![]() Juliana Paulo da Silva-Novelli1,2,
Juliana Paulo da Silva-Novelli1,2, ![]() Heron Oliveira Hilário3,
Heron Oliveira Hilário3, ![]() Daniel Cardoso Carvalho3,
Daniel Cardoso Carvalho3, ![]() Ronaldo Fernando Martins-Pinheiro4 and
Ronaldo Fernando Martins-Pinheiro4 and ![]() Luisa Maria Sarmento-Soares1,4,5
Luisa Maria Sarmento-Soares1,4,5 ![]()
PDF: Download Here | Supplementary: Sup | Cite this article
Associate Editor: ![]() Marcelo Ciof
Marcelo Ciof
Section Editor: ![]() Izeni Farias
Izeni Farias
Editor-in-chief: ![]() José Birindelli
José Birindelli
Abstract
Trichogenes corresponde a um táxon basal na filogenia da família, com três espécies de distribuição endêmica aos riachos da Mata Atlântica. Uma delas, Trichogenes claviger, ocorre em riachos florestados de primeira e segunda ordem, nas cabeceiras do rio Itapemirim, ao sul do Espírito Santo. Até o momento, a espécie é conhecida de apenas duas localidades na sub-bacia do córrego Picada Comprida, um riacho formador do rio Caxixe. O presente estudo usou o DNA ambiental (eDNA metabarcoding) para detecção desta espécie de bagrinho ameaçada de extinção. Foram coletados indivíduos empregando técnicas tradicionais de amostragem e a partir destas amostras foram geradas referências barcode para a espécie, o que permitiu sua detecção através da amostragem por eDNA em três das dez localidades investigadas. Adicionalmente à espécie alvo deste estudo, foi também possível detectar outras espécies de peixes nas amostras de água. Uma comparação dos métodos de estimativa da composição das espécies por riacho amostrado indicou que a metodologia de eDNA metabarcoding é capaz de identificar com maior precisão a diversidade de espécies de peixes em um dado riacho em comparação com a metodologia tradicional com redes de pesca. Os resultados alcançados trazem um melhor entendimento da dinâmica de distribuição espacial de Trichogenes claviger podendo confirmar seu status de conservação. O poder de detecção do método de eDNA metabarcoding se revelou uma ferramenta para monitoramento de espécies raras ou ameaçadas.
Palavras-chave: Conservação,Espécie ameaçada, Espírito Santo, Floresta Atlântica, Riachos.
Introduction
Trichomycteridae is one of the most diverse catfish families in South America, with more than 360 valid species (Ferraris, 2007; Rizzato, Bichuette, 2014; DoNascimiento, 2015; Fricke et al., 2024). Within this family lies the subfamily Trichogeninae, comprising a single genus, Trichogenes, whose species attracted great interest from the scientific community. Recognized as a relictual group within the family (e.g., Grandcolas, Trewick, 2016), the sister-group position of the taxon (sensu Krell, Cranston, 2004) aggregates plesiomorphic features that are critical for polarizing characters in phylogenetic studies (Wiley, Lieberman, 2011; following the principles of Hennig, 1966).
The Trichogenes genus comprises three valid species endemic to the Atlantic Forest biome, being all the known species restricted to southwest Brazil (Fricke et al., 2024). Trichogenes longipinnis Britski & Ortega, 1983, which is found in the headwaters of the rio Parati-Mirim and coastal rivers of southern Rio de Janeiro and northern São Paulo; T. beagle dePinna, Reis & Britski, 2020,from the upper rio Doce in Minas Gerais and T. claviger dePinna, Helmer, Britski & Nunes, 2010from the rio Itapemirim headwaters in southern Espírito Santo.
The distribution of Trichogenes claviger is restricted to a single micro-basin. The species is known only from two proximate sites along the Picada Comprida stream, a headwater tributary of the Caxixe River, located in the municipality of Castelo, within the state of Espírito Santo in southeastern Brazil. These small order rivers correspond to a well-preserved headwater environment. The fishes are small sized (largest known specimen reaching only 5.1 cm), being active swimmers in mid-water during the day and most of the time the only species of fish present in that stretch of the stream (Sarmento-Soares et al., 2018). However, after ten years since its description, T. claviger remains poorly known geographically. Targeted field expeditions conducted between 2018 and 2023 in headwater streams of the Itapemirim and Jucu basins, including areas reported in Sarmento-Soares et al. (2018), failed to record new populations prior to the present study (field expeditions led by LMSS, RFMP, unpublished data).
Indeed, the restricted geographical area of occupation leads the species to be categorized as Critically Endangered (CR) in red lists (MMA, 2014; IUCN, 2022). Trichogenes claviger was ranked as a target species in the Brazilian National Plan for the Conservation of Endangered Atlantic Forest Fish and Aeglid crustaceans (PAN Peixes e Eglas da Mata Atlântica) (MMA/ICMBio, 2019), ended in December 2024.
These critically endangered populations face a high risk of extinction in the wild. Detecting their presence and understanding their distribution is particularly challenging with traditional, invasive methods such as gillnets and dip nets (Bonfil et al., 2021), as these techniques can have a direct impact on the already vulnerable sampled community. EnvironmentalDNA (eDNA) emerged as an important tool for assessing biodiversity and detecting rare and endangered species in their natural environment (Goldberg et al., 2015; Thomsen, Willerslev, 2015; Cristescu, Hebert, 2018). The eDNA metabarcoding technique, originally developed in the late 1980s for the characterization of microbial communities from DNA recovered from soil samples, evolved into a powerful tool that enables detection of many organisms with great sensitivity. As this non-invasive approach allows the detection of multiple species through the DNA released into the environment, it is a particularly effective technique for finding traces of rare organisms (Thomsen, Willerslev, 2015; Boivin-Delisle et al., 2020; De la Ossa-Guerra et al., 2025). The data generated by this method are increasingly being used to direct and prioritize conservation efforts, from monitoring protected species (Biggs et al., 2015) to informing environmental management policies (Lodge et al., 2012; Goldberg et al., 2016).
For eDNA metabarcoding to be able to detect a given species it is necessary that a reference sequence of a marker region is available for it, which is yet rarely the case for rich communities, especially at the Neotropics (Carvalho, Leal, 2023). Up to date, for the Trichogenes species, only T. longipinnis has partial reference sequences available for the mitochondrial regions D-loop, COI and Cytochrome B, more often used for barcoding applications, and has no references for the 12S rRNA region, targeted by the mini barcode marker MiFish (Takahashi et al., 2023). Since this marker is the most widely used for fish eDNA metabarcoding, none of the Trichogenes species can be detected on conventional eDNA studies, they are yet invisible though the eDNA lenses.
A targeted multi-species approach was adopted for analyzing environmental DNA samples by eDNA metabarcoding using a generalist molecular marker for fish. Toward this end we validate the use of eDNA metabarcoding in the detection of T. claviger in streams close to points where the species had been previously reported, expanding records of the species’ geographic distribution.
In this study we have generated new reference sequences from three T. claviger individuals for the 12S rRNA marker region, enabling its detection with eDNA metabarcoding. We also evaluate the eDNA metabarcoding and visual survey methodologies complementarity, and applied both techniques to expand the known area of occurrence for the species, as a model for endangered species detection in remote areas.
Material and methods
Study area. The study area is located between the municipalities of Castelo and Vargem Alta in mountainous areas of southern Espírito Santo. With approximately 85 km2 of surface area and 45 km of perimeter, the area investigated included the headwaters of two river basins, the Itapemirim (formed by the Caxixe stream; Fig. 1A) and Jucu (formed by the rio Fruteiras; Fig. 1B).The elevation within the study area varies between 1,150 and 1,300 m. Two sampling expeditions were carried out: the first in September 2021 focused on specimen collection, and the second in March 2023 aimed at species detection using environmental DNA (eDNA) alongside traditional methods (namely, dip nets for targeted capture and gillnets for passive sampling). In the September expedition, for the investigation of the species T. claviger, it was necessary to collect some specimens to create a PCR and sequencing protocol using tissue fragments from these animals, so that, in the eDNA analyses, it would be possible to compare them with sequences available in the databases for species confirmation.
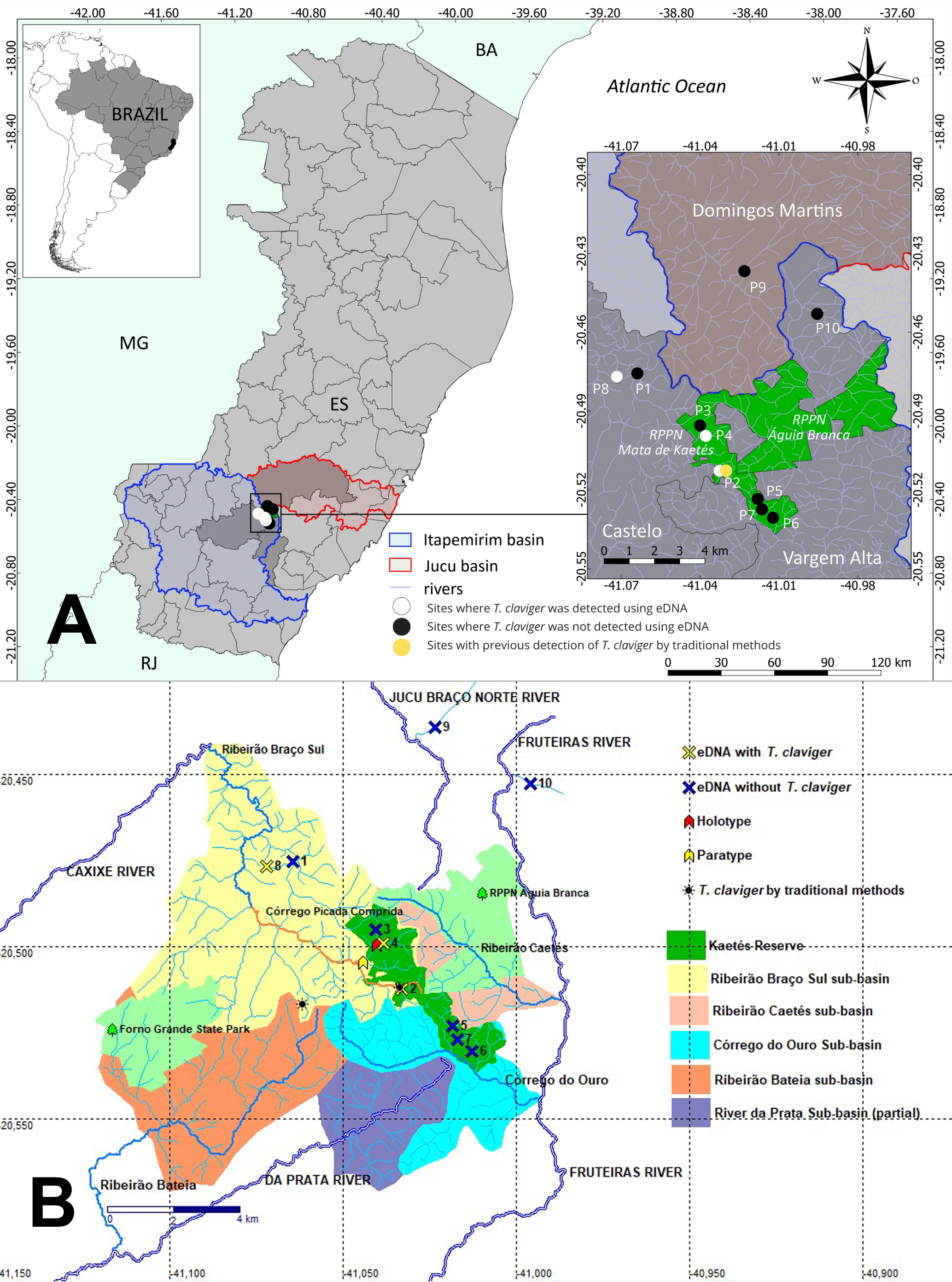
FIGURE 1| A. Rio Itapemirim (blue contour) and rio Jucu (red contour) in southern Espírito Santo. The black circles indicate eDNA sampling points; white circles indicate detection of Trichogenes claviger. Areas highlighted in green, RPPN Mata de Kaetés and RPPN Águia Branca. B. Detail of detection samples on headwaters of Ribeirão braço Sul, the only river with records of T. claviger (in yellow contour). Detected presence (yellow X) in collecting localities 2, 4 (type locality) and 8. Absence (blue X) tentative in collecting localities 1, 3, 5, 6, 7, 9 and 10. Green contour corresponds to protected areas- Parque Estadual do Forno Grande, Reserva Particular do Patrimônio Natural, RPPN Águia Branca (in light green) and Reserva Particular do Patrimônio Natural, RPPN Mata de Kaetés (in dark green).
Ten georeferenced sites were sampled in the mountainous headwaters of the Itapemirim River and Jucu River basins, in streams located between Forno Grande State Park and the Private Natural Heritage Reserve (RPPN) Águia Branca (Figs. 1A, B). In the Itapemirim River basin, the sampled locations include three localities at ribeirão Braço Sul sub-basin: Point P2 at the Picada Comprida stream; Point P3 on a right-bank tributary of Picada Comprida stream; Points P1 and P8 on a right-bank tributary of ribeirão Braço Sul. In the ribeirão Caetés sub-basin: Points P4 and P5 on right-bank tributaries of ribeirão Caetés. In the córrego do Ouro sub-basin: Points P6 and P7 on left-bank tributaries of córrego do Ouro. In the rio Fruteiras sub-basin: Point P10 on a left-bank tributary of rio Fruteiras. Additionally, in the Jucu River basin, a single location was sampled: Point P9 on a right-bank tributary of the rio Jucu Braço Norte (Fig. 1).
Specimens collection for reference sequence generation. For field sampling, sieves and seines were used. At each point, a combination of fishing resources was used to ensure sampling without disturbing the local environment. The locality was sampled, whenever possible, by going through a stretch of approximately 50 m upstream. The specimens were euthanized with menthol, fixed in eugenol and transported to the laboratory, and cataloged. Some specimens collected were photographed in a field aquarium. Institutional abbreviations follow Sabaj (2025), with the Instituto Nacional da Mata Atlântica [formerly Museu de Biologia Professor Mello Leitão], Santa Teresa (MBML) as the institution of deposit of fishes sampled. Additionally, the genetic samples were processed at the Laboratório de Genética da Conservação (LGC) at Pontifícia Universidade Católica of Minas Gerais.
Generation of Trichogenes claviger 12S rRNA reference sequences. Five specimens of T. claviger were collected from the Braço Sul stream (sampling point P8) and RPPN Mata de Kaetés (P2), tributary of the rio Caxixe, Itapemirim basin (Fig. 2), for genetic reference construction. Tissue samples were obtained and deposited in the fish tissue collection of the Conservation Genetics Laboratory of PUC Minas. Genomic DNA was obtained by salt extraction adapted from Aljanabi, Martinez (1997). The 12S rRNA region was amplified using the primers MiFish-U-F 5’-GTCGGTAAAACTCGTGCCAGC-3’ (Miya et al.,2015) and the Teleo REV ACTTCCGGTACACTTACCATG (Valentini et al., 2016).

FIGURE 2| Sampling of Trichogenes claviger during the eDNA fields. A. Live specimens in a field aquarium (Photo by J. P. Silva-Novelli). B–C. Collection in the RPPN Mata de Kaetés. D. Collection in Ribeirão Braço Sul.
The PCR program consisted of an initial cycle of denaturation at 97°C, followed by 35 cycles of denaturation at 95°C, girdling of the primers at 57°C and extension at 72°C. The success of the amplification was evaluated by electrophoresis on a 1% agarose gel. Samples that presented a sharp band with a size (pb) corresponding to the expected were selected for sequencing by the Sanger method for both fragment orientations forward (FWD) and reverse (REV). The quality of the sequences obtained from the FWD and REV amplicons was evaluated, and the sequences were subsequently merged to obtain the consensus sequence corresponding to the complete amplicon. The target 12S rRNA fragment spanned 650 bp and included the regions of three 12S rRNA markers: (1) NeoFish (~190 bp, Milan et al., 2020); (2) MiFish (~170 bp, Miya et al., 2015); and (3) Teleo (~70 bp, Valentini et al., 2016), except for the MiFish FWD and Teleo REV annealing regions. These mini-barcodes are currently in use for most eDNA metabarcoding studies targeting ichthyofauna, as they can amplify a hypervariable region of the 12S rRNA gene, which contains information to effectively identify the tested fish to taxonomic family, genus, and species (Miya et al., 2015). The newly generated 12S rRNA reference sequences for T. claviger are available in the NCBI nucleotidedatabase under the accession numbers PX556613 to PX556616 (Tab. S1).
Water sampling and visual survey, and fish collections inventory. A total of 10 streams were sampled following the protocol established by Mendes et al. (2024a). At each site water was collected using a 1L sterile disposable bag, and filtered through 3 Sterivex (Millipore)cartridges, with 180 ml of water per cartridge with the aid of disposable syringes. A total of three cartridges (replicates) were used at each site. Two filtration controls with sterile mineral water were performed alongside field sampling to monitor potential contamination during the filtration process. All eDNA samples were collected before any other procedure, with the sites intact. Immediately after filtration, the filter cartridges were emptied and filled with 2 ml of ATL buffer (Qiagen) for sample preservation.
After collecting the water samples, a visual survey was performed. Each site was observed for 30 min, paying special attention to preferred niches: puddle water with slow currents, as well as sandy and leaf litter substrates. This method was chosen because T. claviger inhabits shallow, clear-water headwater streams with sandy substrates and accumulated plant material, where visual detection is feasible and traditional seining is often impractical due to irregular topography and limited space. Visual observation has been effectively used in previous ecological studies of the species (Sarmento-Soares et al., 2018). The list of species previously detected for the Itapemirim basin was obtained from Sarmento-Soares et al. (2014), further referred to as fishcollections, and used for comparison with eDNA metabarcoding and visual survey.
eDNA extraction, amplification and sequencing. The eDNA was extracted from the filters using a protocol developed by the Conservation Genetics Laboratory (LGC) (Mendes et al., 2024b) with adjustments on the commercial DNeasy PowerWater Kit (Qiagen) such as initial step of DNA digestion with proteinase K and heating of elution buffer to 55ºC. Purified extracts were checked with DNA concentration (ng/μL) and absorbance ratios (A260/280) estimated via a Nanodrop 2000 (Thermo, Tab. S2). The 12S rRNA MiFish marker was amplified with the primers MiFish-U-F (5’-GTCGGTAAAACTCGTGCCAGC-3’) and MiFish-U-R (5′-CATAGTGGGGTATCTAATCCCAGTTTG-3′) (Miya et al., 2020), on a 25 µl final volume, using AmpliTaq Gold, with 2 µl DNA. The thermal cycles for this step were as follows: initial denaturation at 95°C for 2’, followed by 35 cycles of denaturation at 95°C for 1’, annealing at 60°C for 30’, and elongation at 72°C for 1’, followed by final elongation at 72°C for 7’. PCR negative controls were also included. PCR products were purified using Agencourt Ampure beads (Beckman Coulter) according to the manufacturer’s instructions and used as templates for the second PCR, the library construction. This second PCR was conducted with Illumina adapters and indexes (Nextera Index kit®) following manufacturer protocol instructions. The library was quantified by real-time PCR using a KAPA Biosystems Quantification Kit, and sequenced on an iSeq100 (Illumina) using an iSeq v. 2 300 cycle (2×150 bp) kit. For each sampling site, all PCR replicates for the three sampling replicates were pooled together into a single library per site. The average raw output per sample was 64,502 read pairs After bioinformatic processing, an average of 37,051 sequences per sample remained and were used for downstream analyses.
Bioinformatic and data analyses. Bioinformatics analyses were performed using a custom pipeline written in R v. 4.3.3 (R Development Core Team, 2024) based on Hilário et al. (2023). Primer and adapter removal was performed using the Cutadapt package (Martin, 2011). Low-quality reads filtering, reads merging into ASVs (Amplicon Sequence Variants) and chimeric sequences removal were performed using the DADA2 package (Callahan et al., 2016). ASVs were grouped into OTUs using SWARM v. 2 (Mahé et al., 2015) to evaluate the correlation between unique ASVs, OTUs and taxa on each sample.
All ASVs found in negative controls were completely removed from the other samples. All ASVs were taxonomically assigned using the BLASTr R package (Hilário et al., 2024), by performing a parallelized BLASTn (Camacho et al., 2009) against the NCBI core nucleotide database (Sayers et al., 2022, core_nt,updated June/2025) combined with a custom database composed of 12S rRNA sequences for some species of the Brazilian fish fauna (Hilário et al., 2023), along with the newly added reference sequences for T. claviger generated in this study. BLASTn searches were performed with identity and coverage cutoff values of 80%. All BLASTn matches with identity above 95% were evaluated for taxonomic assignment, revised by ichthyologists considering the local fauna, identity and coverage, and assigned to species, genus, or family taxa. All taxa with identitybelow 95% were discarded. Only taxa assigned to Actinopterii were kept on downstream analyses. Data analysis was performed using dplyr (Wickham et al., 2023) and images were generated using ggplot2 (Wickham, 2016). The Quantum-GIS v. 3.24.0 software was used to draw the map of geographic distribution.
Conservation action methods. To calculate AOO (Area of Occupation) and EOO (Extent of Occurrence), the Geospatial Conservation Assessment Tool (GeoCat, 2024), was employed regarding data on the occurrence of Trichogenes claviger. The EOO corresponds to the area contained in the smallest convex polygon including all the points of occurrence known for the species, while the AOO corresponds to the sum of the effective area of occurrence of the taxon within its EOO.
Results
From the total 645,028 raw sequences obtained for all samples, 33.91% (218,761 sequences) were kept in bioinformatic processment, comprising 151 unique ASVs assigned to Actinopterii. Reads counts for each sample along the quality control and pipeline steps can be found on Tab. S3. All raw reads files and the samples table are available at https://zenodo.org/ (DOI: 10.5281/zenodo.15230137). From these sequences, 8.76% (19,182 sequences) were assigned to T. claviger. This species was detected on the sampling sites P2, P4 and P8 using eDNA metabarcoding. The species was consistently detected across all three filtration replicates at each of these sites, supporting the reliability of the eDNA signal. It was the only species detected at these sites, except for the sampling site P8, where an ASV assigned to Pimelodidae was also detected. Concerning the visual survey, T. claviger was not detected at its original type locality, the sampling site P2, but it was spotted at the sites P4 and P8 (Fig. 3), showing the complementarity of these techniques.
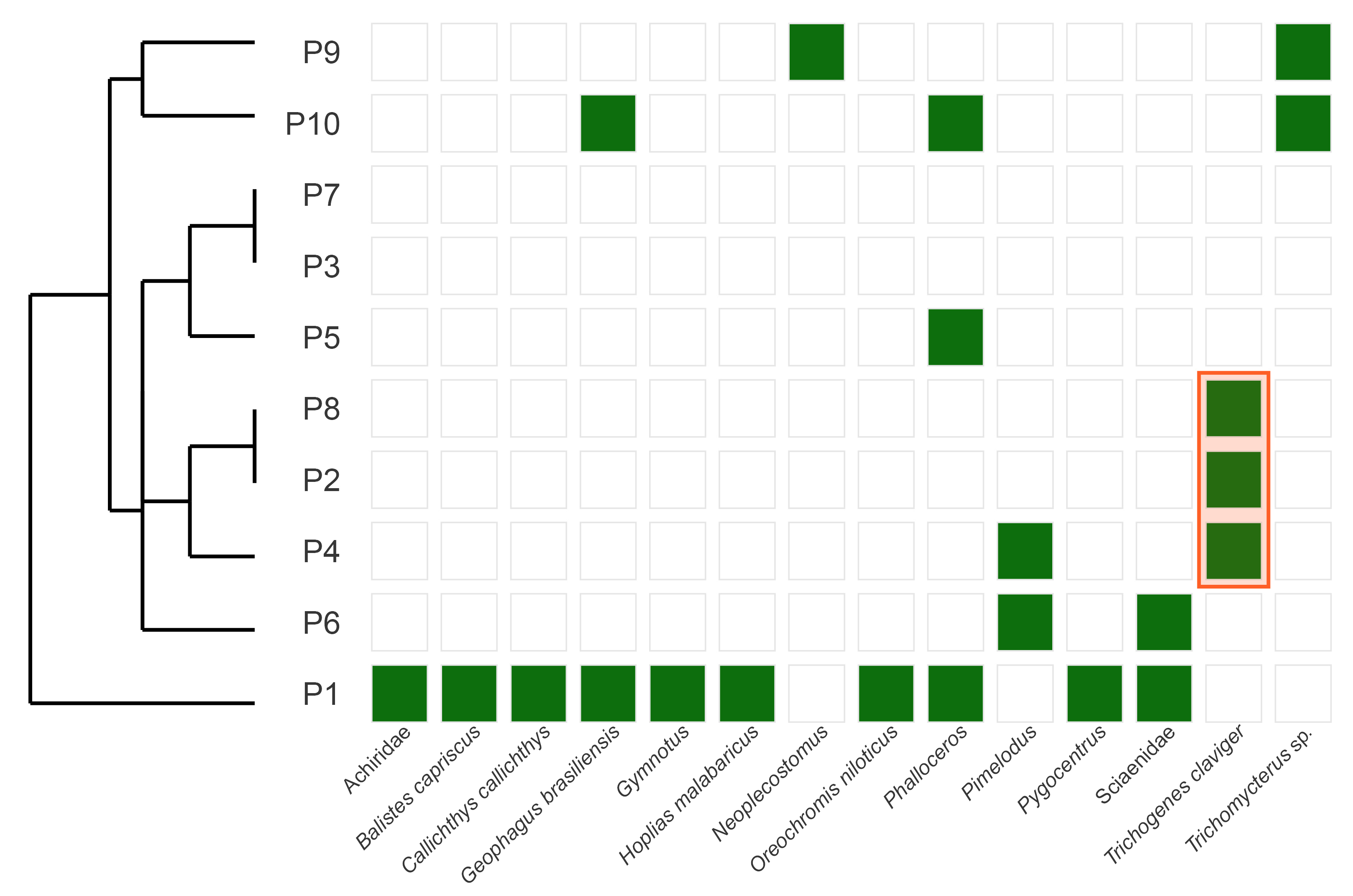
FIGURE 3| Hierarchical clustering dendrogram of eDNA sampling sites based on fish species presence/absence data. The dendrogram was generated using Euclidean distance and Ward’s linkage method. Trichogenes claviger was detected only at sample sites P2, P4 and P8. Site P1 was the sample site with the highest number of species.
In addition to the species T. claviger, 14 other fish species were detected in water samples when considering all sampling sites (Tab. 1). All species detected via eDNA metabarcoding were represented by reference sequences in our curated 12S rRNA database, which included entries from the NCBI core nucleotide collection supplemented with a custom database for Brazilian freshwater fish (Hilário et al., 2023) and the newly generated references for T. claviger. This ensured reliable taxonomic assignment for all reported detections.The species detected in this study belong to six orders, 10 families and 14 genera. The order Siluriformes was the most representative, comprising five species, followed by two species of Perciformes, two of Cichliformes, two species of Characiformes and one species each of Cyprinodontiformes, Gymnotiformes and Pleuronectiformes. All species detected via eDNA metabarcoding were represented by reference sequences in our curated 12S rRNA database, which included entries from the NCBI core nucleotide collection supplemented with a custom database for Brazilian freshwater fish (Hilário et al., 2023) and the newly generated references for T. claviger. This ensured reliable taxonomic assignment for all reported detections.
TABLE 1 | List of species detected by each sampling method: Fish collections, Visual Survey and eDNA metabarcoding. *Refer to fish species used as food source; # Refer to marine fish species.
ORDER/Family/Species | Sampling method | ||
Fish collections | eDNA | Visual Survey | |
CHARACIFORMES | |||
Acestrorhamphidae | |||
Astyanax microschemos Bertaco & Lucena, 2006 | X |
|
|
Deuterodon giton (Eigenmann, 1908) | X |
|
|
Deuterodon janeiroensis (Eigenmann, 1908) | X |
|
|
Deuterodon parahybae Eigenmann, 1908 | X |
|
|
Crenuchidae | |||
Characidium vidali Travassos, 1967 | X |
|
|
Erythrinidae | |||
Hoplias malabaricus (Bloch, 1794)* |
| X |
|
Serrasalmidae | |||
Pygocentrus sp.* |
| X |
|
CYPRINODONTIFORMES | |||
Poeciliidae | |||
Phalloceros harpagos Lucinda, 2008 | X | X |
|
Poecilia reticulata Peters, 1859 | X |
|
|
GYMNOTIFORMES | |||
Gymnotidae | |||
Gymnotus carapo Linnaeus, 1758 | X | X |
|
PERCIFORMES | |||
Balistidae | |||
Balistes capriscus Gmelin, 1789*# |
| X |
|
CICHLIFORMES | |||
Cichlidae | |||
Geophagus brasiliensis (Quoy & Gaimard, 1824) | X | X |
|
Oreochromis niloticus (Linnaeus, 1758)* |
| X |
|
Sciaenidae | |||
Plagioscion sp.*# |
| X |
|
PLEURONECTIFORMES | |||
Achiridae | |||
Hypoclinemus mentalis (Günther, 1862)*# |
| X |
|
SILURIFORMES | |||
Callichthyidae | |||
Callichthys callichthys (Linnaeus, 1758) |
| X |
|
Loricariidae | |||
Hypostomus affinis (Steindachner, 1877) | X |
|
|
Neoplecostomus microps (Steindachner, 1877) | X | X |
|
Pimelodidae | |||
Pimelodidae sp. |
| X |
|
Heptapteridae | |||
Rhamdia quelen (Quoy & Gaimard, 1824) |
|
| X |
Trichomycteridae | |||
Trichogenes claviger de Pinna, Helmer, Britski & Nunes, 2010 | X | X | X |
Trichomycterus caudofasciatus Alencar & Costa, 2004 | X | X |
|
Sampling methods comparison. When we analyzed the three sampling methods, eDNA, visual survey and previous fish inventories even though they did not cover the same stretches, we observed that the estimates of species composition per sampled stream demonstrated that the fish collection method and eDNA complemented each other; considering that of the 12 species reported within the region on previous fish inventories, five were detected by eDNA metabarcoding (41.6%), and 2 fresh water species (40%) were detected only by eDNA (Tab. 1), namely Pimelodidaesp. and Callichthys callichthys. This demonstrates that eDNA metabarcoding should not be used as a replacement, and combining both strategies would maximize the comprehension of local ichthyofauna. Nonetheless, the fact that most previously known species were detected by eDNA metabarcoding with a small sampling effort highlights the power of this technique. The visual survey was only able to detect two species.
Updated Trichogenes claviger distribution. Our results expand the known distributional area of T. claviger. The species was previously known to occur only at its type locality at the Picada Comprida stream (Point P4; Fig. 1B), and one of its tributaries (non-type material collected with traditional methods marked as black dots in Fig. 1B).
Environmental DNA detected presence is assigned to Fig. 1B (yellow X) in the following three collecting localities P2, P4 and P8. Absence of detection of its presence signed in Fig. 1B (blue X). In this case, the tentative was not successful in collecting localities P1, P3, P5, P6, P7, P9 and P10 (see Fig. 1B). The results illustrated that T. claviger is still present in the region delimited for this study, and its detection in the samples expanded the distribution of the species to the ribeirão Braço Sul (Point 8). In calculating the Extension of Occurrence (EOO) that was previously shorter (IUCN, 2022) and now has 3,007 km2 and a revised area of occupation (AOO) of 12 km2 (Fig. 4), calculated by the GeoCAT system.
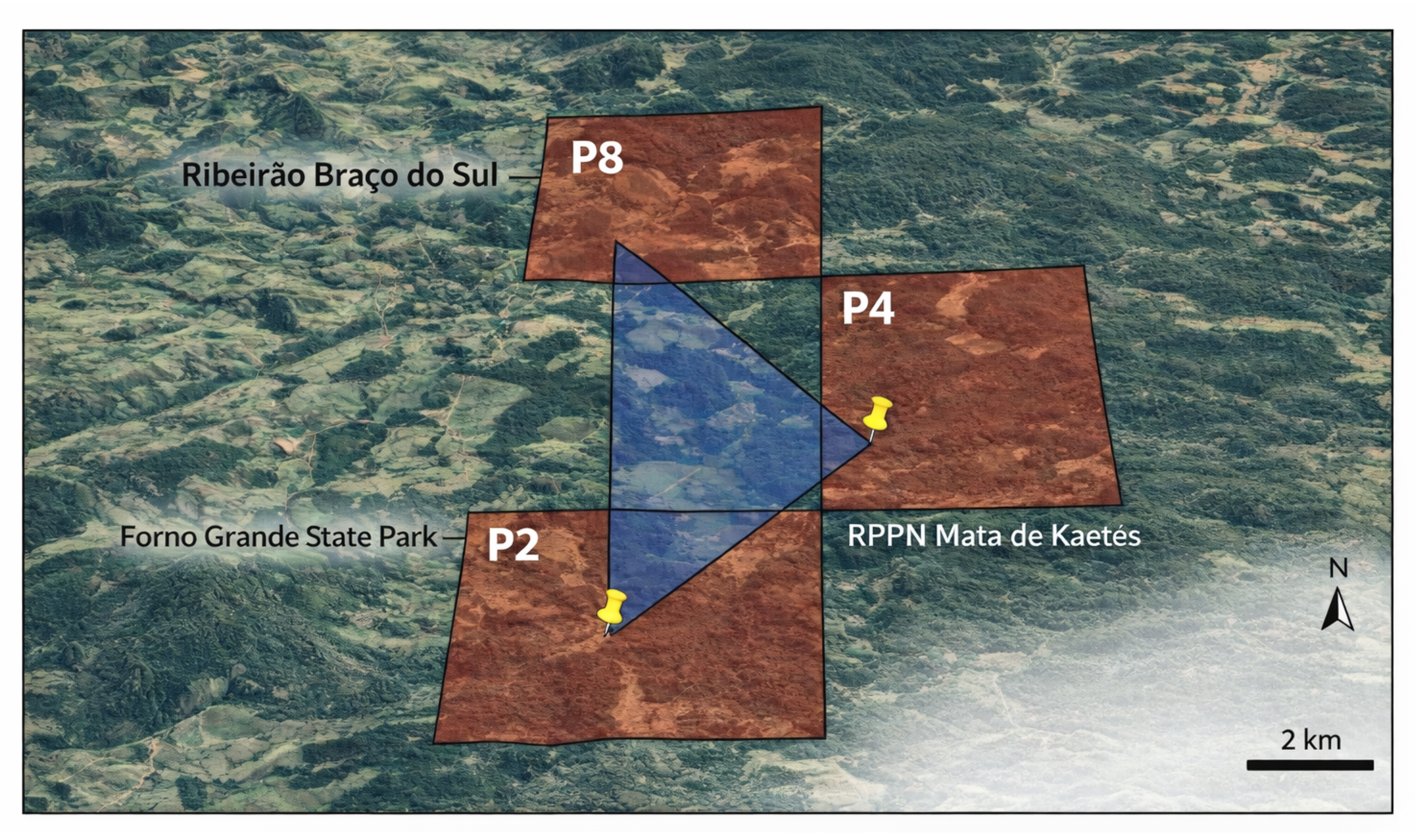
FIGURE 4| Map of the result of the minimum convex polygon, with the GeoCAT, IUCN KEW assessment system, in order to categorize the risk of extinction of Trichogenes claviger AOO 12 km2 and EOO 3,021 km2. See text for further details. P2 at the Picada Comprida stream; P4 on right-bank tributaries of ribeirão Caetés; P8 on a right-bank tributary of ribeirão Braço Sul.
Conservation concerns. We identified an area of about 3 km² of EOO for the species. As T. claviger has an extremely restricted distribution, it still fits in the Critically Endangered (CR) category. The AOO was 12 km², which would classify the species as Endangered (EN). Despite the amplification of the AOO through eDNA detection, the species remains as Critically Endangered, deserving a very restrictive distributional area in mountainous slopes at southern Espírito Santo, Brazil. The few known records (< 5 locations) assert T. claviger as an endemic species of the Itapemirim basin. Although investigated, no records were achieved at the neighbor basin headwaters, at the rio Jucu. The results obtained corroborate the current threat status of the species (IUCN, 2022). Even with an increase in the locations of occurrence, the species continues to have environments restricted to the mountain foothills at southern Espírito Santo. The Point 8, located in ribeirão Braço Sul, in addition to being a short expansion of the distribution of the species, reveals that Trichogenes claviger is endemic to one single river sub-basin at upper rio Itapemirim.
Discussion
Our target species, Trichogenes claviger, is known to inhabit small, high-altitude streams. These aquatic environments shelter small-sized fishes with limited dispersal potential, high endemism, and complex ecological interactions (Abilhoa et al., 2011; Castro, Polaz, 2020). In low-order streams with clear water, shallow depth, and heterogeneous substrates (pebbles, submerged wood, sand, leaf litter), detecting local species using conventional methods is particularly challenging. How extensive is the species’ distributional range? Can we predict it through alternative methods?
The successful detection of T. claviger, a rare and critically endangered species, using a multi-taxa eDNA metabarcoding approach underscores the sensitivity and applicability of this method for conservation monitoring. Environmental DNA has emerged as a non-destructive alternative to traditional, often destructive and labor-intensive freshwater monitoring methods (Cilleros et al., 2019). Our methodological choices, including filtering a substantial water volume (3 × 180 mL per site) and processing triplicate filtration replicates, were critical to increasing detection probability for low-abundance eDNA sources (Deiner et al., 2015; Wood et al., 2020). The consistent detection of T. claviger across all technical replicates at positive sites (P2, P4, P8) reinforces the robustness of the eDNA signal and aligns with recommendations for standardized protocols in rare species surveillance (Cooper et al., 2022). This finding adds to a growing body of literature demonstrating the efficacy of eDNA metabarcoding for detecting elusive freshwater fishes, including other range-restricted and threatened species in diverse ecosystems (e.g., Thomsen et al., 2012b; Stoeckle et al., 2015; Baker et al., 2023).
The detection sensitivity of eDNA-based approaches has proven effective for monitoring endangered or elusive aquatic species, including freshwater fishes with restricted distributions (Thomsen et al., 2012a; Miya et al., 2015; Stoeckle et al., 2015; Holmes et al., 2024; Brys et al., 2021; Baker et al., 2023; Carvalho, Leal, 2023). Although eDNA detection probability is generally higher for species with larger populations, and can be influenced by body size and behavior (Thomsen et al., 2012a; Pilliod et al., 2013), our results demonstrate that eDNA metabarcoding is also a powerful tool for detecting rare, range-restricted species. This was corroborated by the congruence between eDNA signals and visual surveys where individuals were observed swimming. Moreover, eDNA succeeded in detecting the species at its type locality (site P4) where no individuals were sighted during our sampling, underscoring the method’s sensitivity.
In the headwaters of the rio Itapemirim, eDNA facilitated the rapid detection of both endangered and allochthonous species, reducing sampling effort and minimizing impacts on these fragile forested streams (Deiner et al., 2017; Schenekar et al., 2020). The technique proved sensitive for tracking fish assemblages in these environments (Thomsen et al., 2012b; Lacoursière-Roussel et al., 2018), though detecting the target species in neighboring headwaters remains a task for future surveys.
The eDNA metabarcoding results revealed that Trichogenes claviger was frequently the only species detected at sites P2 and P4, and one of only two at P8. This low species richness aligns with the expected pattern for first- and second-order headwater streams in mountainous regions of the Atlantic Forest. Such environments are typically characterized by stringent physical conditions (e.g., higher slope, less stable substrate, lower temperatures) and lower resource availability, resulting in naturally depauperate fish communities dominated by a few specialized species (Abilhoa et al., 2011; Castro, Polaz, 2020). This phenomenon, described by the River Continuum Concept (Vannote et al., 1980), is also observed in other coastal basins of the region, such as the upper Parati-Mirim River (habitat of T. longipinnis; Sazima, 2004) and the upper Doce River (where T. beagle occurs; Santos et al., 2025), both of which harbor similarly low-diversity fish assemblages.
From an ecological perspective, the apparent dominance or exclusive occurrence of T. claviger in some reaches may reflect a combination of habitat specialization, such as the use of marginal microhabitats with leaf-litter accumulations, and reduced competition or predation pressure owing to the naturally species-poor community. However, this ecological simplicity also renders these populations extremely vulnerable. Habitat disturbances (e.g., riparian deforestation, siltation, or introduction of non-native species) can have disproportionate impacts, because no more diverse fish assemblage exists to act as a functional buffer or a source for recolonization.
Our field observations further illustrate the specific habitat associations of T. claviger. The species was found in small riffles within forested areas above 1,000 m elevation in the upper ribeirão Braço Sul and rio Fruteiras headwaters (Itapemirim basin). At site P2 (Córrego Picada Comprida, RPPN Mata de Kaetés), individuals were observed swimming near the margin under the shade of large Eucalyptus trees, approximately 50 m from plantations (elevation 1,145 m). The fast-flowing, transparent water at sites P4 (type locality) and P8 (Ribeirão Braço Sul) allowed visual confirmation of actively swimming individuals, with site P8 appearing to function as a nursery, hosting dozens of juveniles and sub-adults. Notably, at all sites where T. claviger was detected, it was the only fish recorded in the immediate environment and was a common component of the assemblage. This pattern of isolated headwater populations mirrors that reported for congeners T. longipinnis (Sazima, 2004) and T. beagle (Santos et al., 2025).
The rapid advancement of genomic technologies is promising. eDNA not only aided in detecting T. claviger but also provides a framework for continuous monitoring of a species previously known only from its type locality. The 12S rRNA marker reference developed here for T. claviger proved effective, highlighting the importance of generating reference sequences, especially for endangered species inhabiting sensitive environments, to unlock their detection via eDNA metabarcoding.
Some species detected by eDNA metabarcoding were unexpected in freshwater ecosystems, belonging instead to marine habitats. All of these marine taxa are commonly used as food sources and are commercially available (Fig. 3). Importantly, they were detected exclusively at site P1, the most anthropogenically disturbed site, located close to human settlements. This finding underscores the high sensitivity of the eDNA approach and its potential to trace human-mediated subsidies or contamination in aquatic systems.
In this context, our results reinforce the need for headwater-specific conservation strategies. Protecting these environments goes beyond preserving individually threatened species; it is about safeguarding unique ecological processes and simplified yet functionally specialized communities that are irreplaceable for the integrity of the river continuum. eDNA-based monitoring programs, as demonstrated here, are promising tools for tracking not only the presence of target species but also compositional changes in these sensitive communities over time. Our results may be used by Brazilian environmental agencies, in partnership with researchers and local communities, to prioritize and implement timely conservation actions that ensure the protection of this delicate environment and its endemic species.
Material examined. Brazil. Trichogenes claviger. Espírito Santo State, rio Itapemirim. MBML 3289, holotype, 50.4 mm SL, MBML 3290, 11 paratypes, 43.2–14.8 mm SL, tributary of córrego Picada Comprida. MBML 3987, 13, 36.7–23.7 mm SL, córrego Picada Comprida. MZUSP 124882, 33 (2 cleared and stained), 49.9–25.5 mm SL, tributary of córrego Picada Comprida. MBML 14089, 1, 31.4 mm SL, MBML 14091, 8, 48.8–30.2 mm SL, córrego Picada Comprida. MBML 14206, 10, 38.9–29.2 mm SL, MBML 14207, 3, 33.7–30.1 mm SL, tributary of rio Braço Sul.
Genetic samples (Tissue-TS). Brazil,Espírito Santo State,rio Itapemirim. LGC 8125 (TS); LGC 8126 (TS); LGC 8127 (TS); LGC 8128 (TS); LGC 8129 (TS).
Acknowledgments
This study was taken as part of the master dissertation of JPS, at Programa de Biologia Animal, Universidade Federal do Espírito Santo, and all research teams contributed as authors. We would like to thank Franciele Loss Pugnal of the Museu de Biologia Mello Leitão/Instituto Nacional da Mata Atlântica for her collaborative support in accessing the collection, as well as our colleagues Bruno F. E. Novelli, Joelcio Freitas and Lorena Tonini for their field activities. We would like to thank IMD (Marcos Daniel Institute), represented by Gustavo Magnago, for all the support during the field expeditions. We would also like to thank the Faccini family for allowing us to collect on their property and for supporting us during the field trips. We are grateful for all the support from the team at the Conservation Genetics Laboratory (LGC) at PUC/MG, with the bioinformatic analyses that were performed on theeDNA server, hosted at the Conservation Genetics Laboratory, Post-Graduate Program on Biodiversity and Environment, Pontifícia Universidade Católica de Minas Gerais. We are also grateful for the support from LGC students Higor H. A. Nascimento, Lívia C. M. Silva, Gabriel A. Mendes, Júlia Fonseca and Guilherme C. Berger, for their support in the laboratory and on field expeditions. Our gratitude to the Instituto Nossos Riachos laboratory and partnership during research analysis.
References
Abilhoa V, Braga RR, Bornatowski H, Vitule JRS. Fishes of the Atlantic Rain Forest streams: ecological patterns and conservation. In: Grilo O, Venora G, editors. Changing diversity in changing environment: IntechOpen; 2011. p.259–82. Available from: https://www.intechopen.com/chapters/23585
Aljanabi SM, Martinez I. Universal and rapid salt-extraction of high-quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997; 25(22):4692–93. https://doi.org/10.1093/nar/25.22.4692
Baker CS, Claridge D, Dunn C, Fetherston T, Baker DN, Klinck H et al. Quantification by droplet digital PCR and species identification by metabarcoding of environmental (e)DNA from Blainville’s beaked whales, with assisted localization from an acoustic array. PLoS ONE. 2023; 18(9):e0291187. https://doi.org/10.1371/journal.pone.0291187
Biggs J, Ewald N, Valentini A, Gaboriaud C, Dejean T, Griffiths RA et al. Using eDNA to develop a national citizen science-based monitoring programme for the great crested newt (Triturus cristatus). Biol Conserv. 2015; 183:19–28. https://doi.org/10.1016/j.biocon.2014.11.029
Brys R, Haegeman A, Halfmaerten D, Neyrinck S, Staelens A, Auwerx J et al. Monitoring of spatiotemporal occupancy patterns of fish and amphibian species in a lentic aquatic system using environmental DNA. Mol Ecol. 2021; 30(13):3097–110. https://doi.org/10.1111/mec.15742
Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJA, Holmes SP. DADA2: high-resolution sample inference from Illumina amplicon data. Nat Methods. 2016; 13(7):581–83. https://doi.org/10.1038/nmeth.3869
Camacho C, Coulouris G, Avagyan V, Ma N, Papadopoulos J, Bealer K et al. BLAST+: architecture and applications. BMC Bioinformatics. 2009; 10:421. https://doi.org/10.1186/1471-2105-10-421
Carvalho DC, Leal CG. Steps forward in biomonitoring 2.0: eDNA Metabarcoding and community-level modelling allow the assessment of complex drivers of Neotropical fish diversity. Glob Change Biol. 2023; 29(7):1688–90. https://doi.org/10.1111/gcb.16596
Castro RMC, Polaz CNM. Small-sized fish: the largest and most threatened portion of the megadiverse Neotropical freshwater fish fauna. Biota Neotrop. 2020; 20(1):e20180683. https://doi.org/10.1590/1676-0611-BN-2018-0683
Cilleros K, Valentini A, Allard L, Dejean T, Etienne R, Grenouillet G et al. Unlocking biodiversity and ecological studies in high diversity environments using environmental DNA (eDNA): a test with Guianese freshwater fishes. Mol Ecol Resour. 2019; 19(1):27–46. https://doi.org/10.1111/1755-0998.12900
Cooper MK, Villacorta-Rath C, Burrows D, Jerry DR, Carr L, Barnett A et al. Practical eDNA sampling methods inferred from particle size distribution and comparison of capture techniques for a critically endangered elasmobranch. Environ DNA. 2022; 4(5):1011–23. https://doi.org/10.1002/edn3.279
Cristescu ME, Hebert PD. Uses and misuses of environmental DNA in biodiversity science and conservation. Annu Rev Ecol Evol Syst. 2018; 49:209–30. https://doi.org/10.1146/annurev-ecolsys-110617-062306
Deiner K, Walser J-C, Mächler E, Altermatt F. Choice of capture and extraction methods affect detection of freshwater biodiversity from environmental DNA. Biol Conserv. 2015; 183:53–63. https://doi.org/10.1016/j.biocon.2014.11.018
Deiner K, Bik HM, Mächler E, Seymour M, Lacoursière-Roussel A, Altermatt F et al. Environmental DNA metabarcoding: Transforming how we survey animal and plant communities. Mol Ecol. 2017; 26(21):5872–95. https://doi.org/10.1111/mec.14350
DoNascimiento C. Morphological evidence for the monophyly of the subfamily of parasitic catfishes Stegophilinae (Siluriformes, Trichomycteridae) and phylogenetic diagnoses of its genera. Copeia. 2015; 103(4):933–60. https://doi.org/10.1643/CI-14-132
Stoeckle BC, Kuehn R, Geist J. Environmental DNA as a monitoring tool for the endangered freshwater pearl mussel (Margaritifera margaritifera L.): a substitute for classical monitoring approaches? Aquat Conserv. Mar. Freshw. Ecosyst 2015; 26(6):1–10. https://doi.org/10.1002/aqc.2611
Ferraris Jr. CJ. Checklist of catfishes, recent and fossil (Osteichthyes: Siluriformes), and catalog of siluriform primary types. Zootaxa. 2007; 1418:1–628. https://doi.org/10.11646/zootaxa.1418.1.1
Fricke R, Eschmeyer WN, Van der Laan R. Eschmeyer’s catalog of fishes: genera, species, references [Internet]. San Francisco: California Academy of Sciences; 2024. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Geospatial Conservation Assessment Tool (GeoCAT). 2025. Available from: https://geocat.iucnredlist.org/
Goldberg CS, Strickler KM, Pilliod DS. Moving environmental DNA methods from concept to practice for monitoring aquatic macroorganisms. Biol Conserv. 2015; 183:1–3. https://doi.org/10.1016/j.biocon.2014.11.040
Goldberg CS, Turner CR, Deiner K, Klymus KE, Thomsen PF, Murphy MA et al. Critical considerations for the application of environmental DNA methods to detect aquatic species. Methods Ecol Evol. 2016; 7(11):1299–307. https://doi.org/10.1111/2041-210X.12595
Grandcolas P, Trewick SA. What is the meaning of extreme phylogenetic diversity? The case of phylogenetic relict species. In: Pellens R, Grandcolas P, editors. Biodiversity conservation and phylogenetic systematics. Cham: Springer; 2016. p.99–115.
Hennig W. Phylogenetic systematics. Urbana: University of Illinois Press; 1966.
Hilário HO, Mendes IS, Sales NG, Carvalho DC. DNA metabarcoding of mock communities highlights potential biases when assessing Neotropical fish diversity. Environ DNA. 2023; 5(6):1351–61. https://doi.org/10.1002/edn3.456
Hilário HO, Queiroz LR, Brito GAM, Carvalho DC. BLASTr: an R package for parallel taxonomic classification of metabarcoding sequences [Internet]. 2024. Available from: https://github.com/heronoh/BLASTr
International Union for Conservation of Nature (IUCN). The IUCN Red List of Threatened Species. Version 2022-2 [Internet]. 2022. Available from: https://www.iucnredlist.org
Krell FT, Cranston PS. Which side of the tree is more basal? Syst Entomol. 2004; 29(3):279–81. https://doi.org/10.1111/j.0307-6970.2004.00262.x
Lacoursière-Roussel A, Howland K, Normandeau E, Grey EK, Archambault P, Deiner K et al. eDNA metabarcoding as a new surveillance approach for coastal Arctic biodiversity. Ecol Evol. 2018; 8(16):7763–77. https://doi.org/10.1002/ece3.4213
Holmes AE, Baerwald MR, Rodzen J, Schreier BM, Mahardja B, Finger AJ. Evaluating environmental DNA detection of a rare fish in turbid water using field and experimental approaches. PeerJ. 2024; 12:e16453. doi: 10.7717/peerj.16453.
Lodge DM, Turner CR, Jerde CL, Barnes MA, Chadderton L, Egan SP et al. Conservation in a cup of water: estimating biodiversity and population abundance from environmental DNA. Mol Ecol. 2012; 21(11):2555–58. https://doi.org/10.1111/j.1365-294X.2012.05600.x
Mahé F, Rognes T, Quince C, Vargas C, Dunthorn M. Swarm v2: highly-scalable and high-resolution amplicon clustering. PeerJ. 2015; 3:e1420. https://doi.org/10.7717/peerj.1420
Martin M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011; 17(1):10–12. https://doi.org/10.14806/ej.17.1.200
Mendes GA, Affonseca JG, Mendes IS, Hilário HO, Carvalho DC, Berger GC. eDNA extraction from MCE filter and Sterivex cartridge-LGC PUC Minas [Internet]. 2024a. https://doi.org/10.17504/protocols.io.14egn2m9zg5d/v1
Mendes GA, Affonseca JG, Mendes IS, Hilário HO, Carvalho DC, Berger GC. Protocolo de coleta de amostras para eDNA metabarcoding usando filtros Sterivex – LGC PUC Minas [Internet]. 2024b. https://doi.org/10.17504/protocols.io.81wgbz623gpk/v1
Milan DT, Mendes IS, Damasceno JS, Teixeira DF, Sales NG, Carvalho DC. New 12S metabarcoding primers for enhanced Neotropical freshwater fish biodiversity assessment. Sci Rep. 2020; 10:17966. https://doi.org/10.1038/s41598-020-74902-3
Miya M, Sato Y, Fukunaga T, Sado T, Poulsen JY, Sato K et al. MiFish, a set of universal PCR primers for metabarcoding environmental DNA from fishes: detection of more than 230 subtropical marine species. R Soc Open Sci. 2015; 2(7):150088. https://doi.org/10.1098/rsos.150088
Miya M, Gotoh RO, Sado T. MiFish metabarcoding: a high-throughput approach for simultaneous detection of multiple fish species from environmental DNA and other samples. Fish Sci. 2020; 86:939–70. https://doi.org/10.1007/s12562-020-01461-x
Ministério do Meio Ambiente (MMA). Portaria MMA Nº 445, de 17 de dezembro de 2014. Lista Nacional Oficial de Espécies da Fauna Ameaçadas de Extinção. Brasília: Diário Oficial da União; 2014.
Ministério do Meio Ambiente/Instituto Chico Mendes de Conservação da Biodiversidade (MMA/ICMBio). Portaria Nº 370, de 1 de agosto de 2019. Plano de Ação Nacional para a Conservação de Espécies de Peixes e Eglas Ameaçados de Extinção da Mata Atlântica – PAN Peixes e Eglas da Mata Atlântica. Brasília: Diário Oficial da União; 2019.
De la Ossa-Guerra LE, Limeira DM, Dalcin RH, Hilário HO, Carvalho DC, Artoni RF. Illuminating the dark: using eDNA metabarcoding to study the unique ichthyofauna of the Furnas Formation Sinkholes. Environ DNA. 2025; 7(4):e70156. https://doi.org/10.1002/edn3.70156
Pilliod DS, Goldberg CS, Arkle RS, Waits LP. Estimating occupancy and abundance of stream amphibians using environmental DNA from filtered water samples. Can J Fish Aquat Sci. 2013; 70(8):1123–30. https://doi.org/10.1139/cjfas-2013-0047
Rizzato PP, Bichuette ME. Ituglanis boticario, a new troglomorphic catfish (Teleostei: Siluriformes: Trichomycteridae) from Mambaí karst area, central Brazil. Zoologia (Curitiba). 2014; 31(6):577–98. https://doi.org/10.1590/S1984-46702014000600006
Sabaj MH. Codes for natural history collections in ichthyology and herpetology (online supplement). Version 9.7. Washington, DC: American Society of Ichthyologists and Herpetologists; 2025. Available from: https://asih.org
Santos SAD, Reis VJC, Marques IDCC, Pessali TC, Britski HA, de Pinna MCC. Rediscovery of Trichogenes beagle de Pinna, Reis & Britski, 2020 (Siluriformes: Trichomycteridae) with comments on habitat and conservation status. Zootaxa. 2025; 5601(1):196–200. https://doi.org/10.11646/zootaxa.5601.1.13
Sarmento-Soares LM, Martins-Pinheiro RF. A fauna de peixes nas bacias do sul do Espírito Santo, Brasil. Sitientibus Sér Ciênc Biol. 2014; 13:1–37.
Sarmento-Soares LM, Martins-Pinheiro RF, Ferreira Martins LS, Nunes S, Helmer JL. Trichogenes claviger um peixinho capixaba criticamente ameaçado de extinção: “Caetés” uma Unidade de Conservação que pode protegê-lo. Bol Soc Bras Ictiol. 2018; 127:13–19.
Sayers EW, Bolton EE, Brister JR, Canese K, Chan J, Comeau DC et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022; 50(D1):20–26. https://doi.org/10.1093/nar/gkab1112
Sazima I. Natural history of Trichogenes longipinnis, a threatened trichomycetrid catfish endemic to Atlantic Forest streams in southeast Brazil. Ichthyol Explor Freshw. 2004; 15(1):49–60.
Schenekar T, Schletterer M, Lecaudey LA, Weiss SJ. Reference databases, primer choice, and assay sensitivity for environmental metabarcoding: lessons learnt from a re-evaluation of an eDNA fish assessment in the Volga headwaters. River Res Appl. 2020; 36(7):1004–13. https://doi.org/10.1002/rra.3610
Takahashi M, Saccò M, Kestel JH, Nester G, Campbell MA, van der Heyde M et al. Aquatic environmental DNA: a review of the macro-organismal biomonitoring revolution. Sci Total Environ. 2023; 873:162322. https://doi.org/10.1016/j.scitotenv.2023.162322
Thomsen PF, Kielgast J, Iversen LL, Moller PR, Rasmussen M, Willerslev E. Detection of a diverse marine fish fauna using environmental DNA from seawater samples. PLoS ONE. 2012a; 7(8):e41732. https://doi.org/10.1371/journal.pone.0041732
Thomsen PF, Kielgast J, Iversen LL, Wiuf C, Rasmussen M, Gilbert MTP et al. Monitoring endangered freshwater biodiversity using environmental DNA. Mol Ecol. 2012b; 21(11):2565–73. https://doi.org/10.1111/j.1365-294X.2011.05418.x
Thomsen PF, Willerslev E. Environmental DNA – An emerging tool in conservation for monitoring past and present biodiversity. Biol Conserv. 2015; 183:4–18. https://doi.org/10.1016/j.biocon.2014.11.019
Valentini A, Taberlet P, Miaud C, Civade R, Herder J, Thomsen PF et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol Ecol. 2016; 25(4):929–42. https://doi.org/10.1111/mec.13428
Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE. The river continuum concept. Can J Fish Aquat Sci. 1980; 37(1):130–37. https://doi.org/10.1139/f80-017
Wickham H. ggplot2: Elegant Graphics for Data Analysis. New York: Springer-Verlag; 2016.
Wickham H, François R, Henry L, Müller K, Vaughan D. dplyr: A Grammar of Data Manipulation. R package version 1.1.4 [Internet]. 2023. Available from: https://CRAN.R-project.org/package=dplyr
Wiley EO, Lieberman BS. Phylogenetics: Theory and Practice of Phylogenetic Systematics. 2nd ed. Hoboken: Wiley-Blackwell; 2011.
Wood ZT, Erdman BF, York G, Trial JG, Kinnison MT. Experimental assessment of optimal lotic eDNA sampling and assay multiplexing for a critically endangered fish. Environ DNA. 2020; 2(4):407–17. https://doi.org/10.1002/edn3.64
Authors

![]() Juliana Paulo da Silva-Novelli1,2,
Juliana Paulo da Silva-Novelli1,2, ![]() Heron Oliveira Hilário3,
Heron Oliveira Hilário3, ![]() Daniel Cardoso Carvalho3,
Daniel Cardoso Carvalho3, ![]() Ronaldo Fernando Martins-Pinheiro4 and
Ronaldo Fernando Martins-Pinheiro4 and ![]() Luisa Maria Sarmento-Soares1,4,5
Luisa Maria Sarmento-Soares1,4,5 ![]()
[1] Programa de Pós-Graduação em Ciências Biológicas (Biologia Animal), Universidade Federal do Espírito Santo (UFES), Campus Goiabeiras, Av. Fernando Ferrari, 514, Goiabeiras, 29075-910, Vitória, ES, Brazil. (JPS) 1414juliana@gmail.com, (LMSS) sarmento.soares@gmail.com (corresponding author).
[2] Instituto Nacional da Mata Atlântica (INMA), Av. José Ruschi, 4, 29650-000, Santa Teresa, ES, Brazil
[3] Laboratório de Genética da Conservação, Programa de Pós-Graduação em Biodiversidade e Meio Ambiente, Pontifícia Universidade Católica de Minas Gerais, Belo Horizonte, Brazil. (HOH) heronoh@gmail.com, (DCC) danielcarvalho@pucminas.br.
[4] Instituto Nossos Riachos, Estrada de Itacoatiara, 356, 24348-095, Niterói, RJ, Brazil. (RFMP) pinheiro.martins@gmail.com.
[5] Universidade Estadual de Feira de Santana, Departamento de Ciências Biológicas, Av. Universitária s/n, Novo Horizonte, 44036 900, Feira de Santana, BA, Brazil.
Authors’ Contribution 

Juliana Paulo da Silva: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration.
Heron Oliveira Hilário: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Writing-original draft.
Daniel Cardoso Carvalho: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Resources, Software, Validation, Writing-original draft.
Ronaldo Fernando Martins-Pinheiro: Formal analysis, Investigation, Methodology, Software, Validation, Visualization, Writing-original draft, Writing-review and editing.
Luisa Maria Sarmento-Soares: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Ethical Statement
Sistema de Autorização e Informação sobre Biodiversidade (SISBIO) for the collection license (registration 79046–1/2).
Competing Interests
The author declares no competing interests.
Data availability statement
The authors confirm that the data supporting the findings of this study are available within the article.
AI statement
The authors did not use any AI-assisted technologies in the creation of this manuscript or its figures.
Funding
This study was financed in part by the Fundo Brasileiro para a Biodiversidade (FUNBIO), through the grants program FUNBIO – Conservando o Futuro. The Mohamed bin Zayed Species Conservation Fund. The Conservation Genetics Laboratory, Post-Graduate Program on Biodiversity and Environment, Pontifícia Universidade Católica de Minas Gerais as well as a scholarship provided by Fundação de Amparo à Pesquisa e Inovação do Espírito Santo (FAPES process 179/2021). CNPq providing a research fellowship to DCC (312102/2022–4) and Ecomol for conducting molecular analysis.
Supplementary Material
Supplementary material SUP
How to cite this article
Silva-Novelli JP, Hilário HO, Carvalho DC, Martins-Pinheiro RF, Sarmento-Soares LM. Detection of the critically endangered catfish Trichogenes claviger (Siluriformes: Trichomycteridae) using environmental DNA. Neotrop Ichthyol. 2026; 24(2):e260013. https://doi.org/10.1590/1982-0224-2026-0013
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2025 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted February 16, 2026
Accepted February 16, 2026
![]() Submitted April 15, 2025
Submitted April 15, 2025
![]() Epub June 19, 2026
Epub June 19, 2026