![]() Josué Sarino-Araújo1
Josué Sarino-Araújo1 ![]() ,
, ![]() Cárlison Silva-Oliveira2,
Cárlison Silva-Oliveira2, ![]() Flávio C. T. Lima3,
Flávio C. T. Lima3, ![]() André L. C. Canto4 and
André L. C. Canto4 and ![]() Frank R. V. Ribeiro1,4
Frank R. V. Ribeiro1,4
PDF: Download Here | Cite this article
Associate Editor: ![]() Juan Mirande
Juan Mirande
Section Editor: ![]() Bruno Melo
Bruno Melo
Editor-in-chief: ![]() José Birindelli
José Birindelli
Abstract
A new species of Astyanax is described from the rio Jauari and rio Mulato basins, tributaries of the left bank of the lower Amazon River, Pará State, Brazil. The new species is distinguished from the other congeners, except A. kullanderi, A. lineatus, and A. superbus, by presenting chromatophores concentrated on the edges of scales on the sides of the body, forming dark, wavy longitudinal stripes along the flanks. The new species can be diagnosed from A. kullanderi, A. lineatus, and A. superbus by having a greater number of gill rakers in the first branchial arch, body proportions, and color pattern. Among all congeners, the new species is most morphologically similar to A. lineatus. This is the first species of the genus to be described from tributaries of the northern bank of the lower Amazon basin.
Keywords: Astyanax bimaculatus, Astyanax lineatus, Calha Norte Paraense, Taxonomy.
Introduction
The genus Astyanax Baird & Girard, 1854 currently comprises approximately 120 valid species, distributed across freshwater drainages of the Neotropical region, from southern United States to central Argentina (Almirón et al., 2015; Toledo-Piza et al., 2024; Fricke et al., 2025). The non-monophyly of the genus has been consistently demonstrated through multiple lines of evidence, including both morphological and molecular approaches (e.g., Javonillo et al., 2010; Mirande, 2010, 2019; Oliveira et al., 2011; Schmitter-Soto, 2016; Terán et al., 2020).
Terán et al. (2020) combined morphological and molecular data to provide the first comprehensive phylogenetic analysis of the genus recovering it as a polyphyletic group, relocating several species previously attributed to Astyanax to the genera Deuterodon Eigenmann, 1907, Eretmobrycon Fink, 1976, Jupiaba Zanata, 1997, Psalidodon Eigenmann, 1911, Tetragonopterus Cuvier, 1816, and to two newly erected genera, Andromakhe Terán, Benitez & Mirande, 2020, and Makunaima Terán, Benitez & Mirande, 2020. More recently Melo et al. (2024) presented a broad molecular phylogeny of Characidae and related families, further restricting this genus as several species were reassigned to other genera, including 12 species to the genus Psalidodon.
Despite these advances, the diversity of Astyanax remains underestimated (e.g., Melo et al., 2024), particularly in poorly sampled areas of the Amazon basin (e.g., Dagosta, de Pinna, 2019; Dagosta, Marinho, 2022). During recent ichthyological surveys in tributaries of the northern bank of the lower Amazon River in Pará State, Brazil, were collected specimens of an unidentified species of Astyanax very similar to Astyanax lineatus (Perugia, 1891), a species distributed in the La Plata basin in Argentina, Bolivia, Brazil, and Paraguay, and in the upper Río Mamoré basin in Bolivia (e.g., Liotta, 2005; Ferreira et al., 2017; Maldonado et al., 2020).
The so-called Northern Pará Drainage System (sensu Dutra et al., 2020) covers approximately 270,000 km2 and stands out for its uniqueness as it is home to some of the largest Conservation Units (CUs) in the tropics, including the Grão-Pará Ecological Station, the largest in the world, with approximately 4.2 million hectares (IMAZON, 2015). However, the ichthyofauna of this region remains poorly documented, although some fish species have been recently described for the area (e.g., Silva-Oliveira et al., 2018; Dutra et al., 2020; Faria et al., 2020), several additional undescribed species still await formal recognition. In the present study, we formally describe a new species of Astyanax from this poorly explored area, contributing to the knowledge of the biodiversity of the northern Amazon basin.
Material and methods
Counts and measurements follow Fink, Weitzman (1974), except for the number of scale rows below the lateral line, which were counted up to the pelvic-fin origin. Measurements were made point by point on the left side of the body using a digital caliper with an accuracy of 0.1 mm, and are expressed as a percentage of standard length (SL), except for head subunits, which are expressed as a percentage of head length.
Counts of vertebrae, supraneurals, procurrent caudal-fin rays, unbranched anal-fin ray, and minute dentary teeth were taken from specimens prepared according to Taylor, Van Dyke (1985), i.e., cleared and stained (c&s) specimens. Vertebral counts included all four vertebrae in the Weberian apparatus. The compound caudal centrum (PU1+U1) was treated as a single element. A small incision was done at the lateral wall of the abdominal cavity to check the sex and gonadal development of the specimens. In the description, counts are followed by their absolute frequency in parentheses, and holotype counts are indicated by an asterisk. Numbers of gill rakers on the first branchial arch and teeth in the maxilla of Astyanax superbus Myers, 1942, were taken from Dagosta (2011) and Ruiz-C et al. (2018).
In the list of comparative material examined, museum abbreviations and catalogue numbers are followed by the total number of specimens in each lot, standard length range, and collection date. The map was generated using QGis 2.4.0 Chugiak©. Museum abbreviations follow Sabaj (2020).
Results
Astyanax pintacuia, new species
urn:lsid:zoobank.org:act:5889D522-2519-46EE-A21D-391B2E985D6A
(Figs. 1–3; Tab. 1)
Holotype. UFOPA-I 1747, 66.9 mm SL, Brazil, Pará State, Monte Alegre, igarapé Mulato, tributary of rio Mulato, lower Amazon River basin, 01º43’32.11”S 53º59’54.00”W, 4 Nov 2024, J. Sarino-Araújo & C. Silva-Oliveira.

FIGURE 1| Astyanax pintacuia: A. UFOPA-I 1747, holotype, 66.9 mm SL; B. UFOPA-I 1745, paratype, 50.8 mm SL. Brazil, Pará State, Monte Alegre, igarapé Mulato.
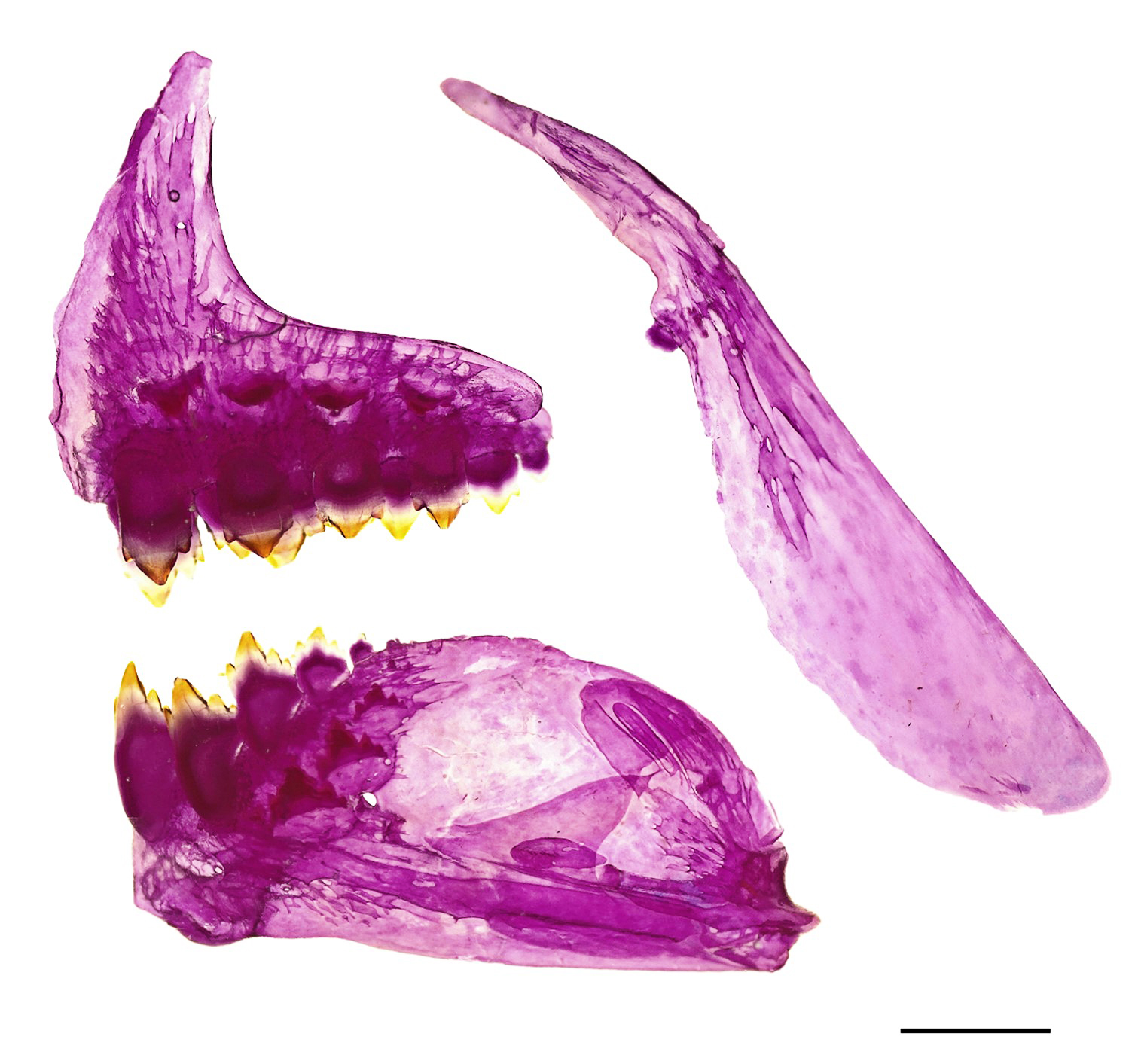
FIGURE 2| Astyanax pintacuia, UFOPA-I 1634, paratype, 46.4 mm SL (c&s). Left side of the premaxillary, maxillary, and dentary. Lateral view. Scale bar = 1 mm.
Paratypes. All from Brazil, Pará State, lower Amazon River basin: INPA 61813, 10, 36.1–43.3 mm SL; UFOPA-I 1745, 58 (4 c&s), 37.2–62.5 mm SL; ZUEC 18560, 10, 41.4–55.1 mm SL, collected with the holotype. UFOPA-I 1634, 8 (1 c&s), 42.0–48.8 mm SL, Monte Alegre, igarapé Água Azul, rio Jauari, 01º20’22”S 53º59’22”W, 11 Aug 2022, C. Silva-Oliveira, M. Lima & S. Silva. UFOPA-I 1636, 2, 40.7–41.5 mm SL, Monte Alegre, rio Jauari, 01º20’28”S 53º56’15”W, 9 Aug 2022, F. R. V. Ribeiro, A. L. C. Canto, J. Sarino-Araújo & S. Silva. UFOPA-I 1637, 3, 51.2–55.0 mm SL, Prainha, cachoeira do Cupim, rio Jauari, 01º22’30”S 53º35’24”W, 12 Aug 2022, C. Silva-Oliveira, M. Lima & S. Silva. UFOPA-I 1644, 1, 72.9 mm SL, Monte Alegre, Igarapé do Lontra, rio Jauari, 01º19’45”S 53º56’49”W, 6 Oct 2023, C. Silva-Oliveira, M. Lima & S. Silva UFOPA-I 1746, 14, 33.9–80.3 mm SL, Monte Alegre, igarapé Mulato, rio Mulato, 01°43’32.11”S 53°59’54.00”W, 7 Jan 2017, C. Silva-Oliveira, A. Canto, F. Ribeiro, A. Silva & J. Souza.
Diagnosis. Astyanax pintacuia differs from its congeners, except A. lineatus, A. kullanderi Costa, 1995 and A. superbus Myers, 1942 by presenting wavy dark stripes between the rows of scales on the sides of the body, forming a well-defined longitudinal striped pattern (vs. absence of a wavy, striped pattern in the remaining congeners). Astyanax pintacuia differs from A. lineatus, A. kullanderi, and A. superbus in the greater number of gill rakers on the first branchial arch (26–28 vs. 19 or 20 in A. kullanderi, 22–24 in A. lineatus and 22 or 23 in A. superbus). Astyanax pintacuia can be diagnosed from A. kullanderi and A. superbus by presenting two vertically elongated humeral blotches (vs. presence of a single, horizontally elongated, oval-shaped humeral blotch). Astyanax pintacuia also differs from A. kullanderi by the number of scales in the lateral line (37–40 vs. 31–32). Additionally, the new species differs from A. superbus by presenting a short maxilla, not reaching the vertical that passes through the anterior margin of the pupil (vs. maxilla long, exceeding the vertical that passes through the center of the pupil), and by the presence of one tricuspid tooth in the maxilla (vs. 2–6 tricuspid teeth). Finally, it additionally differs from A. lineatus by presenting the longitudinal wavy stripes only present along the midbody (vs. longitudinal wavy stripes present both along the midbody and in the dorsal portion of the body).
Description. Morphometric data presented in Tab. 1. Body compressed, greatest body height at vertical through dorsal-fin origin. Dorsal profile of head slightly convex from upper lip to vertical through the posterior nostril, straight from that point to tip of supraoccipital spine. Body profile slightly convex from the distal end of supraoccipital spine to dorsal-fin terminus, straight to slightly convex from that point to adipose fin insertion; slightly concave between the insertion of adipose fin and the origin of the dorsal procurrent caudal–fin rays. Ventral profile of head and body convex from tip of lower jaw to anal-fin origin. Ventral profile straight and posterodorsally inclined along anal-fin base. Ventral profile of caudal peduncle concave.
TABLE 1 | Morphometric data of Astyanax pintacuia (N = 41; holotype included in the range and mean). SD = Standard deviation.
| Holotype | Range | Mean | SD |
Standard length (mm) | 66.9 | 32.9–77.5 | 47.6 | – |
Percents of standard length | ||||
Predorsal distance | 50.6 | 50.1–56.1 | 53.0 | 1.4 |
Prepelvic distance | 49.5 | 47.6–52.0 | 49.6 | 1.3 |
Prepectoral distance | 27.1 | 27.1–33.1 | 29.7 | 1.3 |
Preanal distance | 68.4 | 64.0–69.5 | 67.2 | 1.3 |
Depth at dorsal-fin origin | 40.1 | 36.1–41.3 | 38.7 | 1.5 |
Caudal peduncle depth | 11.8 | 10.7–12.1 | 11.4 | 0.4 |
Caudal peduncle length | 8.5 | 5.9–11.0 | 8.0 | 1.1 |
Anal-fin base length | 29.7 | 26.8–33.7 | 30.8 | 1.3 |
Dorsal-fin base length | 15.4 | 13.6–17.2 | 15.3 | 0.7 |
Dorsal-fin length | 31.4 | 28.1–34.1 | 30.8 | 1.1 |
Pelvic-fin length | 18.3 | 15.7–22.0 | 18.1 | 1.2 |
Anal fin length | 20.0 | 15.0–22.5 | 20.2 | 1.3 |
Pectoral-fin length | 22.4 | 21.7–25.5 | 23.5 | 1.0 |
Head length | 28.2 | 28.2–33.5 | 30.8 | 1.2 |
Percents of head length | ||||
Snout length | 19.2 | 17.9–23.0 | 20.0 | 1.1 |
Upper jaw length | 37.3 | 35.9–40.5 | 37.7 | 1.0 |
Orbital diameter | 37.6 | 35.7–43.5 | 40.5 | 2.0 |
Interorbital width | 31.3 | 26.2–33.8 | 30.6 | 1.4 |
Mouth terminal; jaws isognathous. Posterior terminus of maxilla reaching vertical through anterior margin of orbit. Premaxilla with two teeth rows; outer row with 4(5) tricuspid teeth; inner row with 5(5) tricuspid to pentacuspid teeth. Maxilla with 1(5) tricuspid or pentacuspid tooth. Dentary with 5(2) or 6(3) tricuspid to pentacuspid teeth followed by 4 small conical teeth.
Scales cycloid, moderately large, with 4 to 7 well-marked radii; circuli present in the anterior and posterior fields of scales. Lateral line complete, slightly curved, with 37(10), 38*(11), 39(13) or 40(6) perforated scales. Longitudinal series of scales between dorsal-fin origin and lateral line, 7*(32) or 8(9). Longitudinal series of scales between lateral line and pelvic-fin origin, 6*(41). Predorsal scales 11(13), 12(22) or 13*(6), arranged in regular series. Circumpeduncular scales 16(9) or 17*(31). Single series of 20*(29), 21(8), 22(2), 23(1) or 24(1) scales covering base of anal fin.
Dorsal-fin rays ii,9*(41). Dorsal-fin origin at midbody, slightly posterior to vertical through pelvic fin origin. First unbranched ray approximately half length of second unbranched ray. Adipose fin present, origin approximately vertical through base of 23rd to 24th branched anal-fin rays. Pectoral-fin rays i,10(9), i,11*(27), i,12(5); tip of pectoral fin reaching pelvic-fin origin when compressed. Pelvic-fin rays i,7*(41); tip of pelvic fin extending beyond urogenital papillae and usually reaching first anal-fin rays when compressed. Anal-fin origin located vertically through posterior terminus of dorsal-fin base. Anal-fin rays iv, 24(2), 25(7), 26*(14), 27(12), 28(3), or 29(3). Last unbranched and first three branched anal-fin rays longest, subsequent rays gradually decreasing in length. Caudal fin forked, lobes equal in size, slightly rounded. Principal caudal-fin rays 10+9*(41); dorsal procurrent caudal-fin rays 11(4) or 12(1); ventral procurrent caudal-fin rays 9(2) or 10(3).
Total vertebrae 36(3) or 37(2). Supraneurals 4(3) or 5(2), with bony lamellae in the upper portion. Branchiostegal rays 4(5). First branchial arch with 26(29), 27(11) or 28(1) rakers: 3(40) or 4(1) on hypobranchial, 11(15), 12(25) or 13(1) on ceratobranchial, 1(41) on cartilage between the ceratobranchial and epibranchial and 9(1), 10(14), 11(25) or 12(1) on epibranchial.
Coloration in alcohol. Overall coloration of body brown to silver. Top of head and snout dark as a result of dense concentration of dark chromatophores. Infraorbitals, maxillary, ventral region of opercle, and gular region silvery to yellowish. Dorsolateral region of body darkened. Scales of latero-dorsal region of body with chromatophores concentrated. Humeral region with two vertically elongated blotches; first humeral blotch conspicuous, extending vertically over five to six rows of scales, extending beyond lateral line ventrally; upper portion broader and more evident than lower portion. Second humeral blotch diffuse, extending horizontally over two to three rows of scales and vertically over four rows of scales, not extending beyond lateral line ventrally. Darker diamond-shaped blotch on caudal peduncle, reaching to end of middle caudal fin rays. Faint, wide, longitudinal dark to silvery band extending posteriorly to second humeral blotch to caudal peduncle blotch. Six to eight wavy dark longitudinal stripes along middle portion of body. Abdominal region clear, with few scattered dark chromatophores. Dorsal, adipose, pectoral, pelvic and anal fins hyaline, with few dark chromatophores on interradial membranes. Median caudal-fin rays densely pigmented, forming a black band.
Coloration in life. Overall body coloration mainly silvery. Dorsal portion of the head and dorsal region of body light gray to yellowish. Lower portion of head and abdominal region silvery. Dorsal portion of orbit yellow to orange, lateral and ventral portions silvery. Dorsal, adipose, caudal, anal and pelvic fins yellowish with a reddish hue at fin tips. Pectoral fin hyaline, with few scattered chromatophores (Fig. 3).

FIGURE 3| Astyanax pintacuia, UFOPA-I 1745, paratype, 45.1 mm SL, in life. Lateral view, left side.
Sexual dimorphism. Observed in three mature males (UFOPA-I 1746, 55.6–62.1 mm SL). Mature males have 11 thin, retrorse hooks on the largest unbranched anal-fin ray and 13 hooks on the first branched ray. Number of hooks gradually decreasing to five hooks on the 11th or 12th branched anal-fin rays. Three first branched pelvic-fin rays with six to nine slightly retrorse hooks, with a more robust base and smaller than those present in the anal fin. One hook per segment of anal and pelvic-fin rays. No other secondary sexually dimorphic features were identified.
Geographical distribution. Astyanax pintacuia is known from tributaries of rio Jauari and rio Mulato, lower Amazon River basin, Pará State, Brazil (Fig. 4).
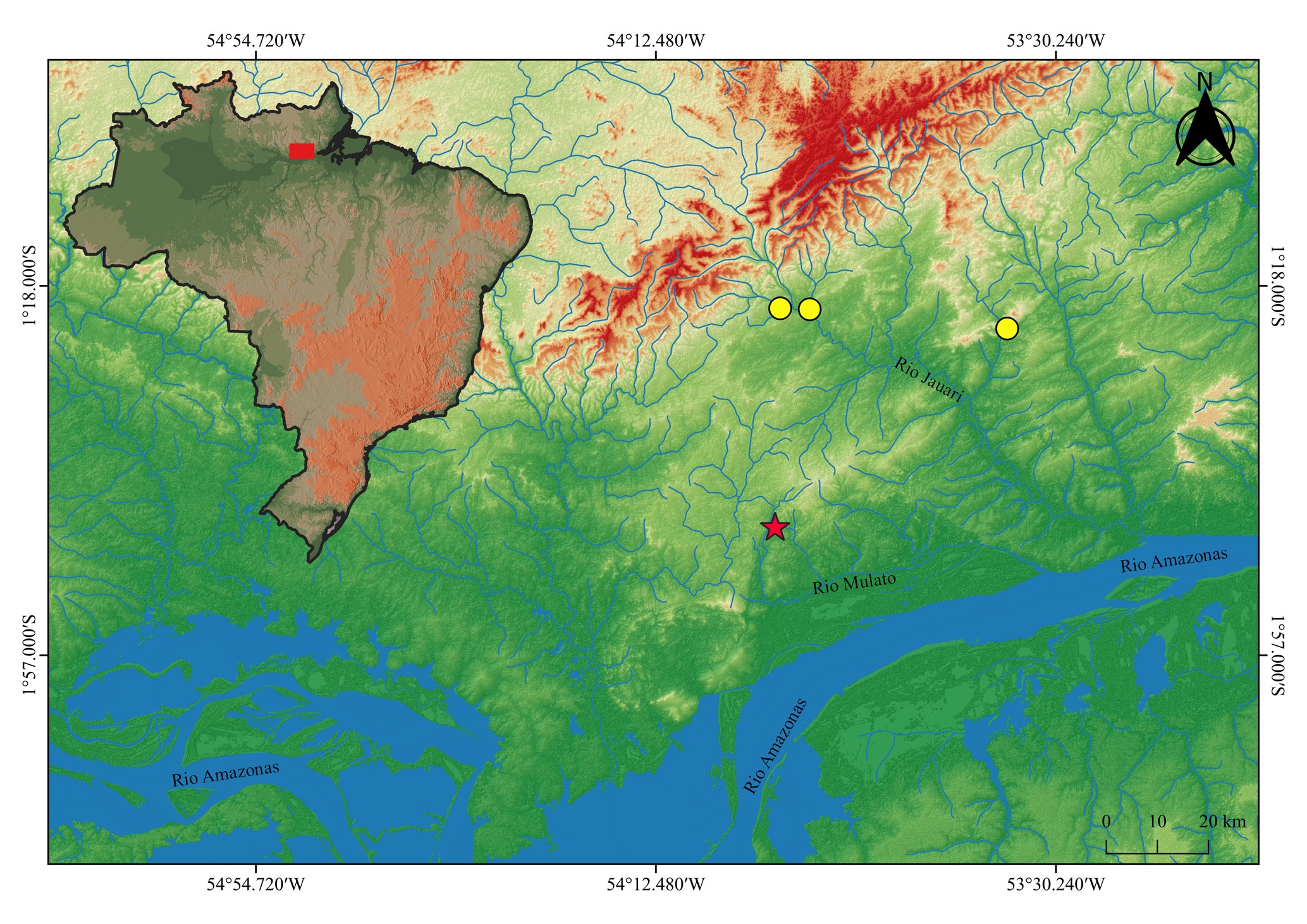
FIGURE 4| Distribution of Astyanax pintacuia (yellow circles). The red star indicates the type locality.
Conservation status. Astyanax pintacuia presents a wide Extent of Occurrence (EOO), and additionally, is common and abundant across its range. Furthermore, no threats were identified in the sites of occurrence of the species. Thus, according to the International Union for Conservation of Nature categories and criteria (IUCN, 2024), we suggest that A. pintacuia should be classified as Least Concern (LC).
Etymology. The specific epithet “pintacuia” is a tribute to the pinta-cuia (painted gourds) culture, a local custom inherited from the indigenous peoples of the Gurupatuba settlement, a term later affectionately extended to the natives of the municipality of Monte Alegre (Martins, 2015; Magnani, 2023). A noun in apposition.
Discussion
In a phylogenetic hypothesis of Astyanax and related genera based on combined morphological and DNA data, Terán et al. (2020) recovered a clade composed of A. lineatus, A. metae Eigenmann, 1914, A. pirapuan Tagliacollo, Britzke, Silva & Benine, 2011, and A. venezuelae Schultz, 1944. This monophyletic group, called A. lineatus clade by Téran et al. (2020), was supported by ten molecular synapomorphies and a single morphological synapomorphy, namely “scale sheath covering three-quarters or more of the base of the anal fin”. However, among the four species, only A. lineatus was analyzed using both morphological and molecular data, while the other species were included based solely on molecular evidence.
More recently, Melo et al. (2024), based on ultraconserved elements, obtained a close relationship among the species of the clade. Although A. lineatus was not included in their analysis, the remaining species of the A. lineatus clade were recovered in close phylogenetic proximity, partially corroborating the hypothesis of Terán et al. (2020). However, in Melo et al. (2024), this clade forms the sister group of all remaining Astyanax species, whereas in Téran et al. (2020), it was recovered as the sister group of a more inclusive clade, that includes Gymnocharacinus Steindachner, 1903, Astyanax sensu stricto, Andromakhe, Oligosarcus Günther, 1864, and Psalidodon.
The only morphological synapomorphy proposed for the Astyanax lineatus clade, i.e., the scale sheath extending over at least ¾ of the anal-fin base, is present in A. pintacuia, which exhibits a scale sheath covering approximately 80% of the anal-fin base. However, because this character is highly homoplastic and occurs in multiple taxa closely related to the A. lineatus clade, including several Astyanax species, the presence of this feature alone is insufficient to confidently place A. pintacuia within the A. lineatus clade.
Among all species currently included in Astyanax, A. pintacuia is morphologically most similar to A. lineatus (Fig. 5). Both species share the presence of two vertically elongated humeral blotches and longitudinal wavy stripes along the sides of the body. Furthermore, A. pintacuia shares with the species of the A. lineatus clade the predorsal scales arranged in a regular series. These morphological similarities suggest that A. pintacuia is more closely related to the members of this clade, although a study with a molecular approach is necessary to confirm the placement of the new species within Astyanax.

FIGURE 5| Astyanax lineatus: A. ZUEC 6811, 74.7 mm SL, Brazil, Mato Grosso, Chapada dos Guimarães, rio Claro; B. ZUEC 18550, 69.1 mm SL, Brazil, Mato Grosso, Nortelândia, rio Santana.
As mentioned in the Introduction, the diversity of Astyanax is still underestimated in the Northern Pará drainage system. Notably, a second putative species exhibiting a color pattern strikingly similar to that of A. pintacuia was recently collected from the same general region (ZUEC 18346). Despite the similar coloration, this morphotype presents some differences in body proportions, including body depth (30.3–35.4% vs. 36.1–41.3% SL in A. pintacuia) and predorsal length (49.2–52.7% vs. 50.1–56.1% SL in A. pintacuia). The two species were recorded occurring syntopically in the upper portion of the rio Jauari, while only A. pintacuia was recorded from the rio Mulato. Furthermore, at least three additional putative species belonging to the A. bimaculatus species complex have been recorded in the Northern Pará drainage system (JSA, 2025, pers. obs.). An integrative taxonomic approach of Astyanax species inhabiting the rivers of northern Pará is currently being conducted by JSA to assess the true diversity of the genus in the region.
Comparative material examined. Brazil: Astyanax cf. argyrimarginatus Garutti, 1999: UFOPA-I 1638, 1, 97.8 mm SL. UFOPA-I 1640, 1, 63.9 mm SL. UFOPA-I 1645, 2, 63.1–86.0 mm SL. Astyanax cf. bimaculatus (Linnaeus, 1758): UFOPA-I 1635, 8, 45.3–95.9 mm SL. Astyanax gr. bimaculatus: UFOPA-I 867, 2, 53.4–65.2 mm SL. Astyanax sp.: ZUEC 18346, 5, 42.6–49.2 mm SL. Astyanax sp. 1: UFOPA-I 1641, 27, 38.8–88.1 mm SL. Astyanax sp. 2: UFOPA-I 1576, 33, 42.6–81.1 mm SL. Astyanax kullanderi:ZUEC 13447, 3, 50.5–54.5 mm SL. ZUEC 13452, 1 of 2, 48.7 mm SL. Astyanax lineatus: LBP 35852, 16, 33.1–50.1 mm SL. MZUSP 90013, 4, 59.3–65.4 mm SL. MZUSP 91118, 8, 64.3–82.2 mm SL. ZUEC 4188, 2, 72.0–74.6 mm SL. ZUEC 6811, 11, 51.12–100.3 mm SL. ZUEC 18505, 6, 43.5–71.2 mm SL. ZUEC 10569, 1, 50.1 mm SL. ZUEC 18550, 14, 63.2–83.1 mm SL. Astyanax pirapuan: LBP 17539, 16, 16.3–41.4 mm SL. ZUEC 7634, 8, 31.0–76.6 mm SL. Venezuela: Astyanax metae: LBP 18700, 5, 58.3–71.3 mm SL. Astyanax venezuelae: LBP 18701, 5, 48.3–64.5 mm SL.
Acknowledgments
The authors are grateful to Alberto Silva, Sinara Silva and Marcos Lima for help during field trips. We also thank Gabriel Deprá and Cláudio Oliveira (LBP), Murilo Pastana, Michel Gianeti, Alessio Datovo, Osvaldo Oyakawa, and Mario de Pinna (MZUSP) for allowing the examination of specimens under their care.
References
Almirón AE, Casciotta JRC, Ciotek L, Giorgis P. Guía de los peces del Parque Nacional Pre-Delta. Ciudad autónoma de Buenos Aires, Argentina: Administración de Parques Nacionales; 2015. Available from: https://sib.gob.ar/archivos/bfa004429.pdf
Dagosta FCP. Taxonomia e relações filogenéticas do gênero Astyanacinus Eigenmann, 1907 (Characiformes: Characidae). [Master Dissertation]. Ribeirão Preto: Universidade de São Paulo; 2011.
Dagosta FCP, Marinho MMF. New small-sized species of Astyanax (Characiformes: Characidae) from the upper rio Paraguai basin, Brazil, with discussion on its generic allocation. Neotrop Ichthyol. 2022; 20(1):e210127. https://doi.org/10.1590/1982-0224-2021-0127
Dagosta FCP, de Pinna M. The fishes of the Amazon: distribution and biogeographical patterns, with a comprehensive list of species. Bull Am Mus Nat Hist. 2019; 2019(431):1–163. https://doi.org/10.1206/0003-0090.431.1.1
Dutra GM, Freitas TMS, Prudente BS, Salvador GN, Leão MDV, Peixoto LAW et al. Rapid assessment of the ichthyofauna of the southern guiana shield tributaries of the amazonas river in Pará, Brazil. Acta Amaz. 2020; 50(1):24–36. https://doi.org/10.1590/1809-4392201901402
Faria TC, Lima FCT, Wosiacki WB. A new Hyphessobrycon (Characiformes: Characidae) from the Guiana Shield in Northern Brazil. Copeia. 2020; 108(2):369–75. https://doi.org/10.1643/CI-19-311
Ferreira FS, Vicentin W, Súarez YR. Astyanax lineatus (Perugia, 1891) (Characiformes: Characidae): first record in the upper Paraná river basin, Mato Grosso do Sul, Brazil. Check List. 2017; 13(2):1–04. https://doi.org/10.15560/13.2.2094
Fink WL, Weitzman SH. The so-called cheirodontin fishes of Central America with descriptions of two new species (Pisces: Characidae). Smithson Contrib Zool. 1974(172):1–46. https://doi.org/10.5479/SI.00810282.172
Fricke R, Eschmeyer WN, Van der Laan R. Eschmeyer’s catalog of fishes: genera, species, references [Internet]. San Francisco: California Academy of Natural Sciences; 2025. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Instituto do Homem e Meio Ambiente da Amazônia (IMAZON). Calha Norte Sustentável: situação atual e perspectivas. [Internet]. 2015. Available from: https://imazon.org.br/calha-norte-sustentavel-situacao-atual-e-perspectivas/
International Union for Conservation of Nature (IUCN). Standards and petitions committee. Guidelines for using the IUCN Red List categories and criteria. Version 16 [Internet]. Gland; 2024. Available from: http://www.iucnredlist.org/documents/RedListGuidelines.pdf
Javonillo R, Malabarba LR, Weitzman SH, Burns JR. Relationships among major lineages of characid fishes (Teleostei: Ostariophysi: Characiformes), based on molecular sequence data. Mol Phylogenet Evol. 2010; 54(2):498–511. https://doi.org/10.1016/J.YMPEV.2009.08.026
Liotta JR. Distribución geográfica de los peces de aguas continentales de la República Argentina. ProBiota Ser Doc. 2005; 3.
Maldonado M, Barra E, Arraya M, Zubieta J, Machicao F, Carvajal-Vallejos FM et al. Patrones de distribución, composición y riqueza de la ictiofauna en los Andes de Bolivia. Neotrop Hydrobiol Aquat Conserv. 2020; 1(2):139–60.
Magnani AOM, Ferreira SLM. Mulheres do Brasil: artes e artistas. Sevilla: Enredars/Universidad Pablo de Olavide, Roma, Roma TrE-Press/Università degli studi Roma Tre; 2023.
Martins RMA. Além do olhar: as fontes sobre a apropriação das técnicas e dos materiais das culturas indígenas nas artes da Amazônia Colonial (séculos XVII e XVIII). In: Knauss P, Malta M, editors. Objetos do olhar: história e arte. São Paulo: Rafael Copetti Editor; 2015. p.139–54.
Melo BF, Ota RP, Benine RC, Carvalho FR, Lima FCT, Mattox GMT et al. Phylogenomics of Characidae, a hyper-diverse Neotropical freshwater fish lineage, with a phylogenetic classification including four families (Teleostei: Characiformes). Zool J Linn Soc. 2024; 202(1):1–37. https://doi.org/10.1093/zoolinnean/zlae101
Mirande JM. Phylogeny of the family Characidae (Teleostei: Characiformes): from characters to taxonomy. Neotrop Ichthyol. 2010; 8(3):385–568. https://doi.org/10.1590/S1679-62252010000300001
Mirande JM. Morphology, molecules and the phylogeny of Characidae (Teleostei, Characiformes). Cladistics. 2019; 35(3):282–300. https://doi.org/10.1111/cla.12345
Oliveira C, Avelino GS, Abe KT, Mariguela TC, Benine RC, Ortí G et al. Phylogenetic relationships within the speciose family Characidae (Teleostei: Ostariophysi: Characiformes) based on multilocus analysis and extensive ingroup sampling. BMC Evol Biol. 2011; 11(1):275. https://doi.org/10.1186/1471-2148-11-275
Ruiz-C. RI, Román-Valencia C, Taphorn DC, Buckup PA, Ortega H. Revision of the Astyanax orthodus species-group (Teleostei: Characidae) with descriptions of three new species. Eur J Taxon. 2018; 2018(402). https://doi.org/10.5852/EJT.2018.402
Sabaj MH. Codes for natural history collections in ichthyology and herpetology. Copeia. 2020; 108(3):593–669. https://doi.org/10.1643/ASIHCODONS2020
Schmitter-Soto JJ. A phylogeny of Astyanax (Characiformes: Characidae) in Central and North America. Zootaxa. 2016; 4109(2):101–30. https://doi.org/10.11646/ZOOTAXA.4109.2.1
Silva-Oliveira C, Lima FCT, Bogotá-Gregory JD. A new species of Bryconops Kner (Characiformes: Iguanodectidae) from the rio Maicuru, lower Amazon basin, Brazil. Zootaxa. 2018; 4514(3):372–82. https://doi.org/10.11646/ZOOTAXA.4514.3.4
Taylor WR, Van Dyke GC. Revised procedure for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium. 1985; 9(2):107–19.
Terán GE, Benitez MF, Mirande JM. Opening the Trojan horse: phylogeny of Astyanax, two new genera and resurrection of Psalidodon (Teleostei: Characidae). Zool J Linn Soc. 2020; 190(4):1217–34. https://doi.org/10.1093/zoolinnean/zlaa019
Toledo-Piza M, Baena E, Dagosta FCP, Menezes NA, Ândrade M, Benine RC et al. Checklist of the species of the order Characiformes (Teleostei: Ostariophysi). Neotrop Ichthyol. 2024; 22(1):e230086. https://doi.org/10.1590/1982-0224-2023-0086
Authors

![]() Josué Sarino-Araújo1
Josué Sarino-Araújo1 ![]() ,
, ![]() Cárlison Silva-Oliveira2,
Cárlison Silva-Oliveira2, ![]() Flávio C. T. Lima3,
Flávio C. T. Lima3, ![]() André L. C. Canto4 and
André L. C. Canto4 and ![]() Frank R. V. Ribeiro1,4
Frank R. V. Ribeiro1,4
[1] Programa de Pós-Graduação em Biodiversidade, Universidade Federal do Oeste do Pará, Rua Vera Paz, s/n, 68040-255, Santarém, PA, Brazil. (JSA) josuesarinoaraujo@gmail.com (corresponding author), (FRVR) fraynner@yahoo.com.br.
[2] Instituto Federal de Educação, Ciência e Tecnologia do Pará – Campus Itaituba, Rua Universitário, s/n, 68183-300, Itaituba, PA, Brazil. (CSO) carlison3@gmail.com.
[3] Museu de Diversidade Biológica, Instituto de Biologia, Universidade Estadual de Campinas, Rua Monteiro Lobato, 255, 13083 682, Campinas, SP, Brazil. (FCTL) fctlima@gmail.com.
[4] Instituto de Ciências e Tecnologia das Águas, Universidade Federal do Oeste do Pará, Rua Vera Paz, s/n, 68040-255, Santarém, PA, Brazil. (ALCC) cantoandre@gmail.com.
Authors’ Contribution 

Josué Sarino-Araújo: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Writing-original draft, Writing-review and editing.
Cárlison Silva-Oliveira: Conceptualization, Investigation, Methodology, Supervision, Writing-original draft, Writing-review and editing.
Flávio C. T. Lima: Data curation, Formal analysis, Investigation, Supervision, Writing-original draft, Writing-review and editing.
André L. C. Canto: Data curation, Methodology, Project administration, Supervision, Writing-original draft, Writing-review and editing.
Frank R. V. Ribeiro: Project administration, Supervision, Writing-original draft, Writing-review and editing.
Ethical Statement
Collection Licenses of SISBIO number 10227–14.
Competing Interests
The author declares no competing interests.
Data availability statement
The authors confirm that the data supporting the findings of this study are available within the article.
AI statement
The author declares no competing interests.
Funding
Support for the project was provided by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, process 88887.199140/2025–00) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, process 436763/2018–4). CSO and FRVR are funded by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq process number 317781/2021–9 and 310480/2022–1, respectively). ALCC and FRVR were partly supported by Instituto de Conservação Ambiental The Nature Conservancy do Brasil (Águas Tapajós Project, Cooperation BR FY23 264).
Peer Review
How to cite this article
Sarino-Araújo J, Silva-Oliveira C, Lima FCT, Canto ALC, Ribeiro FRV. A new tetra of the genus Astyanax (Characiformes: Acestrorhamphidae) with lateral stripes from lower Amazon basin, Brazil. Neotrop Ichthyol. 2026; 24(2):e250154. https://doi.org/10.1590/1982-0224-2025-0154
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2025 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted February 11, 2026
Accepted February 11, 2026
![]() Submitted September 1, 2025
Submitted September 1, 2025
![]() Epub June 26, 2026
Epub June 26, 2026