimethylammonium bromide) method of Murray, Thompson (1980). DNA was used to amplify the barcode region of the COI by PCR using the primers Fish F1and Fish R1 (Ward et al., 2005). Reaction mix contained: 1x Taq Reaction buffer (200 mM Tris pH 8.4, 500 mM KCl), 1 mM MgCl2, 0.2 mM dNTPs, 0.4 µM of each primer, 1 U Taq DNA polymerase (Invitrogen), and 40 ng of DNA. The following reaction program was used: initial denaturation for 10 min at 94°C, 35 cycles of 94°C for 1 min, 54.5°C for 45 sec, and 72°C for 90 sec, and a final extension at 72 °C for 10 min. The PCR products were purified using the Illustra GFX PCR DNA and Gel Band Purification kit (GE Healthcare) and sequenced on an ABI-prism 3500 Genetic Analyzer (Applied Biosystems).
Electropherograms were reviewed, the nucleotide sequences were corrected in Geneious v. 7.1.9 (Kearse et al., 2012) and deposited in GenBank. COI sequences of the species Parodon alfonsoi (1), P. apolinari Myers, 1930 (3), P. caliensis Boulenger, 1895 (1), P. guyanensis Géry, 1960 (9), P. hilarii (3), P. magdalenensis (3), P. nasus (5) and P. suborbitalis (1) were mined from the Barcode of Life Database (BOLD) (Ratnasingham, Hebert, 2007) and GenBank database (Benson et al., 2013) (Tab. S1), and were aligned with the sequences of Parodon sp., P. cf. buckleyi, P. hilarii,and P. pongoensis (this study) using the algorithm ClustalW, integrated with the software Geneious. Sequences were separated into groups, and genetic distances were calculated using MEGA X (Kumar et al., 2018), under the Kimura-2-parameters (K2P) evolution model with 1,000 bootstrap replications. The number of haplotypes was verified in DnaSP v5 (Librado, Rozas, 2009), and a haplotype network was generated using the Minimum Spanning Network criterion (Bandelt et al., 1999) in PopArt v. 1.7 (Leigh, Bryant, 2015).
One Leporinus piau Fowler, 1941 COI sequence (HM405030.1) was used to root the trees and was aligned with Parodon’s sequences using ClustalW. Sequences were submitted to jModelTest 2 (Darriba et al., 2012) using corrected Akaike information criterion (AICc) to select the best-fit nucleotide evolution GTR + G model for downstream analyses.
Four methods of species delimitation were applied: (a) General mixed Yule-coalescent (GMYC) (Pons et al., 2006; Fujisawa, Barraclough, 2013); (b) Bayesian Poisson Tree Processes (bPTP) (Zhang et al., 2013); (c) Automatic Barcode Gap Discovery (ABGD) (Puillandre et al., 2012); (d) Assemble Species by Automatic Partitioning (ASAP) (Puillandre et al., 2021). For GMYC analysis, an ultrametric gene tree was inferred in Beast v. 2.6.1 (Bouckaert et al., 2019). The GTR + G substitution model, a Strick Clock and the Yule model prior were employed. The Markov chains included 100,000,000 generations, storing trees every 10,000 generations, to obtain a total of 10,001 trees. Tracer v. 1.6 (Rambaut et al., 2014) was used to examine the average standard deviation of split frequencies and the convergence of MCMC searches, with >200 considered an appropriate effective sample size. The first 1,000 trees were discarded as burn-in, and 9,001 trees were summarized in the Maximum Clade Credibility tree (MCC) from the posterior distribution in TreeAnnotator v. 2.6 (Bouckaert et al., 2019). The tree was imported into the R software (R Development Core Team, 2013), and a species delimitation test was made with the SPLITS package (Species’ Limits by Threshold Statistics; http://r-forge.r-project.org/projects/splits/), using the single threshold method. For bPTP analysis, a Bayesian inference tree was generated in MrBayes 3.2 program (Huelsenbeck, Ronquist, 2001) applying 100,000,000 iterations of MCMC, sampling trees every 10,000 generations and burn-in of 10,000,000, and used as an input file in the bPTP web server (https://species.h-its.org/ptp/). 500,000 MCMC generations run (thinning = 500), and the other parameters were kept at their default values. The alignment generated for the sequences was used as an input file for the delimitation of species in the ABGD web server (https://bioinfo.mnhn.fr/abi/public/abgd/abgdweb.html) and in the ASAP web server (https://bioinfo.mnhn.fr/abi/public/asap/asapweb.html), the Kimura (K80) TS/TV was selected for distance mode, assuming that Ts and TV have different rates, and the others parameters were kept at default, as suggested by Puillandre et al. (2012, 2021).
Results
Cytogenetic data. The 2n of 54 was verified for Parodon sp. and P. cf. buckleyi, but with distinct karyotypic formula: 48 m/sm + 6 st and FN = 108 for Parodon sp. (Figs. 2A–B), and 50 m/sm + 4 st and FN = 108 for P. cf. buckleyi (Figs. 2E–F) (Tab. 1). Heterochromatin was preferentially located in the centromeric and terminal portions of the chromosomes in Parodon sp. and P. cf. buckleyi (Figs. 2C–D, G–H). Interstitial heterochromatic blocks were located at the q arms of pairs 6, 13, 26, and 27 in Parodon sp. (Figs. 2C–D) and at the q arms of pairs 8, 13, and 27 in P. cf. buckleyi (Figs. 2G–H).

FIGURE 2| Karyotypes of Parodon sp. (A–D) and Parodon cf. buckleyi (E–H) after conventional Giemsa-staining (A–B, E–F) and C-banding (C–D, G–H). A, C, E and G correspond to male individuals, and B, D, F and H correspond to female individuals. Scale bar = 10µm.
TABLE 1 | Cytogenetic and chromosomal markers data available for Parodon species. ♂: male; ♀: female; m: metacentric; sm: submetacentric; st: subtelocentric; p: short arm; q: long arm; Ref.: References. 1 – Moreira-Filho et al. (1993); 2 – Jesus, Moreira-Filho (2000); 3 – Moreira-Filho et al. (1984); 4 – Moreira-Filho et al. (1985); 5 – Vicente et al. (2001); 6 – Vicente et al. (2003); 7 – Centofante et al. (2002); 8 – Bellafronte et al. (2005); 9 – Bellafronte et al. (2011); 10 – Schemberger et al. (2011); PS: Present study.
Species | Karyotypic formulae | Sex chromosomes system | 18S rDNA sites | 5S rDNA sites | pPh2004 sites | Ref. |
Parodon hilarii | 54 m/sm ♂ 53 m/sm + 1 st ♀ | ZZ/ZW | 16q m (terminal) | 11p m (proximal) | 2pq m, 4p m, 9q m, 15pq sm, 21p m, 25q m, Zq sm, and Wp sm (all sites terminals) | 1, 2, 5, 6, 10 |
Parodon moreirai | 54 m/sm ♂♀ | ZZ/ZW | 15q m (terminal) | 9p m (proximal) | 9q m, Zq sm, and Wp m (all sites terminals) | 7, 10 |
Parodon nasus | 48 m/sm + 6 st ♂♀ | Proto-sex chromosome | 25q st (terminal) | 25p st (proximal) | 6q sm (terminal and interstitial), and 13q sm, 26q st and 27q st (terminals) | 2, 3, 4, 5, 7, 8, 9, 10 |
Parodon pongoensis | 50 m/sm + 4 st ♂♀ | Proto-sex chromosome | 2q m (terminal) | 9 m (proximal) | 13q m (terminal) | 2, 5, 9, 10 |
Parodon sp. | 48 m/sm + 6 st ♂♀ | Proto-sex chromosome | 25q st (terminal) | 25p st (proximal) | 6q sm, 26q st, and 27q (all sites terminals) | PS |
Parodon cf. buckleyi | 50 m/sm + 4 st ♂♀ | Proto-sex chromosome | 26q st (terminal) | 26p st (proximal) | 8 sm (centromeric and q terminal) and 27q st (terminal) | PS |
Double-FISH using 18S rDNA and 5S rDNA probes showed a syntenic arrangement in Parodon sp. and P. cf. buckleyi karyotypes, with 45S rDNA located on the 25q terminal region, and 5S rDNA located on the 25p proximal region in Parodon sp. (Fig. 3A), and 45S rDNA located on the 26q terminal region, and 5S rDNA located on the 26p proximal region in P. cf. buckleyi (Fig. 3B). The pPh2004 satellite DNA was located on the q terminal regions of submetacentric pair 6, and subtelocentric pairs 26 and 27 in Parodon sp. (Fig. 3C). For P. cf. buckleyi, pPh2004 satellite DNA was located on the centromeric and q terminal region in the submetacentric pair 8, and on the q terminal region in the subtelocentric pair 27 (Fig. 3D).
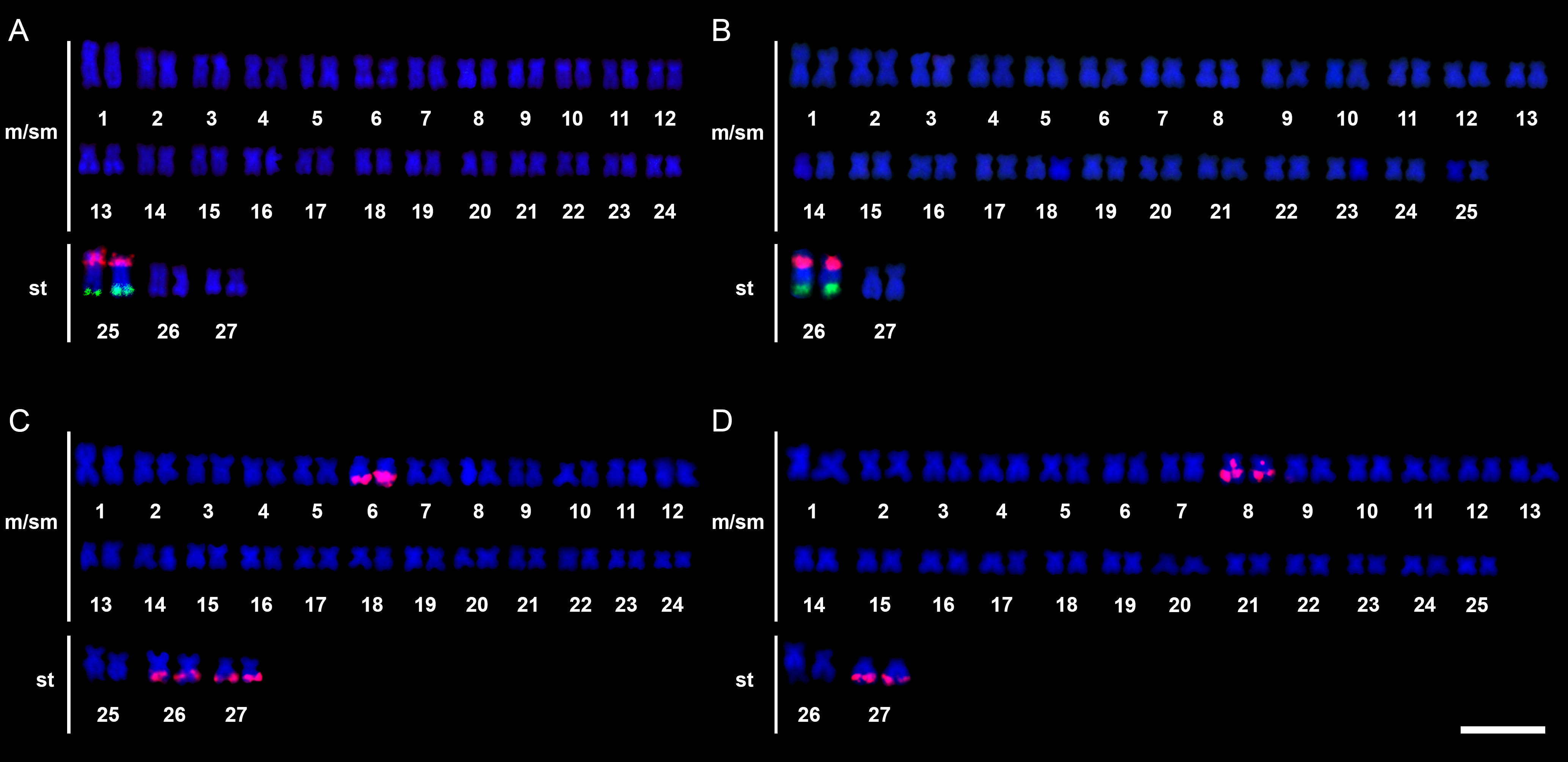
FIGURE 3| Karyotypes of Parodon sp. (A, C) and Parodon cf. buckleyi (B, D) submitted to fluorescence in situ hybridization using 18S (green signals) and 5S (red signals) rDNA probes (A–B), and pPh2004 satellite DNA probe (C–D). Scale bar = 10 µm.
(GATA)n showed an evident block at the p arm of a submetacentric chromosome pair (pair 9) and some dispersed signals in Parodon sp. chromosomes (Fig. 4A). In P. cf. buckleyi, evident signals of (GATA)n were located on the p arms of a submetacentric (pair 9) and a subtelocentric (pair 25) chromosome, with dispersed signals in other chromosomes (Fig. 4B). The Tc1-Mariner repetitive DNA was located on the terminal regions of almost all chromosomes in Parodon sp. and P. cf. buckleyi (Figs. 4C–D). Helitron sequence was located on the q distal regions in chromosome pair 6 in Parodon sp. (Fig. 4E), and in chromosome pairs 8 and 27 in P. cf. buckleyi (Fig. 4F).
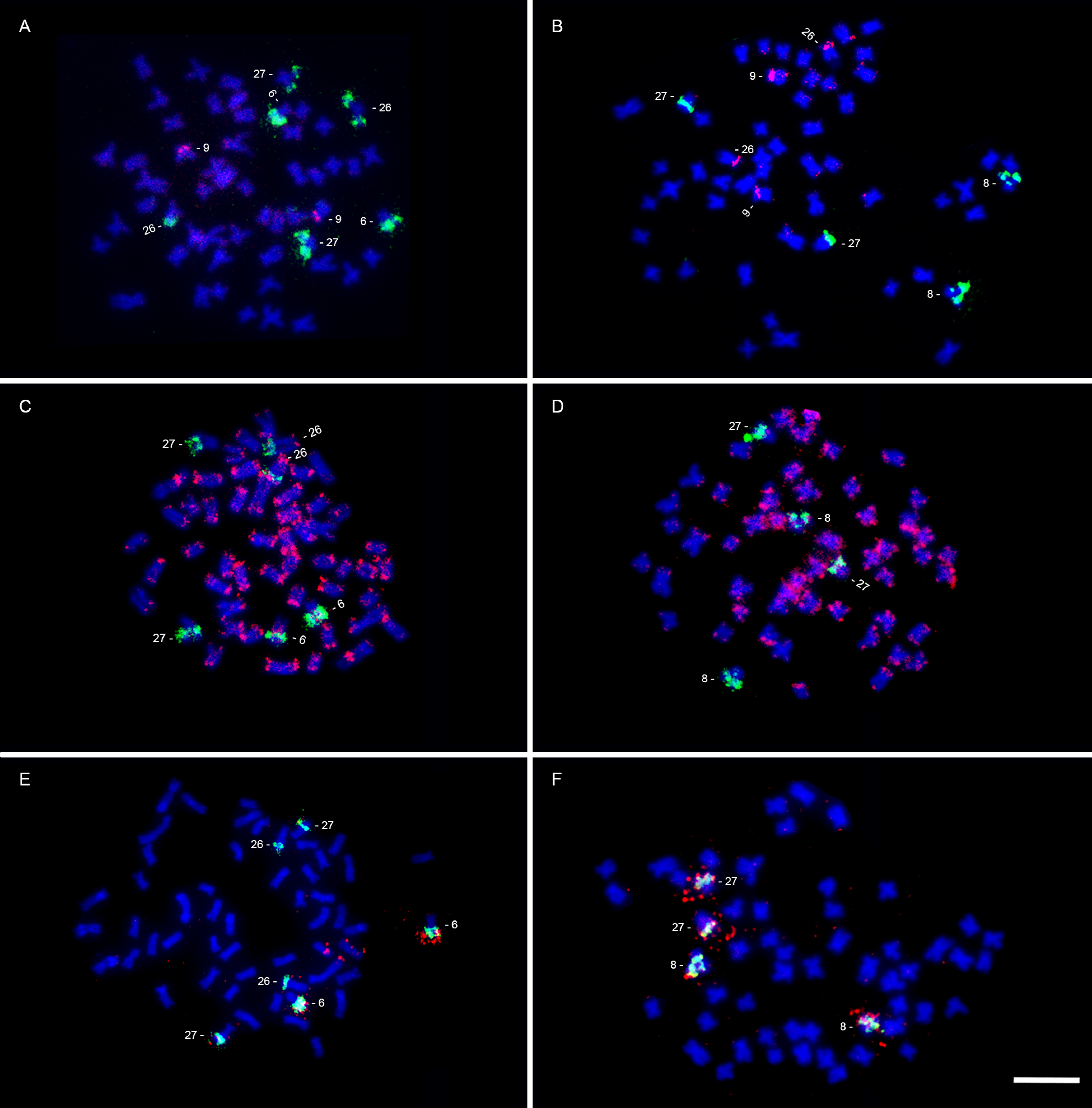
FIGURE 4| Parodon sp. (A, C, E) and Parodon cf. buckleyi (B, D, F) metaphases submitted to double FISH using pPh2004 satellite DNA probe (green signals; A–F) and (GATA)n sequence (red signals; A–B), Tc1-mariner probe (red signals; C–D), and Helitron probe (red signals; E–F). Scale bar = 10 µm.
Molecular analysis. Partial COI gene sequences were obtained for Parodon sp., P. cf. buckleyi, P. hilarii,and P. pongoensis (GenBank accession numbers: PV834389–PV834411). All sequences were of high quality and showed no evidence of indels, deletions, or stop codons. Sequences were aligned with the Parodon sequences retrieved from BOLD and GenBank, and a 602 bp matrix comprising 49 sequences with 167 polymorphic sites was generated. Twenty haplotypes were identified in the matrix: Parodon apolinari, P. caliensis, P. guyanensis, P. hilarii, and P. suborbitalis showed unique haplotypes, P. nasus presented two haplotypes, while four different haplotypes for each one of the populations of Parodon sp., P. cf. buckleyi,and P. pongoensis were observed (Fig. 5A). Parodon alfonsoi and P. magdalenensis shared a unique haplotype (Fig. 5A).
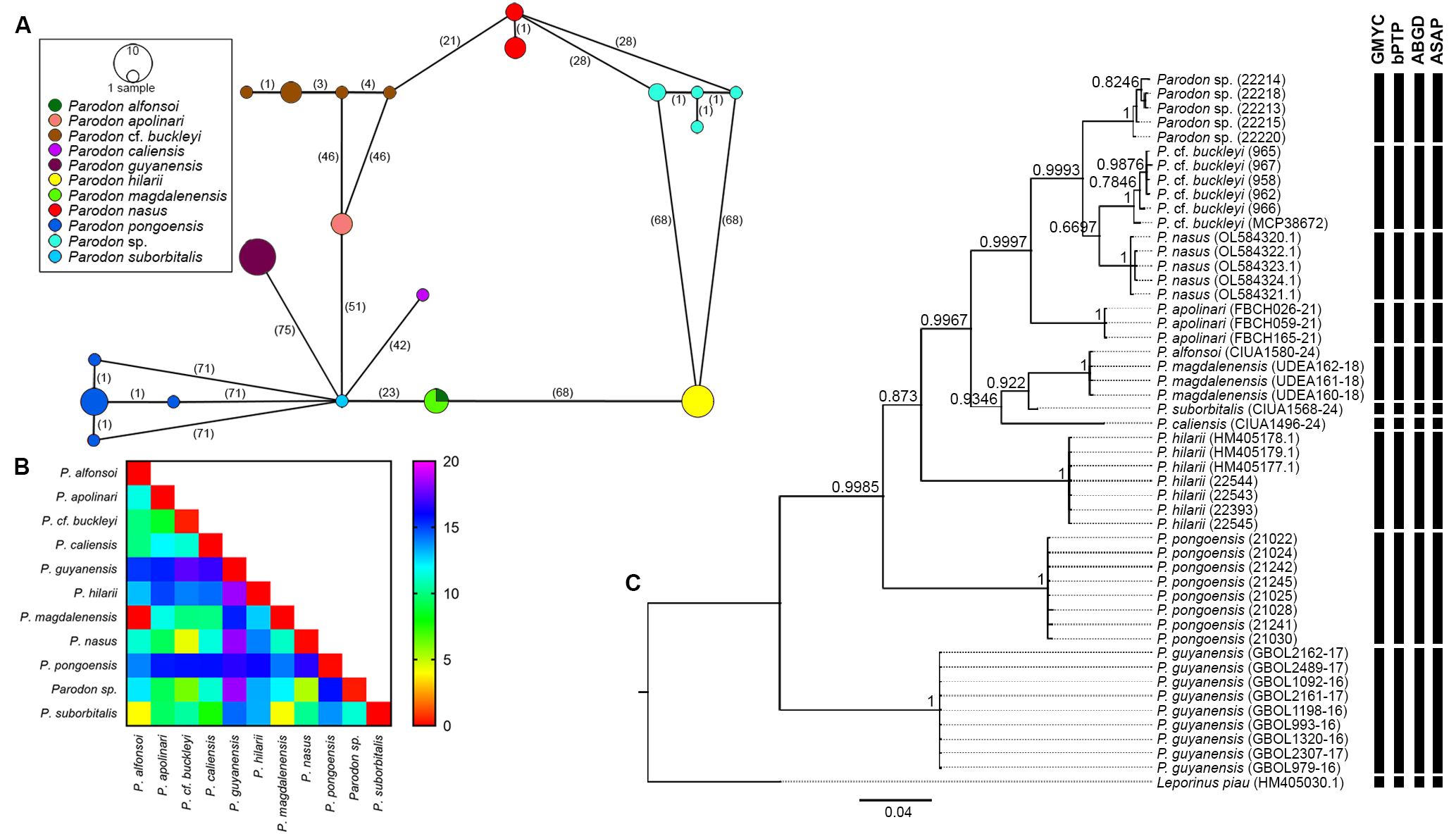
FIGURE 5| Molecular data of Parodon species. A. Haplotype network of COI sequences. B. Heatmap of genetic distance among Parodon species (values were presented in %). C. Bayesian inference tree showing phylogenetic relationships and molecular species delimitation (vertical black bars) (Numbers on the branches correspond to posterior probability).
The intraspecific genetic distance ranged from 0 to 0.46% and the interspecific genetic distance ranged from 0 to 18.4% among the Parodon species (Fig. 5B; Tab. S2). The phylogenetic tree demonstrated monophyly for the nine Parodontidae species analyzed (Fig. 5C). The recovered topology showed the occurrence of eight groups: (1) Parodon guyanensis;(2) P. pongoensis; (3) P. hilarii; (4) P. caliensis; (5) P. suborbitalis; (6) P. magdalenensis + P. alfonsoi; (7) P. apolinari;(8) P. cf. buckleyi, P. nasus and Parodon sp. (Fig. 5C). Group 8 also indicated that P. cf. buckleyi is the sister group to P. nasus, with Parodon sp. forming a sister lineage to both.
The four molecular methods of species delimitation tested recovered eleven MOTUs (outgroup included): Parodon apolinari, P. cf. buckleyi, P. caliensis, P. guyanensis, P. hilarii, P. magdalenensis + P. alfonsoi, P. nasus, P. pongoensis, Parodon sp., P. suborbitalis, and Leporinus piau (Fig. 5C). The GMYC analysis suggested eight clusters (confidence interval 8–8) and the number of eleven entities (confidence interval 11–12; likelihood of null model: 316.5217; maximum likelihood of GMYC model: 339.2739; likelihood ratio: 45.50428; LR test: 1.314836e-10***; threshold time: -0.0058). The ML solution for bPTP also identified 11 species for the analyzed tree. Eight partitions were observed with the ABGD method: one partition found seventeen groups, while the other seven found eleven groups. For partition 5, eleven species (outgroup included) were observed (prior maximal distance P = 0.007743). The best partition identified by the ASAP method recognized eleven species (ASAP-score: 1.50; p-value: 8.93e-03; W: 2.51e-03; Threshold dist.: 0.022016; outgroup included).
Discussion
The integration of multiple lines of evidence (morphology, cytogenetics, and DNA barcoding, among others) within a unified framework underpins the concept of integrative taxonomy (Dayrat, 2005). This approach, combining morphological, cytogenetic, and molecular data, has proven decisive for delimiting some Neotropical characiforms (Ramirez et al., 2017; Gavazzoni et al., 2024). Species in the “P. suborbitalis complex” lack reliable external diagnostic characters and have been grouped primarily by coloration,especially the shape of the lateral stripe,and geographic distribution (Pavanelli, 2003; Londoño-Burbano et al.,2011). Within thiscomplex, species exhibit overlaps in morphological traits that typically serve as diagnostic characters for other Parodontid species, such as the number of pored scales along the lateral line, premaxillary cusps, and preanal scales (Pavanelli, 2003; Londoño-Burbano et al., 2011). Comprehensive morphological assessments of Parodon sp. and P. cf. buckleyi remain necessary to clarify their taxonomic placement. Notably, Parodon cf. buckleyi may also represent a distinct MOTU in Parodon, given that the type locality of P. buckleyi is in Ecuador, and populations from the Machado River sub-basin appear isolated (Pavanelli, 1999). In the absence of decisive external diagnostic, cytogenetic techniques (cytotaxonomy) and DNA barcoding have been effective tools for species delimitation in Parodontidae (Bellafronte et al., 2011, 2013; Schemberger et al., 2011; Azambuja et al., 2023).
Parodon sp. and P. cf. buckleyi show a karyotype quite similar, with the same 2n and FN, and no heteromorphic sex chromosomes, indicating a close relationship between these lineages. Nevertheless, differences were identified in the karyotype formula, heterochromatin accumulation, and the in situ localization of repetitive DNAs. Within Parodon, cytogenetic studieshave been limited to P. hilarii, P. moreirai, P. nasus, and P. pongoensis. All these species show 2n = 54 and FN = 108, however, heteromorphic sex chromosomes (ZZ/ZW) have been described for P. hilarii and P. moreirai, while a pair of proto-sex chromosomes has been proposed for P. nasus and P. pongoensis (Tab. 1). The cytogenetic analysis reveals that Parodon sp. and P. cf. buckleyi have chromosomal features similar to P. nasus (Tab. 1). The syntenic condition for 45S and 5S rDNAs (i.e., 45S rDNA located on the q arm and 5S rDNA located on the p arm of the first subtelocentric chromosome pair) observed in Parodon sp. and P. cf. buckleyi has been proposed as an apomorphic chromosomal condition in P. nasus (Bellafronte et al., 2005, 2011; Azambuja et al., 2022a). Thus, the homeology of the rDNA-bearing chromosome pair designates this condition as a synapomorphy shared by Parodon sp., P. nasus,and P. cf. buckleyi within Parodontidae, and differentiates these species from P. hilarii, P. moreirai,and P. pongoensis.
The number and chromosomal locations of the pPh2004 satellite DNA sites are other chromosomal characteristics that differ among Parodontidae karyotypes (Bellafronte et al., 2011; Schemberger et al., 2011). However, some conservation is observed in homeologous chromosomal loci of closely related species, which is useful for karyotype-based species delimitation (Schemberger et al., 2011; Traldi et al., 2020). Differences in the distribution of pPh2004 satellite DNA are evident between Parodon sp. and P. cf. buckleyi, suggesting interspecific divergence between these taxa. Furthermore, both the number and location of pPh2004 satellite DNA sites show interspecific divergence when compared to other Parodon species such as P. pongoensis, P. nasus, P. moreirai,and P. hilarii (Tab. 1). Satellite DNAs loci have been considered hotspots for chromosomal rearrangements in many karyotypes due to their dynamic and fast-evolving nature (Paço et al., 2013; Gatto et al., 2018; Moraes et al., 2023; Ribas et al., 2026). The pPh2004 site, observed in the centromeric region of chromosome pair 8 in P. cf. buckleyi, represents a species-specific marker within Parodon species, which may have resulted from ectopic recombination or a chromosomal inversion involving pPh2004 sequences.
Chromosome painting with the W heterochromatic fraction of Apareiodon sp. (WAp probe) in combination with the pPh2004 satellite DNA was employed to propose the proto-sex chromosome pair in P. nasus and P. pongoensis (Schemberger et al., 2011). Subsequently, the proto-sex chromosome of P. nasus and P. pongoensis was also described as containing (GATA)n repeats in the heterochromatic region of the short arm (Ziemniczak et al., 2014). Currently, it is known that the W heterochromatic region of Apareiodon sp. is predominantly composed of microsatellite expansion and DNA transposons, especially Helitron, Tc1-Mariner, and EnSpm (Schemberger et al., 2019; Oliveira et al., 2025). The in situ localization shows that chromosome pairs 6 and 8 are enriched with these repetitive elements in Parodon sp. and P. cf. buckleyi, respectively, supporting the proposition that they represent the proto-sex chromosomes. Besides that, the colocalization of pPh2004 sites with the Helitron DNA transposon suggest that this transposable element may mediate satellite dispersal, promoting karyotype diversification between Parodon sp. and P. cf. buckleyi.
In addition to cytogenetic data, the K2P genetic distances and molecular methods of species delimitation indicate that Parodon sp. and P. cf. buckleyi represent two distinct MOTUs within Parodontidae. Ward (2009) suggested a K2P value of 2% as a threshold for fish species identification, a criterion supported by Pereira et al. (2013) for various Neotropical fish. Within Parodontidae, interspecific K2P values above 2% have been reported (Bellafronte et al., 2013; Santos et al., 2018; Traldi et al., 2020). However, low genetic divergence based on COI has been reported between some paired species (Apareiodon piracicabae (Eigenmann, 1907) and A. vittatus Garavello, 1977, and between A. machrisi Travassos, 1957, and A. cavalcante Pavanelli & Britski, 2003), with K2P distances below 2% (Bellafronte et al., 2013; Traldi et al., 2020). Despite this reduced mitochondrial divergence, cytogenetic and morphological differences support the validity of these taxa as distinct species, suggesting that they may represent cases of recent or ongoing speciation (Bellafronte et al., 2013; Traldi et al., 2020). Haplotype network, K2P distance, and phylogenetic analyses suggest a close evolutionary relationship among Parodon sp., P. nasus, and P. cf. buckleyi. Cytogenetic comparisons provide additional support for the molecular findings and suggest recent divergence among these species. The quite similar karyotypic formula, the synteny of 18S and 5S rDNAs, and the presence of a pair of proto-sex chromosomes reaffirm their close relationship and distinguish these species karyotypically from other congeners.
The molecular divergence of COIsequence analyzed here comprises several well-supported Parodon lineages exhibiting high interspecific K2P distances (Tab. S2; Fig. 5). When the molecular phylogeny is compared with the geographic distribution of species across South American river basins (Fig. 1), a clear biogeographic pattern emerges, in which species occurring within the same sub-basin show closer phylogenetic relationships to one another. Most of the species in the previously described “P. suborbitalis complex” inhabit regions close to the Andean Cordillera (Londoño-Burbano et al., 2011). However, the presence of multiple well-supported branches representing Parodon species, together with the high observed genetic distances, indicates that the “P. suborbitalis complex” is not monophyletic.
The establishment of major Neotropical drainage systems, including the Amazon, Orinoco, and La Plata, occurred over the past 10 million years (Myr), highlighting the influence of Andean foreland dynamics and paleoarches on present-day watersheds through vicariance and headwater-capture events (Montoya-Burgos et al., 2003; Hubert, Renno, 2006). Within the proposed “P. suborbitalis complex”, only P. alfonsoi, P. magdalenensis, and P. suborbitalis exhibit a close phylogenetic relationship, all occurring in the northern portion of the trans-Andean region. However, these data should be interpreted with caution, as P. alfonsoi and P. magdalenensis share a haplotype and exhibit a K2P distance of 0%, suggesting recent diversification, incomplete lineage sorting, or potential hybridization, given that both species occur in the Magdalena River basin. Also inhabiting the northern trans-Andean region, P. apolinari and P. caliensis, originally not included in the “P. suborbitalis complex” due to morphological features (Pavanelli, 1999), show well-supported subclades close to P. suborbitalis in the molecular phylogeny, corroborating the non-monophyly of the species complex. These findings suggest that multiple Parodon lineages may have undergone diversification in the trans-Andean region. It is important to note that, of the nine species originally included in the complex (Londoño-Burbano et al., 2011), three of them, P. bifasciatus, P. carrikeri, and P. atratoensis, were not included in the present analyses due to the absence of available data, which limits a comprehensive assessment of the monophyly of the originally proposed complex.
All other Parodon species analyzed are cis-Andean and, similarly, have branches in the phylogenetic tree consistent with a biogeographic context. Parodon guyanensis occurs in the freshwater systems of the Guianas region, shows high genetic distance from congeners, and has a well-supported phylogenetic branch. Similarly, P. pongoensis and P. hilarii, which occur in the freshwater systems of the Araguaia-Tocantins and São Francisco, respectively, also have well-supported phylogenetic branches. Finally, Parodon sp., P. cf. buckleyi, and P. nasus, which are grouped in a single phylogenetic clade, inhabit the Amazon (first two) and La Plata (last one) basins. Marine regressions and changes in the Andean forelands between 10 and 8 Myr ago played a key role in the final establishment of the Amazon Basin (Hubert, Renno, 2006). In addition, around 10 Myr ago, the Paraná–Paraguay system (La Plata basin) separated from the proto-Amazon (Lundberg et al., 1998). Nevertheless, some evidence suggests that headwater-capture events connecting the Paraná system to the Amazon occurred within this timeframe (Räsänen et al., 1995; Lundberg et al., 1998), which could explain the dispersion and similarity among Parodon sp., P. cf. buckleyi, and P. nasus.
In this context, the Parodon species analyzed here are arranged in smaller groups presenting consistent genetic distances, which are closely associated within a biogeographic framework. These findings in Parodon are consistent with other phylogenetic studies of Neotropical freshwater fishes, which demonstrate that species diversification is closely associated with the geological history of South America’s hydrographic basins (Ribolli et al., 2021; Elías et al., 2023; Volpi et al., 2023).
It is important to emphasize that the molecular phylogeny presented here is based on the substitution patterns of a single mitochondrial gene. Although mitochondrial markers are informative and widely used in fishes (Morais-Silva et al., 2023; Souza et al., 2023; Almeida et al., 2024, among others), multilocus approaches incorporating independent nuclear loci provide a more robust reconstruction of species trees by better accounting for processes such as incomplete lineage sorting, species tree discordance, and historical gene flow (López-Fernández et al., 2010; Frable et al., 2016; de Queiroz et al., 2020; Ramirez et al., 2020). More recently, phylogenomic frameworks based on genome-wide data, such as ultraconserved elements (UCEs), RADseq, among others, have substantially improved phylogenetic resolution and provided greater power to infer diversification processes, timing of lineage splits, and patterns of speciation (Díaz-Arce et al., 2016; Ilves et al., 2018; Souza et al., 2022; Wang et al., 2026). Therefore, future studies integrating multilocus or phylogenomic datasets will be essential to refine evolutionary relationships and further test species boundaries within the Parodon. Despite the limitation of single-locus analyses, cytogenetic data indicate low divergence among Parodon sp., P. cf. buckleyi, and P. nasus, but clearly distinguish P. moreirai, P. pongoensis, and P. hilarii from both these species and from each other.
In conclusion, this study analyzed two Parodon species from the Amazon basin and identified several cytogenetic and molecular characteristics that differentiate them from other Parodon species. The findings support the hypothesis that Parodon sp. may constitute an undescribed species. Additionally, the presence of P. cf. buckleyi in the Madeira River basin, which is distant from its type locality in western Amazonia, Ecuador, may also reflect the existence of another undescribed species within the genus Parodon. Molecular analysis of specimens from the type locality of P. buckleyi would help clarify its taxonomic status. Furthermore, the data indicate that the “Parodon suborbitalis complex” comprises distinct lineages that are closely associated within a biogeographic framework.
Acknowledgments
References
Almeida RB, Azambuja MA, Nogaroto V, Oliveira C, Roxo FF, Zawadzki CH et al. DNA barcode shows discordant cases among morphological and molecular species identification in Isbrueckerichthys (Siluriformes: Loricariidae). Neotrop Ichthyol. 2024; 22(3):e240040. https://doi.org/10.1590/1982-0224-2024-0040
Antonelli A, Ariza M, Albert J, Andermann T, Azevedo J, Bacon C et al. Conceptual and empirical advances in Neotropical biodiversity research. PeerJ. 2018; 6:e5644. https://doi.org/10.7717/peerj.5644
Azambuja M, Marcondes DS, Nogaroto V, Moreira-Filho O, Vicari MR. Population structuration and chromosomal features homogeneity in Parodon nasus (Characiformes: Parodontidae): a comparison between lower and upper Paraná River representatives. Neotrop Ichthyol. 2022a; 20(1):e210162. https://doi.org/10.1590/1982-0224-2021-0162
Azambuja M, Nogaroto V, Moreira-Filho O, Vicari MR. U2 and U4 snDNA comparative chromosomal mapping in the neotropical fish genera Apareiodon and Parodon (Characiformes: Parodontidae). Zebrafish. 2023; 20(5):221–28. https://doi.org/10.1089/zeb.2023.0025
Azambuja M, Schemberger MO, Nogaroto V, Moreira-Filho O, Martins C, Vicari MR. Major and minor U small nuclear RNAs genes characterization in a neotropical fish genome: Chromosomal remodeling and repeat units dispersion in Parodontidae. Gene. 2022b; 826:146459. https://doi.org/10.1016/j.gene.2022.146459
Bandelt HJ, Forster P, Röhl A. Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol. 1999; 16(1):37–48. https://doi.org/10.1093/oxfordjournals.molbev.a026036
Bellafronte E, Margarido VP, Moreira-Filho O. Cytotaxonomy of Parodon nasus and Parodon tortuosus (Pisces, Characiformes): a case of synonymy confirmed by cytogenetic analyses. Genet Mol Biol. 2005; 28(4):710–16. https://doi.org/10.1590/S1415-47572005000500010
Bellafronte E, Schemberger MO, Artoni RF, Moreira-Filho O, Vicari MR. Sex chromosome system ZZ/ZW in Apareiodon hasemani Eigenmann, 1916 (Characiformes, Parodontidae) and a derived chromosomal region. Genet Mol Biol. 2012; 35(4):770–76. http://dx.doi.org/10.1590/S1415-47572012005000077
Bellafronte E, Mariguela TC, Pereira LHG, Oliveira C, Moreira-Filho O. DNA barcode of Parodontidae species from the La Plata river basin – applying new data to clarify taxonomic problems. Neotrop Ichthyol. 2013; 11(3):497–506. http://dx.doi.org/10.1590/S1679-62252013000300003
Bellafronte E, Schemberger MO, Moreira-Filho O, Almeida MC, Artoni RF, Margarido VP et al. Chromosomal markers in Parodontidae: an analysis of new and reviewed data with phylogenetic inferences. Rev Fish Biol Fish. 2011; 21(3):559–70. https://doi.org/10.1007/s11160-010-9177-3
Benson DA, Cavanaugh M, Clark K, Karsch-Mizrachi I, Lipman DJ, Ostell J et al. GenBank. Nucleic Acids Res. 2013; 41:36–42. https://doi.org/10.1093/nar/gks1195
Bertollo LAC, Cioffi MB, Moreira-Filho O. Direct chromosome preparation from freshwater teleost fishes. In: Ozouf-Costaz C, Pisano E, Foresti F, Almeida Toledo LF, editors. Fish cytogenetic techniques (Ray-fin fishes and Chondrichthyans). Boca Raton: CRC Press; 2015. p.21–26.
Blaxter ML. The promise of a DNA taxonomy. Philos Trans R Soc Lond B Biol Sci. 2004; 359(1444):669–79. https://doi.org/10.1098/rstb.2003.1447
Bouckaert R, Vaughan TG, Barido-Sottani J, Duchêne S, Fourment M, Gavryushkina A et al. Beast 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput Biol. 2019; 15(4):e1006650. https://doi.org/10.1371/journal.pcbi.1006650
Centofante L, Bertollo LAC, Moreira-Filho O. A ZZ/ZW sex chromosome system in a new species of the genus Parodon (Pisces, Parodontidae). Caryologia. 2002; 55(2):139–50. https://doi.org/10.1080/00087114.2002.10589270
Darriba D, Taboada GL, Doallo R, Posada D. jModelTest 2: more models, new heuristics and parallel computing. Nat Methods. 2012; 9:772. https://doi.org/10.1038/nmeth.2109
Dayrat B. Towards integrative taxonomy. Biol J Linn Soc. 2005; 85(3):407–17. https://doi.org/10.1111/j.1095-8312.2005.00503.x
Díaz-Arce N, Arrizabalaga H, Murua H, Irigoien X, Rodríguez-Ezpeleta N. RAD-seq derived genome-wide nuclear markers resolve the phylogeny of tunas. Mol Phylogenet Evol. 2016; 102:202–07. https://doi.org/10.1016/j.ympev.2016.06.002
Elías DJ, McMahan CD, Alda F, García-Alzate C, Hart PB, Chakrabarty P. Phylogenomics of trans-Andean tetras of the genus Hyphessobrycon Durbin 1908 (Stethaprioninae: Characidae) and colonization patterns of Middle America. PLoS ONE. 2023;18(1):e0279924. https://doi.org/10.1371/journal.pone.0279924
Fernandes MB, Bitencourt JÁ, Silva AT, Vicari MR, Azambuja M, Affonso PRAM. Small fishes, big issues: species delimitation in Hemigrammus marginatus Gill, 1958 (Acestrorhamphidae: Pristellinae) from Brazilian coastal basins based on integrative genetics. Zebrafish. 2025; 22(2):46–58. https://doi.org/10.1089/zeb.2024.0174
Floyd R, Abebe E, Papert A, Blaxter M. Molecular barcodes for soil nematode identification. Mol Ecol. 2002; 11(4):839–50. https://doi.org/10.1046/j.1365-294x.2002.01485.x
Frable BW, Melo BF, Sidlauskas BL, Hoekzema K, Vari RP, Oliveira C. Data on the multilocus molecular phylogenies of the Neotropical fish family Prochilodontidae (Teleostei: Characiformes). Data Brief. 2016; 9:128–42. https://doi.org/10.1016/j.dib.2016.08.015
Fricke R, Eschmeyer WN, Fong JD. Eschmeyer’s catalog of fishes: genera/ species by family/subfamily [Internet]. San Francisco: California Academy of Science; 2025. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp
Fujisawa T, Barraclough TG. Delimiting species using single-locus data and the generalized mixed yule coalescent approach: a revised method and evaluation on simulated data sets. Syst Biol. 2013; 62(5):707–24. https://doi.org/10.1093/sysbio/syt033
Gatto KP, Mattos JV, Seger KR, Lourenço LB. Sex chromosome differentiation in the frog genus Pseudis involves satellite DNA and Chromosome rearrangements. Front Genet. 2018; 9:301. https://doi.org/10.3389/fgene.2018.00301
Gavazzoni M, Brezinski FC, Pedroso TH, Pavanelli CS, Graça WJ, Blanco DR et al. Integrative taxonomy suggests resurrection of species of the Astyanax bimaculatus group (Characiformes, Characidae). Zebrafish. 2024; 21(5):349–59. https://doi.org/10.1089/zeb.2024.0132
Hatanaka T, Galetti Jr. PM. Mapping of the 18S and 5S ribosomal RNA genes in the fish Prochilodus argenteus Agassiz, 1829 (Characiformes, Prochilodontidae). Genetica. 2004; 122:239–44. https://doi.org/10.1007/s10709-004-2039-y
Hebert PDN, Cywinska A, Ball SL, deWaard JR. Biological identifications through DNA barcodes. Proc R Soc Lond B Biol Sci. 2003; 270(1512):313–21. https://doi.org/10.1098/rspb.2002.2218
Hubert N, Renno J-F. Historical biogeography of South American freshwater fishes. J Biogeogr. 2006; 33(8):1414–36. https://doi.org/10.1111/j.1365-2699.2006.01518.x
Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics. 2001; 17(8):754–55. https://doi.org/10.1093/bioinformatics/17.8.754
Ilves KL, Torti D, López-Fernández H. Exon-based phylogenomics strengthens the phylogeny of Neotropical cichlids and identifies remaining conflicting clades (Cichliformes: Cichlidae: Cichlinae). Mol Phylogenet Evol. 2018; 118:232–43. https://doi.org/10.1016/j.ympev.2017.10.008
Jesus CM, Moreira-Filho O. Karyotypes of three species of Parodon (Teleostei: Parodontidae). Ichthyol Explor Freshw. 2000; 11(1):75–80.
Jézéquel C, Tedesco PA, Bigorne R, Maldonaldo-Ocampo JA, Ortega H, Hidalgo M et al. A database of freshwater fish species of the Amazon basin. Sci Data. 2020; 7:96. https://doi.org/10.1038/s41597-020-0436-4
Kapli P, Lutteropp S, Zhang J, Kobert K, Pavlidis P, Stamatakis A et al. Multi-rate poisson tree processes for single-locus species delimitation under maximum likelihood and Markov chain Monte Carlo. Bioinformatics. 2017; 33(11):1630–38. https://doi.org/10.1093/bioinformatics/btx025
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S et al. Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012; 28(12):1647–49. https://doi.org/10.1093/bioinformatics/bts199
Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol Biol Evol. 2018; 35(6):1547–49. https://doi.org/10.1093/molbev/msy096
Levan A, Fredga K, Sandberg AA. Nomenclature for centromeric position on chromosomes. Hereditas. 1964; 52(2):201–20. https://doi.org/10.1111/j.1601-5223.1964.tb01953.x
Leigh JW, Bryant D. POPART: full-feature software for haplotype network construction. Methods Ecol Evol. 2015; 6(9):1110–16. https://doi.org/10.1111/2041-210X.12410
Librado P, Rozas J. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics. 2009; 25(11):1451–52. https://doi.org/10.1093/bioinformatics/btp187
Limeira Filho D, França ERR, Costa DKP, Lima RC, Nascimento MHS, Batista JS et al. Molecular evidence reveals taxonomic uncertainties and cryptic diversity in the Neotropical catfish of the genus Pimelodus (Siluriformes: Pimelodidae). Biology. 2024; 13(3):162. https://doi.org/10.3390/biology13030162
Londoño-Burbano A, Román-Valencia C, Taphorn DC. Taxonomic review of Colombian Parodon (Characiformes: Parodontidae), with descriptions of three new species. Neotrop Ichthyol. 2011; 9(4):709–30. https://doi.org/10.1590/S1679-62252011000400003
López-Fernández H, Winemiller KO, Honeycutt RL. Multilocus phylogeny and rapid radiations in Neotropical cichlid fishes (Perciformes: Cichlidae: Cichlinae). Mol Phylogenet Evol. 2010; 55(3):1070–86. http://dx.doi.org/10.1016/j.ympev.2010.02.020
Lundberg JG, Marshall LG, Guerrero J, Horton B, Malabarba MCSL, Wesselingh F. The stage for Neotropical fish diversification: a history of tropical South American rivers. In: Malabarba, LR, Reis RE, Vari RP, Lucena ZM, Lucena CAS, editors. Phylogeny and classification of Neotropical fishes Porto Alegre: Edipucrs; 1998. p.13–48.
Luo A, Ling C, Ho SYW, Zhu C. Comparison of methods for molecular species delimitation across a range of speciation scenarios. Syst Biol. 2018; 67(5):830–46. https://doi.org/10.1093/sysbio/syy011
Martins C, Galetti Jr. PM. Chromosomal localization of 5S rDNA genes in Leporinus Fish (Anostomidae, Characiformes). Chromosome Res. 1999; 7:363–67. https://doi.org/10.1023/A:1009216030316
Milliman JD, Farnsworth KL. River discharge to the coastal ocean: a global synthesis. Cambridge: Cambridge University Press; 2011.
Montoya-Burgos JI. Historical biogeography of the catfish genus Hypostomus (Siluriformes: Loricariidae), with implications on the diversification of Neotropical ichthyofauna. Mol Ecol. 2003. 12(7):1855–67. https://doi.org/10.1046/j.1365-294x.2003.01857.x
Moraes RLR, Sassi FMC, Vidal JAD, Goes CAG, Santos RZ, Stornioli JHF et al. Chromosomal rearrangements and satellite DNAs: extensive chromosomal reshuffling and the evolution of neo-sex chromosomes in the genus Pyrrhulina (Teleostei; Characiformes). Int J Mol Sci. 2023; 24(17):13654. https://doi.org/10.3390/ijms241713654
Morais-Silva JP, Scorsim B, Gonçalves G, Frota A, Graça WJ, Oliveira AV. Molecular marks reveal a new and possibly threatened species of Cnesterodon (Poeciliidae, Cnesterodontini) from the upper Paraná River basin, Brazil. Zebrafish. 2023; 20(1):37–45. https://doi.org/10.1089/zeb.2022.0052
Moreira-Filho O, Bertollo LAC, Galetti Jr. PM. Structure and variability of nucleolar organizer regions in Parodontidae fish. Can J Genet Cytol. 1984; 26(5):564–68. https://doi.org/10.1139/g84-089
Moreira-Filho O, Bertollo LAC, Galetti Jr. PM. Karyotypic study of some species of family Parodontidae (Pisce-Cypriniformes). Caryologia. 1985; 38(1):47–55. https://doi.org/10.1080/00087114.1985.10797729
Moreira-Filho O, Bertollo LAC, Galetti Jr. PM. Distribution of sex chromosome mechanism in neotropical fish and description of a ZZ/ZW system in Parodon hilarii (Parodontidae). Caryologia. 1993; 46:115–25. https://doi.org/10.1080/00087114.1993.10797253
Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980; 8(19):4321–26. https://doi.org/10.1093/nar/8.19.4321
Nascimento VD, Coelho KA, Nogaroto V, Almeida RB, Ziemniczak K, Centofante L et al. Do multiple karyomorphs and population genetics of freshwater darter characine (Apareiodon affinis) indicate chromosomal speciation? Zool Anz. 2018; 272:93–103. https://doi.org/10.1016/j.jcz.2017.12.006
Nirchio M, Masache MC, Paim FG, Cioffi MB, Moreira-Filho O, Barriga R et al. Chromosome analysis in Saccodon wagneri (Characiformes) and insights into the karyotype evolution of Parodontidae. Neotrop Ichthyol. 2021; 19(1):e200103. https://doi.org/10.1590/1982-0224-2020-0103
Oliveira FS, Azambuja M, Schemberger MO, Nascimento VD, Oliveira JIN, Wolf IR et al. Characterization of hAT DNA transposon superfamily in the genome of Neotropical fish Apareiodon sp. Mol Genet Genomics. 2024; 299:96. https://doi.org/10.1007/s00438-024-02190-x
Oliveira FS, Brann T, Wolf IR, Nogaroto V, Martins C, Protasio AV et al. The landscape of transposable elements distribution in the genome of Neotropical fish Apareiodon sp. (Characiformes: Parodontidae). Chromosome Res. 2025; 33:6. https://doi.org/10.1007/s10577-025-09765-3
Paço A, Chaves R, Vieira-da-Silva A, Adega F. The involvement of repetitive sequences in the remodelling of karyotypes: The Phodopus genomes (Rodentia, Cricetidae). Micron. 2013; 46:27–34. https://doi.org/10.1016/j.micron.2012.11.010
Pavanelli CS. Revisão taxonômica da família Parodontidae (Ostariophysi: Characiformes). [PhD Thesis]. São Carlos: Universidade Federal de São Carlos; 1999. Avaliable from: ftp://ftp.nupelia.uem.br/users/Carla/Parodontidae.pdf
Pavanelli CS. Family Parodontidae. In: Reis RE, Kullander SO, Ferraris Jr. CJ, editors. Check list of the freshwater fishes of South and Central America. Porto Alegre: Edipucrs; 2003. p.46–50.
Pavanelli CS, Britski HA. Apareiodon Eigenmann, 1916 (Teleostei, Characiformes), from the Tocantins-Araguaia basin, with description of three new species. Copeia. 2003; 2:337–48. https://doi.org/10.1643/0045-8511(2003)003[0337:AETCFT]2.0.CO;2
Pereira LHG, Hanner R, Foresti F, Oliveira C. Can DNA barcoding discriminate megadiverse Neotropical freshwater fish fauna? BMC Genet. 2013; 14:20. https://doi.org/10.1186/1471-2156-14-20
Pinkel D, Straume T, Gray JW. Cytogenetic analysis using quantitative, high-sensitivity, fluorescence hybridization. Proc Natl Acad Sci USA. 1986; 83(9):2934–38. https://doi.org/10.1073/pnas.83.9.2934
Pons J, Barraclough TG, Gomez-Zurita J, Cardoso A, Duran DP, Hazell S et al. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst Biol. 2006; 55(4):595–609. https://doi.org/10.1080/10635150600852011
Puillandre N, Brouillet S, Achaz G. ASAP: assemble species by automatic partitioning. Mol Ecol Resour. 2021; 21(2):609–20. https://doi.org/10.1111/1755-0998.13281
Puillandre N, Lambert A, Brouillet S, Achaz G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol Ecol. 2012; 21(8):1864–77. https://doi.org/10.1111/j.1365-294X.2011.05239.x
de Queiroz LJ, Cardoso Y, Jacot-des-Combes C, Bahechar IA, Lucena CA, Rapp Py-Daniel L et al. Evolutionary units delimitation and continental multilocus phylogeny of the hyperdiverse catfish genus Hypostomus. Mol Phylogenet Evol. 2020; 145:106711. https://doi.org/10.1016/j.ympev.2019.106711
R Development Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2013. Available from: https://www.r-project.org/
Rambaut A, Suchard MA, Xie D, Drummond AJ. Tracer v1.6. 2014. Available from: http://beast.bio.ed.ac.uk/Tracer
Ramirez JL, Birindelli JLO, Galetti Jr PM. A new genus of Anostomidae (Ostariophysi: Characiformes): Diversity, phylogeny and biogeography on cytogenetic, molecular and morphological data. Mol Phylogenet Evol. 2017; 107:308–23. https://doi.org/10.1016/j.ympev.2016.11.012
Ramirez JL, Santos CA, Machado CB, Oliveira AK, Garavello JC, Britski HA et al. Molecular phylogeny and species delimitation of the genus Schizodon (Characiformes, Anostomidae). Mol Phylogenet Evol. 2020; 153:106959. https://doi.org/10.1016/j.ympev.2020.106959
Räsänen ME, Linna AM, Santos JC, Negri FR. Late miocene tidal deposits in the amazonian foreland basin. Science. 1995; 269(5222):386–90. https://doi.org/10.1126/science.269.5222.386
Ratnasingham S, Hebert PDN. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol Ecol Notes. 2007; 7(3):355–64. https://doi.org/10.1111/j.1471-8286.2007.01678.x
Ribas MS, Azambuja M, Nogaroto V, Vicari MR. Emergence of satellite DNAs suggests centromeric repositioning as a driver of karyotypic variation of the freshwater darter characines (Apareiodon affinis). Chromosome Res. 2026; 34(1):2. https://doi.org/10.1007/s10577-026-09793-7
Ribolli J, Zaniboni Filho E, Scaranto BMS, Shibatta OA, Machado CB. Cryptic diversity and diversification processes in three cis-Andean Rhamdia species (Siluriformes: Heptapteridae) revealed by DNA barcoding. Genet Mol Biol. 2021; 44(3):e20200470. https://doi.org/10.1590/1678-4685-GMB-2020-0470
Santos EO, Deon GA, Almeida RB, Oliveira EA, Nogaroto V, Silva HP et al. Cytogenetics and DNA barcode reveal an undescribed Apareiodon species (Characiformes: Parodontidae). Genet Mol Biol. 2019; 42(2):365–73. http://dx.doi.org/10.1590/1678-4685-gmb-2018-0066
Schemberger MO, Bellafronte E, Nogaroto V, Almeida MC, Schühli GS, Artoni RF et al. Differentiation of repetitive DNA sites and sex chromosome systems reveal closely related group in Parodontidae (Actinopterygii: Characiformes). Genetica. 2011; 139:1499–508. https://doi.org/10.1007/s10709-012-9649-6
Schemberger MO, Nascimento VD, Coan R, Ramos É, Nogaroto V et al. DNA transposon invasion and microsatellite accumulation guide W chromosome differentiation in a Neotropical fish genome. Chromosoma. 2019; 128:547–60. https://doi.org/10.1007/s00412-019-00721-9
Serrano ÉA, Melo BF, Freitas-Souza D, Oliveira MLM, Utsunomia R et al. Species delimitation in Neotropical fishes of the genus Characidium (Teleostei, Characiformes). Zool Scr. 2018; 48(1):69–80. https://doi.org/10.1111/zsc.12318
Souza TB, Ferreira DC, Silva HP, Netto-Ferreira AL, Venere PC. DNA Barcoding of Pyrrhulina australis (Characiformes: Lebiasinidae) reveals unexpected cryptic diversity in the group. Neotrop Ichthyol. 2023; 21(4):e230037. https://doi.org/10.1590/1982-0224-2023-0037
Souza CS, Melo BF, Mattox GM, Oliveira C. Phylogenomic analysis of the Neotropical fish subfamily Characinae using ultraconserved elements (Teleostei: Characidae). Mol Phylogenet Evol. 2022; 171:107462. https://doi.org/10.1016/j.ympev.2022.107462
Sumner AT. A simple technique for demonstrating centromeric heterochromatin. Exp Cell Res. 1972; 75(1):304–06. https://doi.org/10.1016/0014-4827(72)90558-7
Tang CQ, Humphreys AM, Fontaneto D, Barraclough TG. Effects of phylogenetic reconstruction method on the robustness of species delimitation using single-locus data. Methods Ecol Evol. 2014; 5(10):1086–94. https://doi.org/10.1111/2041-210X.12246
Tedesco PA, Beauchard O, Bigorne R, Blanchet S, Buisson L, Conti L et al. A global database on freshwater fish species occurrence in drainage basin. Sci Data. 2017; 4:170141. https://doi.org/10.1038/sdata.2017.141
Terán GE, Benitez MF, Marcos Mirande. Opening the Trojan horse: phylogeny of Astyanax, two new genera and resurrection of Psalidodon (Teleostei: Characidae). Zool J Linn Soc. 2020; 190(4):1217–34. https://doi.org/10.1093/zoolinnean/zlaa019
Tisseuil C, Cornu J-F, Beauchard O, Brosse S, Darwall W, Holland R et al. Global diversity patterns and cross-taxa convergence in freshwater systems. J Anim Ecol. 2013; 82:365–76. https://doi.org/10.1111/1365-2656.12018
Traldi JB, Blanco DR, Vicari MR, Martinez JF, Lui RL, Artoni RF et al. Physical mapping of (GATA)n and (TTAGGG)n sequences in species of Hypostomus (Siluriformes, Loricariidae). J Genet. 2013; 92:127–30. https://doi.org/10.1007/s12041-013-0224-4
Traldi JB, Vicari MR, Martinez JF, Blanco DR, Lui RL, Azambuja M et al. Recent Apareiodon species evolutionary divergence (Characiformes: Parodontidae) evidenced by chromosomal and molecular inference. Zoo Anz. 2020; 289:166–76. https://doi.org/10.1016/j.jcz.2020.10.010
Traldi JB, Vicari MR, Martinez JF, Blanco DR, Lui RL, Moreira-Filho O. Chromosome analyses of Apareiodon argenteus and Apareiodon davisi (Characiformes, Parodontidae): An extensive chromosomal polymorphism of 45S and 5S ribosomal DNAs. Zebrafish. 2016; 13(1):19–25. https://doi.org/10.1089/zeb.2015.1124
Vicente VE, Bertollo LAC, Valentini SR, Moreira-Filho O. Origin and differentiation of a sex chromosome system in Parodon hilarii (Pisces, Parodontidae). Satellite DNA, G- and C-banding. Genetica. 2003; 119:115–20. https://doi.org/10.1023/A:1026082904672
Vicente VE, Jesus CM, Moreira-Filho O. Chromosomal localization of 5S and 18S rRNA genes in three Parodon species (Pisces, Parodontidae). Caryologia. 2001; 54(4):365–69. https://doi.org/10.1080/00087114.2001.10589247
Volpi TA, Monjardim M, Sarmento-Soares LM, Fagundes V. Pleistocene aquatic refuges support the East-West separation of the Neotropical catfish Trichomycterinae (Siluriformes: Trichomycteridae) and high diversity in the Magdalena, Guiana, and Paraná-Paraguay basins. Diversity. 2023; 15(8):929. https://doi.org/10.3390/d15080929
Wang A, Stiassny ML, Melo BF. Phylogenomics, biogeography, and description of a new subfamily and genus of African characiform fishes (Teleostei: Alestidae). Mol Phylogenet Evol. 2026; 2027:108546. https://doi.org/10.1016/j.ympev.2026.108546
Ward RD. DNA barcode divergence among species and genera of birds and fishes. Mol Ecol Res. 2009; 9(4):1077–85. https://doi.org/10.1111/j.1755-0998.2009.02541.x
Ward RD, Zemlak TS, Innes BH, Last PR, Hebert PDN. DNA barcoding Australia’s fish species. Philos Trans R Soc Lond B Biol Sci. 2005; 360:1847–57. https://doi.org/10.1098/rstb.2005.1716
Wolf IR, Schemberger MO, Azambuja M, Oliveira FS, Nogaroto V, Valente GT et al. The long-read assembly of Apareiodon sp., a neotropical fish with ZZ/ZW sex chromosome system. Genet Mol Biol. 2024; 47(4):e20240098. https://doi.org/10.1590/1678-4685-GMB-2024-0098
Zhang J, Kapli P, Pavlidis P, Stamatakis A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics. 2013; 29(22):2869–76. https://doi.org/10.1093/bioinformatics/btt499
Ziemniczak K, Traldi JB, Nogaroto V, Almeida MC, Artoni RF, Moreira-Filho O et al. In situ localization of (GATA)n and (TTAGGG)n repeated DNAs and W sex chromosome differentiation in Parodontidae (Actinopterygii: Characiformes). Cytogenet Genome Res. 2014; 144(4):325–32. https://doi.org/10.1159/000370297
Authors

![]() Matheus Azambuja1,
Matheus Azambuja1, ![]() Ezequiel A. de Oliveira2,
Ezequiel A. de Oliveira2, ![]() Francisco de M. C. Sassi2,3,
Francisco de M. C. Sassi2,3, ![]() Viviane Nogaroto4,
Viviane Nogaroto4, ![]() Orlando Moreira-Filho2,
Orlando Moreira-Filho2, ![]() Carla S. Pavanelli5 and
Carla S. Pavanelli5 and ![]() Marcelo R. Vicari1,4
Marcelo R. Vicari1,4 ![]()
[1] Programa de Pós-Graduação em Genética, Universidade Federal do Paraná, Avenida Coronel Francisco H. dos Santos, 100, Jardim das Américas, 81531-990, Curitiba, PR, Brazil. (MA) matheus_azambuja@hotmail.com, (MRV) vicarimr@uepg.br (corresponding author).
[2] Programa de Pós-Graduação em Genética, Universidade Federal do Paraná, Avenida Coronel Francisco H. dos Santos, 100, Jardim das Américas, 81531-990, Curitiba, PR, Brazil. (MA) matheus_azambuja@hotmail.com, (MRV) vicarimr@uepg.br (corresponding author).
[3] School of Life Sciences, Southwest University, Tiansheng Road, Beibei District, Chongqing 400715, P. R. China.
[4] Departamento de Biologia Estrutural, Molecular e Genética, Universidade Estadual de Ponta Grossa, Avenida Carlos Cavalcanti, 4748, 84030-900, Ponta Grossa, PR, Brazil. (VN) vivianenogaroto@hotmail.com.
[5] Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura, Universidade Estadual de Maringá, Avenida Colombo, 5790, 87020-900, Maringá, PR, Brazil. (CSP) carlasp@nupelia.uem.br.
Authors’ Contribution 

Matheus Azambuja: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing-original draft, Writing-review and editing.
Ezequiel A. de Oliveira: Formal analysis, Investigation, Writing-original draft.
Francisco de M. C. Sassi: Formal analysis, Investigation, Writing-original draft.
Viviane Nogaroto: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Visualization, Writing-original draft, Writing-review and editing.
Orlando Moreira-Filho: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Project administration, Visualization, Writing-original draft, Writing-review and editing.
Carla S. Pavanelli: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Visualization, Writing-original draft, Writing-review and editing.
Marcelo R. Vicari: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Ethical Statement
Fish were collected with the authorization of the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBIO), Sistema de Autorização e Informação em Biodiversidade (SISBIO, license numbers 10538 and 15117), and Sistema Nacional de Gestão do Patrimônio Genético e do Conhecimento Tradicional Associado (SISGEN Nº AE12D3D). The procedures were approved by the Comição de Ética no Uso de Animal of the Universidade Estadual de Ponta Grossa (Process CEUA Nº 6/2019) and Biosafety Certification according to Comissão Técnica Nacional de Biossegurança – CTNBio (CQB Nº 0063/98). The procedures of this study are in agreement with the Ethics Committee of Animal Usage of the Universidade Estadual de Ponta Grossa, Brazil (Protocol: 06/2019).
Competing Interests
The author declares no competing interests.
Data availability statement
All the data supporting the findings are included in this published article, its supplementary information files.
AI statement
Language editing was assisted by Grammarly, without affecting the scientific content.
Funding
This study was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) to MRV #313566/2023–2, and CSP #307124/2023–1), Fundação Araucária (Fundação Araucária de Apoio ao Desenvolvimento Científico e Tecnológico do Estado do Paraná, MRV: 9/2017), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) to MA – Finance Code 001.
Supplementary Material
Supplementary material SUP
How to cite this article
Azambuja M, Oliveira EA, Sassi FMC, Nogaroto V, Moreira-Filho O, Pavanelli CS, Vicari MR. Integrative cytogenetic and molecular evidence reveals a new cryptic lineage within the “Parodon suborbitalis complex” (Characiformes: Parodontidae). Neotrop Ichthyol. 2026; 24(2):e250119. https://doi.org/10.1590/1982-0224-2025-0119
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2025 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted February 6, 2026
Accepted February 6, 2026
![]() Submitted July 7, 2025
Submitted July 7, 2025
![]() Epub June 19, 2026
Epub June 19, 2026