![]() Matías Briones1,
Matías Briones1, ![]() Jorge Cortés-Miranda1,
Jorge Cortés-Miranda1, ![]() Claudio Veloso1,
Claudio Veloso1, ![]() David Véliz1,2 and
David Véliz1,2 and ![]() Caren Vega-Retter1
Caren Vega-Retter1 ![]()
PDF: Download Here | Supplementary: Sup | Cite this article
Associate Editor: ![]() Fernando Pelicice
Fernando Pelicice
Section Editor: ![]() Fernando Pelicice
Fernando Pelicice
Editor-in-chief: ![]() José Birindelli
José Birindelli
Abstract
La contaminación en ecosistemas de agua dulce degrada la calidad del agua y altera las comunidades de macroinvertebrados, afectando su valor como recurso alimentario. En cuencas agrícolas, como el río Maipo en Chile, la canalización es una perturbación adicional. Mediante isótopos estables (δ¹5N y δ¹³C), este estudio evaluó cómo la contaminación y el curso de agua influyen en la cantidad y calidad del alimento disponible para el pez Basilichthys microlepidotus. Se midieron riqueza, abundancia y valor nutricional de los macroinvertebrados, contenido estomacal, preferencias alimentarias, las señales isotópicas y posición trófica del pez. Se analizaron tres sitios contaminados y uno no contaminado (dos naturales y dos artificiales). Los sitios contaminados presentaron menor diversidad de macroinvertebrados y menor contenido calórico y lipídico. Los canales mostraron menor calidad del alimento, pero una distribución más diversa que los cauces naturales. Sin embargo, contaminación y curso de agua no afectaron la condición corporal de los peces detectándose selección alimentaria por gastrópodos Physidae. Altos valores de δ¹⁵N y señales variables de δ¹³C en áreas contaminadas indican cambios dietarios, aunque la posición trófica se mantuvo estable. En conjunto, los impactos antrópicos influyen en la calidad y composición de presas, siendo vital considerarlos en la gestión de los ecosistemas dulceacuícolas.
Palabras clave: Canalización, Contaminación del agua, Dieta del pez, Invertebrados bentónicos, Isotopos estables.
Introduction
Aquatic ecosystems are among the most impacted in the world, with pollution being one of the main causes of water quality degradation (World Water Assessment Programme, 2017). Pollution negatively affects freshwater biota by increasing mortality, decreasing fertility, and changing the structure and function of the community (Cooper, 1993; Smith et al., 1999). Pollution stands as one of the most pervasive threats to freshwater biodiversity. Among threatened freshwater species, 54% are affected by pollution, making it the most prevalent single threat. Pollution originates from multiple sources, with agricultural and forestry effluents affecting 45% of threatened fish species, domestic and urban wastewaters impacting 29–33%, and industrial and military effluents affecting 21% (Sayer et al., 2025).
In freshwater ecosystems, macroinvertebrates play a fundamental role in the trophic structure (Fu et al., 2016). Macroinvertebrates are key to the recycling of organic matter and the transfer of energy from organic matter and primary producers to organisms at higher trophic levels, such as fishes (Philips, 2004; Fu et al., 2016). The diversity and abundance of macroinvertebrates in an ecosystem can be influenced by a variety of environmental factors, such as habitat characteristics (Fierro et al., 2015), water quality (Figueroa et al., 2003), and seasonality (Tupinambás et al., 2007), as well as biological factors like competition and predation (Kohler, 1992). Low-altered rivers typically host more sensitive taxa, like Ephemeroptera, Megaloptera, Plecoptera, Odonata, and Trichoptera, that usually require well-oxygenated, flowing waters (Simaika, Samways, 2011). In contrast, polluted rivers show a shift in macroinvertebrate communities, with sensitive species declining and more tolerant taxa, such as nematodes, annelids, gastropods, and chironomids larvae, increasing (Theodoropoulos et al., 2015; Fierro et al., 2017).
Shifts in macroinvertebrate diversity in response to pollution can affect organisms at higher trophic levels; for example, Scharnweber et al. (2024) observed that benthic macroinvertebrate abundance was significantly reduced at sites contaminated with iron oxides from mining in the Spree River, Germany. These authors found that the abundance and condition of the native piscivorous pike (Esox lucius) were significantly lower at polluted sites compared to remediated sites, suggesting reduced food availability due to macroinvertebrate depletion. Moreover, it has been suggested that pollution can reduce the quality of macroinvertebrates as food. A study by Kokotović et al. (2024) detected lower total lipid content in caddisflies and mayfly exposed to increased temperature and wastewater. Additionally, some studies show that certain pollutants can cause an increase in lipid peroxidation, such as in mollusks exposed to heavy metals (Belcheva et al., 2015) and crustaceans exposed to dissolved organic matter (Timofeyev et al., 2006).
Pollution can also modify the trophic structure of freshwater ecosystems (Fry, 2008). As a complement to stomach contents analyses, which has been widely used to study the trophic ecology of the organism, (Tripp-Valdez, Arreguin-Sanchez, 2009) stable isotope analyses has been proposed as a useful tool providing information about nutrient sources, and how these nutrients are transferred through food chains (Steffy, Kilham, 2004; Brito et al., 2006; Smucker et al., 2018). Land use around rivers and sewage pollution are another key factor in the isotopic load of the environment and the food web of an ecosystem (Lee et al., 2018; Rau et al., 1981). In addition to environmental load and diet, nutritional stress (starvation) can also enrich the tissues of organisms in 15N (Hertz et al., 2015).
Central Chile’s rivers and lakes face intense anthropogenic pressure (Habit et al., 2024), with the Maipo River heavily affected by urban and agricultural pollution, irrigation, and recreation (DGA, 2004). Agricultural activity in central Chile consumes high volumes of water, making water channelization for irrigation a common practice (Habit et al., 2005). This creates strong environmental disturbances for most of the riverine fish (Redding, Midlen, 1992) altering fish assemblages (Travnichek, Maceina, 1994). Nonetheless, Habit et al. (2005) observed that irrigation channels can serve as temporary habitats, notably for Trichomycterus areolatus which completes its life cycle there. This success may stem from similar macroinvertebrate availability between the irrigation channels and the main river course. Koetsier, McCauley (2015) reported that the macroinvertebrate community in the channel did follow community assembly patterns of small streams reported in the literature, and it also was similar to the community of a nearby natural stream. However, the study of the macroinvertebrate availability and its quality between the irrigation channels and the main river course continues to be little studied.
In the Maipo River, Basilichthys microlepidotus (Jenyns, 1841) has declined notably (Muñoz Donoso, 2007). Distributed between 28°–39° S, it feeds on insect larvae and small macroinvertebrates (Véliz et al., 2012) reproducing from August to January (Comte, Vila, 1992). Microsatellite data revealed five genetically distinct populations in the river, with two in polluted and three in less-polluted areas, inhabiting both main natural channels (central) and artificial irrigation channels (channel) (Vega-Retter et al., 2014). Moreover, Vega-Retter et al. (2018) performed gene expression analyses and found differences in genes related to gluconeogenesis and proteolysis between individuals from polluted and less-polluted sites, suggesting possible starvation due to reduced food quality or availability. Therefore, this study aimed to assess the effects of pollution and watercourse type (central and channel) on the food resources relevant for B. microlepidotus by assessing the richness and abundance of the macroinvertebrate community and the quality of macroinvertebrates as a food source (in terms of energy value and lipid content). We also evaluate the stomach content, the stable isotope of nitrogen (δ¹⁵N) and carbon (δ¹3C) and trophic position of B. microlepidotus across different study sites in the Maipo River. We hypothesize that pollution and water channelization affect negatively the availability and quality of food resources (macroinvertebrates), with effects in the diet, body condition and trophic position of B. microlepidotus.
Material and methods
Sampling sites and physicochemical characterization. Four sites within the Maipo River basin that contain genetically independent populations of the silverside Basilichthys microlepidotus were analyzed. The Maipo river runs from the Andean mountains to the Pacif ocean. Due to the high-altitude origin of the Maipo river streams (around 4,500 m a.s.l.) and the river’s short extension (100 km), the sampling sites generally contained rocky sediment (Vila et al., 2006; Farías et al., 2008). San Francisco de Mostazal (SFM), present less population (around 13,000 inhabitants), no industries in the main city, and in general far less industries and other economic activities if compared to other studied sites. On the contrary Pelvin (PEL) site is in the Peñaflor municipality (around 90,000 inhabitants) and it is downstream of Santiago city, the capital of Chile, and near to the Mapocho river which receive the wastewater discharges of treatment plants. In this same line, Melipilla (MEL) is placed near Melipilla city (around 150,000 inhabitants), which presents higher extraction of aggregate (mining sand, gravel, and crushed stone from natural sources) and agricultural activities and is also downstream from Santiago city. Near both sites, MEL and PEL, the presence of polycyclic aromatic hydrocarbons (PAHs) and organic and nano pollutants have been recorded (Wilkinson et al., 2022; Soriano et al., 2024). In the case of Isla de Maipo (IM) it is placed in Isla de Maipo municipality (around 36,000 inhabitants) which is characterized by a higher number of agricultural activities.
The physicochemical properties of these sites were previously characterized using principal component analysis and the “British Columbia Water Quality” index by Vega-Retter et al. (2014) but were analyzed again through resampling in 2016, the same year and season that the present study was conducted and a clear separation of the non-polluted site (SFM) from the polluted sites (MEL, IM, PEL) is observed. Moreover, a segregation of MEL from the other two polluted sites is also detected, indicating that the pollution profile is not the same in all the study sites (Cortés-Miranda et al., 2024). Additionally, Cortés-Miranda et al. (2024) analyzed stored physical and chemical data from 2007 to 2016, demonstrating that the sampling sites showed a consistent pollution profile, with the MEL, PEL, and IM sites being more polluted, and the San Francisco de Mostazal (SFM) site less polluted. We have provided the water quality variables recorded in Tab. S1, but for a more detailed analysis check Cortés-Miranda et al. (2024).
Taking into consideration all the available information, we classified the SFM site as less polluted (non-polluted here for simplicity), and the MEL, IM, and PEL sites as polluted (Fig. 1). Additionally, we classified the sites according to watercourse type as either central-channel sites (MEL and SFM) or channel sites (IM and PEL). The MEL site, a central-channel site, exhibits a concrete structure modifying part of the riverbed. At SFM, the other central-channel site, the only evident anthropogenic activity are roads that cross the river. Among the channel sites, only PEL has a barrier to regulate flow, whereas IM is an irrigation channel used for crop production in the surrounding area. All sites have riparian vegetation, except for MEL. We were unable to sample a non-polluted channel; although IM was previously considered non-polluted, its physical and chemical characteristics changed in 2016, showing a profile consistent with a polluted site (Cortés-Miranda et al., 2024).
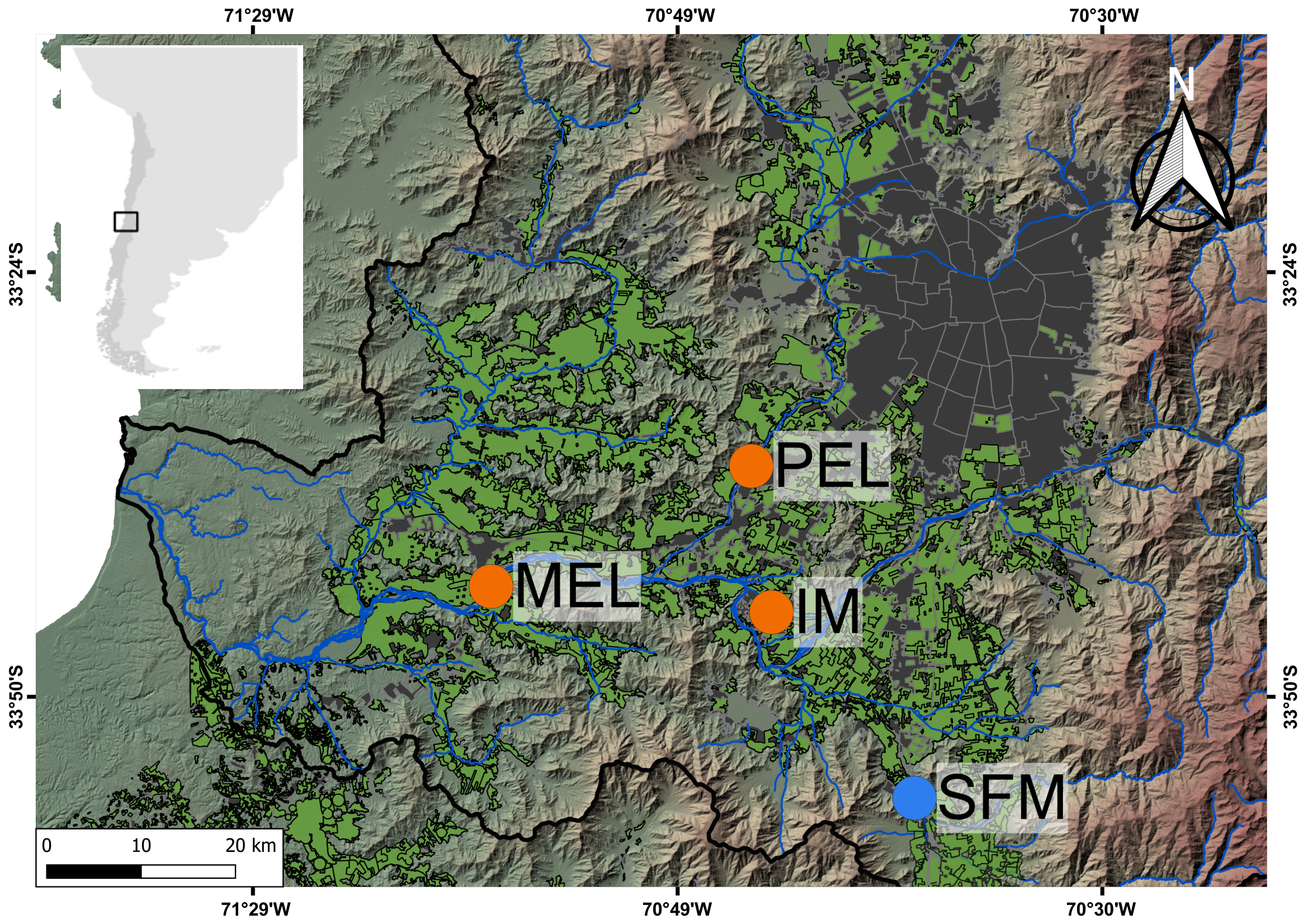
FIGURE 1| Map of the sampling sites in the Maipo river basin; the polluted sites are shown in orange, and the non-polluted site is shown in blue, according to previous characterizations. Melipilla (MEL); Pelvin (PEL); Isla de Maipo (IM); San Francisco de Mostazal (SFM). Urban areas are shown in gray and agricultural areas are shown in green.
Collection of macroinvertebrates and fish samples. During the spring of 2016, between October and December, the four sampling sites were visited to collect macroinvertebrate and silverside (Basilichthys microlepidotus) samples. Fifteen macroinvertebrates samples were collected per site using a 30 × 30 cm Surber net with a 250 micron mesh filter, ensuring that all organisms adhered to rocks were carefully removed. Of these 15 samples, five were preserved in 95% ethanol to obtain community indices. The remaining 10 samples were stored in water and transported in a cooler to the laboratory where they were frozen until analysis: five were used to determine the lipid percentage and caloric content of the macroinvertebrates, while gastropods were extracted from the other five samples and analyzed for 15N isotopic content to be used as a baseline for trophic position calculations.
Fish were collected using a portable electric fishing device (Samus, 15 volts). Only adult fish measuring approximately 10 cm in standard length were targeted and transported to the laboratory; six fish were captured per site, for a total of 24 fish. A small piece of dorsal muscle tissue (weighing between 0.07 and 0.3 g wet weight) was dissected from each individual to measure the stable isotope 15N and ¹3C signal. From these specimens many parts were used for diverse analyses (liver, gills, muscle, digestive tract, fin, etc.) so there was no material left to be deposited in the museum, and the biological remains were discarded according to the approved protocols and those of the faculty. However, individuals of the same species and collected in the same basin of this study are available in the Ichthyological Collection of Museo Natural de Historia de Chile, Santiago (vouchers: MNHNCP 7373, 7372, and 7374).
Macroinvertebrate community indices. To determine the abundance and richness of the sampled macroinvertebrates in the environment and the stomach content, species identification and manual counting of individuals per species were conducted using a stereoscopic microscope (NIKON model SMZ-645). The species identification was performed following the dichotomous keys for benthic macroinvertebrates developed by Palma (2013). In this study, most of the macroinvertebrate organisms were identified to the family level. Using data on macroinvertebrate richness and abundance at each site, the following community indices were calculated: i) taxon richness, which accounts for the number of taxonomic groups detected, ii) total abundance, representing the number of individuals within each detected taxonomic group, iii) Shannon–Weaver diversity index (Shannon, Weaver, 1949), which incorporates the abundance of individuals per taxonomic group, iv) evenness index, which measures the uniformity of individuals’ distribution across the detected taxa (Pielou, 1966) and v) dominance index, which assesses the degree of representativeness of taxa within the community (Simpson, 1949). These indices were calculated using the software PAST v. 3.13 (Hammer et al., 2001).
Macroinvertebrates caloric and lipid content. Macroinvertebrates were dried in an oven at 60°C for approximately 24 h. Each zoobenthos sample was weighed both before and after drying. The energy content of the dried samples was determined using a Parr 1261 bomb calorimeter, with sample sizes ranging from 0.04 to 0.3 g of dried zoobenthos. For lipid content determination, the Soxhlet lipid extraction technique (Soxhlet, 1879) was employed, using methyl-tert-butyl ether as the solvent for approximately 2 h. After lipid extraction, the samples were dried and reweighed, and the lipid percentage was calculated gravimetrically. Additionally, we performed a spearman correlation in R software (R Development Core Team, 2024) between the average lipid percentage and the average macroinvertebrates caloric content and the average SMI body condition index for each population.
Silverside body condition index. We calculated the scaled mass index (SMI) (Peig, Green, 2009) of all silverside populations using the eviscerated weight and the total length of each individual. The analysis was performed in R using a custom script (R Development Core Team, 2024).
Alimentary preference of B. microlepidotus. Using the stomach content data, the Manly’s alpha food selectivity index (Krebs, 2014) was calculated, which allows determining whether there is a preference for a particular food item by relating the proportion of each item in the stomach to that in the environment. After calculating Manly’s alpha index, it was standardized using Chesson’s equation (Krebs, 2014). Chesson’s index provides values ranging from +1 to -1, where positive values close to 1 indicate preference for a food item, negative values close to -1 indicate rejection, and values near 0 indicate that the item is consumed according to its environmental availability (Krebs, 2014).
Stable isotope δ¹⁵N and δ¹3C signal and trophic position of B. microlepidotus. To assess differences in the stable isotope δ¹⁵N and δ¹3C signal and the trophic position of B. microlepidotus between the non-polluted site, the polluted sites, and the different channel types, the stable isotope δ¹⁵N and δ¹3C signal was analyzed in the muscles of fish and in gastropods (genera Physa and Littoridina) from each site. The gastropods were used as a baseline reference.
Muscle tissue samples from each silverside and the gastropods were dried in an oven at 60°C for approximately 24 h and then weighed. Once dried, both the silverside muscle samples, and the gastropods were defatted using the Soxhlet method to remove the isotopic signal of ¹³C present in the carbon chains of lipids. All samples were sent to the Laboratory of Biogeochemistry and Applied Stable Isotopes (LABOSI) at the Pontificia Universidad Católica de Chile, where an Isotope Ratio Mass Spectrometer (IRMS, Thermo Delta Advantage) coupled to a Flash EA2000 Elemental Analyzer was used to assess the 15N and ¹3C isotopic content of each sample. The standards used were acetanilide, atropine, caffeine, high organic sediment, and glutamic acid.
The silverside trophic position was estimated using the tRophicPosition package (Quezada-Romegialli et al., 2018) and an average enrichment per trophic level of 3.4‰ (Minagawa, Wada, 1984; Post, 2002). Trophic enrichment factors for δ¹⁵N in fish typically range from 2.5‰ to 4.3‰ depending on feeding ecology, with omnivorous species averaging 3.4‰ which could be the case for B. microlepidotus (Stephens et al., 2023; Letourneur et al., 2024). According to Post (2002) snails represent a suitable baseline for quantifying the trophic position of secondary organisms in aquatic ecosystems and Lake et al. (2019) showed that the use of different organisms as baseline, including gastropods do not affect the trophic position estimation. Moreover, it has been stated that it is useful to consider site specific baseline to account for spatial variability when stable isotopes analysis is performed in degraded environments (Braun et al., 2018; Qiu et al., 2023). Taking this into consideration Physa sp. And Littoridina sp. were used as baseline, since they present an isotopic signature similar to that of the primary producers (periphyton and detritus). Therefore, based on Post (2002) the baseline trophic-position value in our study was considered equal to 2. The analysis was run using 10,000 iterations, 10,000 adaptive iterations, 5 parallel chains and a burning of 10,000. We also statistically tested the trophic position estimates considering condition and watercourse type using the pairwiseComparison function implemented in tRophicPosition package.
Statistical analysis. To assess the richness and abundance and to detect differences or overlaps in the composition of the macroinvertebrate community between conditions and sites, a non-metric multidimensional scaling analysis (NMDS) was performed using the relative abundance of each taxa at family level in the environmental data and the stomach content of silverside. Stress analysis was used to validate the data representation through dimensionality reduction. Additionally, a PERMANOVA analysis was performed to test statistical differences in environmental and stomach content data in R software (R Development Core Team, 2024). A contrast-based ANOVA was used to compare total abundance and community indices in both environmental and stomach content, caloric and lipid content, δ¹⁵N signals for silverside and snails, and body condition, grouped by watercourse type (central-channel vs. channel) and pollution level (polluted vs. non-polluted). Analyses were performed using the “multcomp” package in R (R Development Core Team, 2024). Residual normality was tested with the Shapiro-Wilk test in R, showing no significant deviations except for the macroinvertebrate dominance index in the environment, which was log-transformed. Homoscedasticity was assessed using the Fligner-Killeen test, with no significant variance differences detected.
Results
Macroinvertebrate community and stomach content indices. A total of 17 macroinvertebrate families belonging to 11 orders and six classes (Tab. S2) were found in the four sampling sites. The classes Insecta and Gastropoda were the most abundant, with 60.52% and 26.47% of the total abundance, respectively. When sites were grouped according to their condition (polluted vs. non-polluted), Insecta represented approximately 60% of the abundance in the polluted sites (IM, MEL, and PEL), and Gastropoda approximately 30%. In the non-polluted site (SFM), Insecta represented approximately 84% of the total abundance, followed by individuals of the phylum Nematoda, which represented approximately 14% of the total abundance. When sampling sites were grouped by watercourse type, Gastropoda accounted for approximately 59.2% of the abundance of macroinvertebrates in channel-type sites (IM and PEL), followed by Insecta, with 25.5% of the total density. However, in central site types (SFM and MEL), Insecta accounted for 86.5% of the abundance of macroinvertebrates in these sites.
The NMDS analysis of the relative abundance and diversity of macroinvertebrate families in the environment showed site segregation along the first axis, separating the central-channel-type sites (SFM and MEL) from the channel-type sites (IM and PEL). The sites could not be clearly differentiated based on their pollution status (Fig. 2A). The results indicated that the families Chironomidae, Athericidae, and Physidae were predominantly represented in central-channel-type sites; in contrast, the families Hydroptilidae and Hydrobiidae were mainly represented in channel-type sites. The NMDS graph indicates that channel type appears to be an important environmental factor influencing the diversity and relative abundance of benthic macroinvertebrates taxa present at each site, with sites of the same channel type showing similar diversity and abundance. The stress analysis showed a value of 0.031 (<0.1) validating the results obtained. Nevertheless, the PERMANOVA analysis showed a statistically significant difference for both factors (Pollution condition: p < 0.05; Watercourse type: p < 0.001), pointing out that pollution could have some impact on macroinvertebrate diversity and abundance, but not as strong as the watercourse type factor.
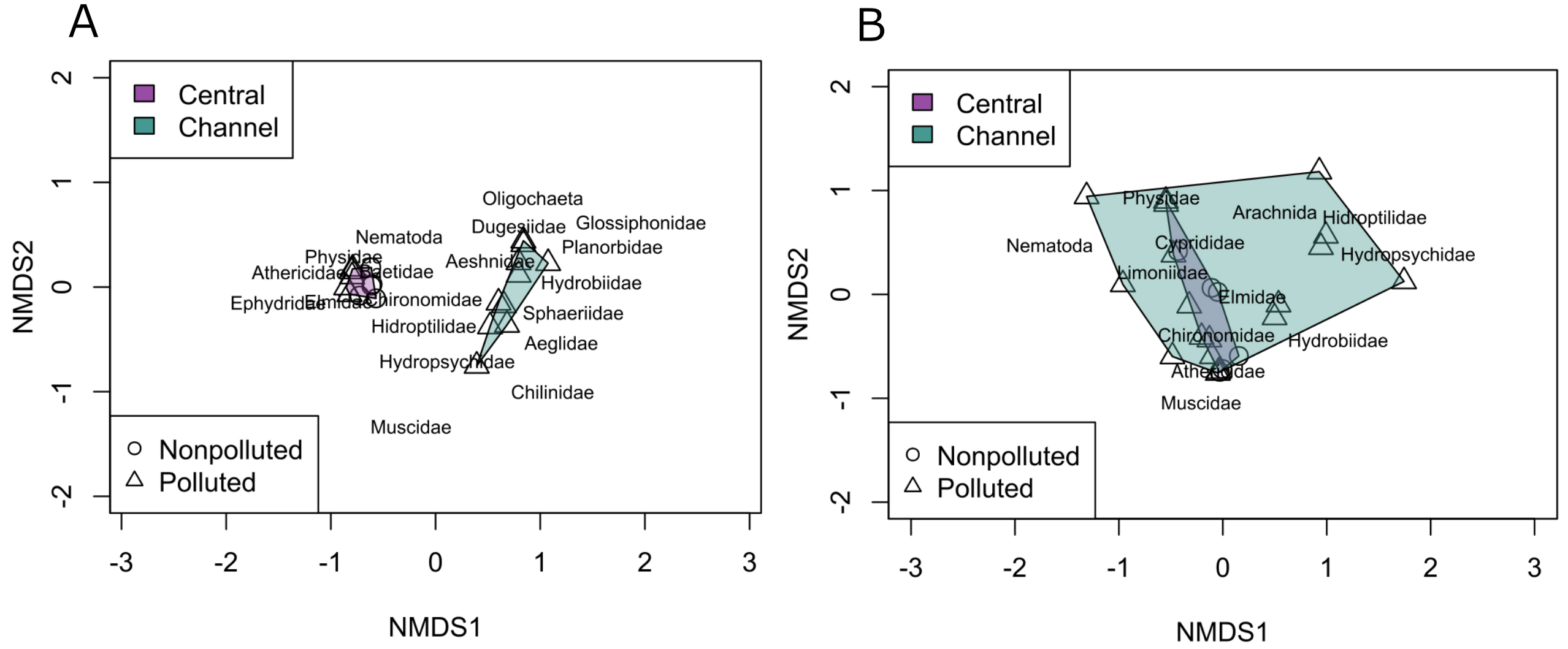
FIGURE 2| Non-metric multidimensional scaling of macroinvertebrates’ relative abundance. Environmental data (stress value of 0.035) (A) and stomach content (stress value of 0.099) (B). The macroinvertebrate families are shown along with the phylum Nematoda and the class Oligochaeta.
The NMDS analysis of the relative abundance and diversity of macroinvertebrate families in the stomach content showed an overlap between central-channel-type sites and channel-type sites, but channel-type sites present a broader amplitude than the central-channel-type sites (Fig. 2B). Regarding the pollution condition factor an overlap is also observed. The stress analysis produced a value of 0.099 and the PERMANOVA analysis only showed statistical differences when the watercourse type factor is considered (p < 0.05), and no differences were found when pollution condition factor was considered (p = 0.25).
The ANOVA results for the environmental community indices showed that watercourse type and pollution had no effect on the variables of total abundance (pollution factor: F = 0.172, p = 0.866; watercourse type factor: F = -0.953, p = 0.355) (Tab. 1) and dominance index (pollution factor: F = 1.552, p = 0.140; watercourse type factor: F = -1.483, p = 0.158). This indicates that the abundance of the environmental food supply for the silverside did not show differences across the study sites. Additionally, the sites presented a dominance index value close to 0.5 (Tab. 1), indicating an intermediate level of dominance of macroinvertebrate taxa in the environment. However, the ANOVA revealed a significant effect of pollution on the Shannon index (F = 3.806; p = 0.00155) and taxon richness (F = 2.384; p = 0.0298) (Figs. 3A, C). In both cases, the non-polluted site (SFM) exhibited a significantly higher Shannon index and taxon richness compared to the more-polluted sites in the Maipo basin. For the watercourse type, the ANOVA revealed a significant effect of this variable on the evenness index (F = 2.740; p = 0.0145) and Shannon index (F = 5.466; p < 0.0001) (Figs. 3D, F). In both cases, the channel-type sites showed significantly higher evenness and Shannon index values compared to the central-channel-type sites.
TABLE 1 | Diversity indices for each sampling site. The sites’ classification according to condition and watercourse type is shown. Mean and standard deviation (±) of all the replicates per site are shown.
Condition | Non-polluted | Polluted | Polluted | Polluted |
Watercourse | Central | Channel | Central | Channel |
Site | SFM | IM | MEL | PEL |
Abundance | 303.8 ± 94.14 | 213.8 ± 127.42 | 311.6 ± 108.93 | 243.8 ± 125.27 |
Taxa Richness | 7 ± 1.41 | 5.4 ± 1.14 | 5.2 ± 1.10 | 6.2 ± 1.10 |
Shannon index | 1.18 ± 0.18 | 1.05 ± 0.09 | 0.82 ± 0.21 | 1.34 ± 0.08 |
Dominance index | 0.44 ± 0.07 | 0.46 ± 0.07 | 0.60 ± 0.12 | 0.33 ± 0.04 |
Evenness index | 0.47 ± 0.04 | 0.55 ± 0.11 | 0.46 ± 0.12 | 0.63 ± 0.10 |
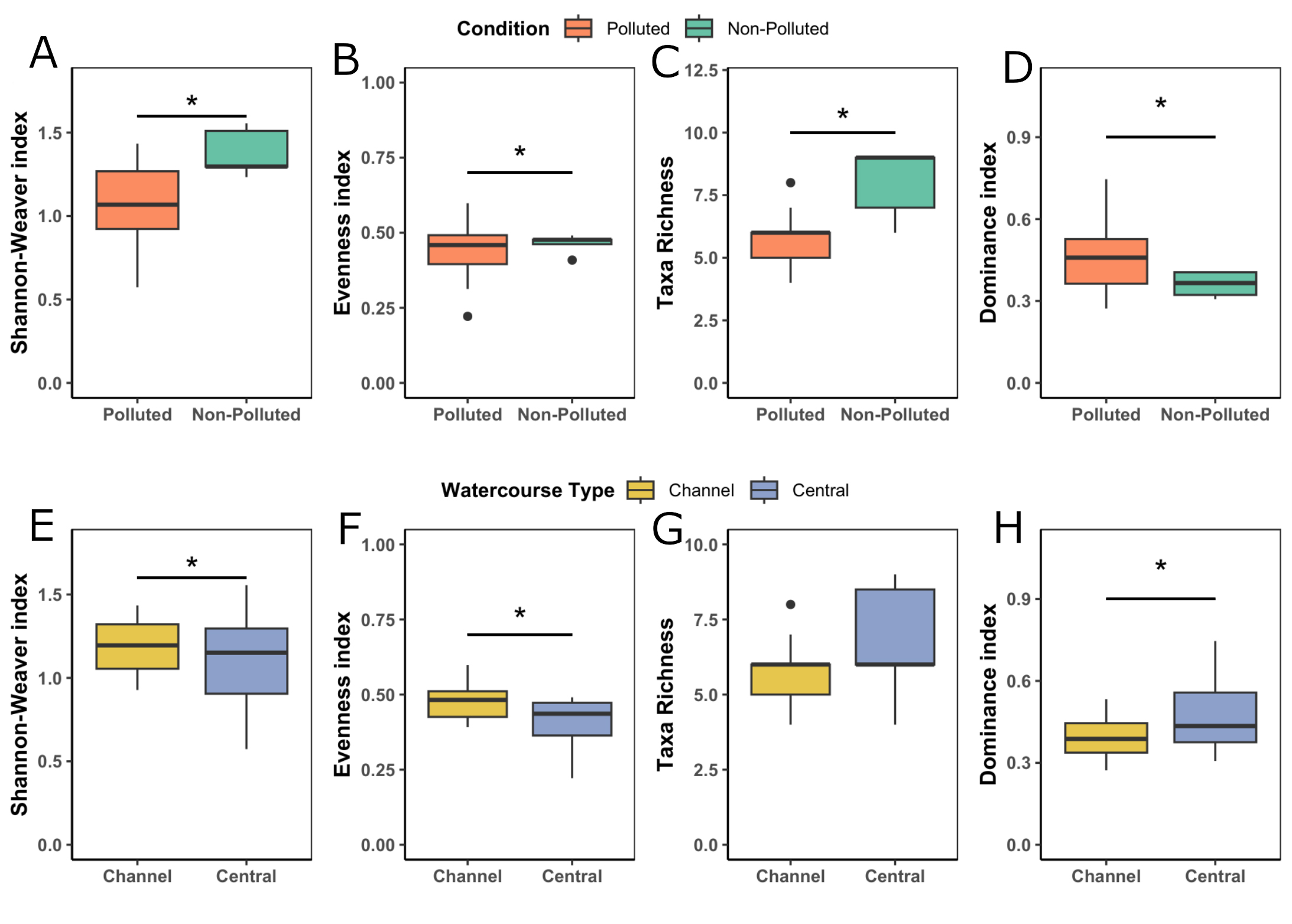
FIGURE 3| Boxplots of diversity indices compared across polluted and non-polluted sites in environmental data: A. Shannon-Weaver, B. Evenness index, and C. Taxon richness. Boxplots of diversity indices compared across watercourse types in environmental data: D. Shannon-Weaver, E. Evenness index, and F. Taxon richness. *Indicate significant differences.
Regarding the stomach content the ANOVA showed that the watercourse type and pollution had no effect on taxa richness variable (pollution factor: F = -0.341, p = 0.737; watercourse type factor: F = 0.682, p = 0.503) but it does affect the abundance variable (pollution factor: F = -2.382, p = 0.027; watercourse type factor: F = -3.655, p = 0.001), with non-polluted and central-channel-type sites showing more abundance than polluted and channel-type sites (Figs. 4A, E). Pollution factor had no effect on Shannon index (F = 0.738, p = 0.4692), dominance index (F = -0.588, p = 0.5631) and evenness index (F = 1.629, p = 0.119) (Figs. 4B–D) contrary to what was observed in the watercourse type factor, which does show an effect on these indices (Shannon index, F = 2.428, p = 0.0248; dominance index, F = -2.102; p = 0.0484; evenness index, F = 2.803; p = 0.011) (Figs. 4F–H). This means that pollution does not affect what silverside consumes but macroinvertebrates abundance consumed is affected. In the case of watercourse type factor differences in abundance and diversity of what is consumed are detected, with channel-type sites showing higher values of Shannon index and evenness index, but lower values of dominance index.
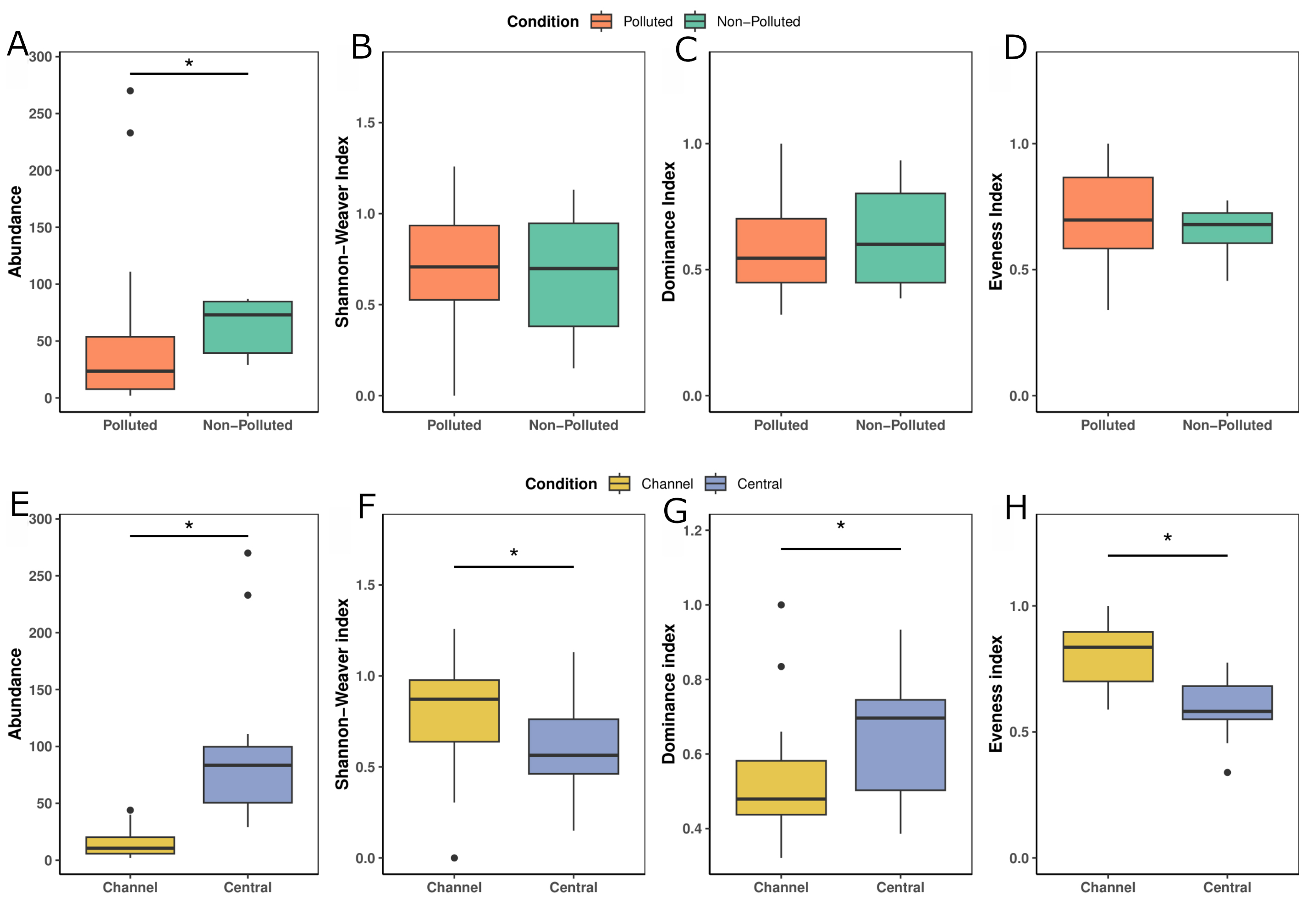
FIGURE 4| Boxplots of diversity indices compared across polluted and non-polluted sites in stomach content: A. Abundance, B. Shannon-Weaver, C. Dominance index, and D. Evenness index. Boxplots of diversity indices compared across watercourse types in stomach content: E. Abundance, F. Shannon-Weaver, G. Dominance index, and H. Evenness index.
Macroinvertebrates caloric and lipid content. Regarding the quality of macroinvertebrates as a food source, the ANOVA by contrast showed a significant effect of pollution (F = 2.873; p = 0.011) and watercourse type (F = -7.023; p < 0.0001) on the caloric content of macroinvertebrates. In this analysis, the non-polluted site exhibited higher macroinvertebrate caloric content (Fig. 5A). Moreover, the central-channel sites showed higher macroinvertebrate caloric content compared to that of channel-type sites (Fig. 5B). Additionally, the ANOVA by contrast revealed the near-significant effect of pollution (F = 2.050; p = 0.05962) (Fig. 5C) and the significant effect of watercourse type (F = -4.865; p = 0.00025) on the macroinvertebrate lipid percentage. The central-channel sites exhibited a higher macroinvertebrate lipid percentage compared to that of channel-type sites (Fig. 5D).
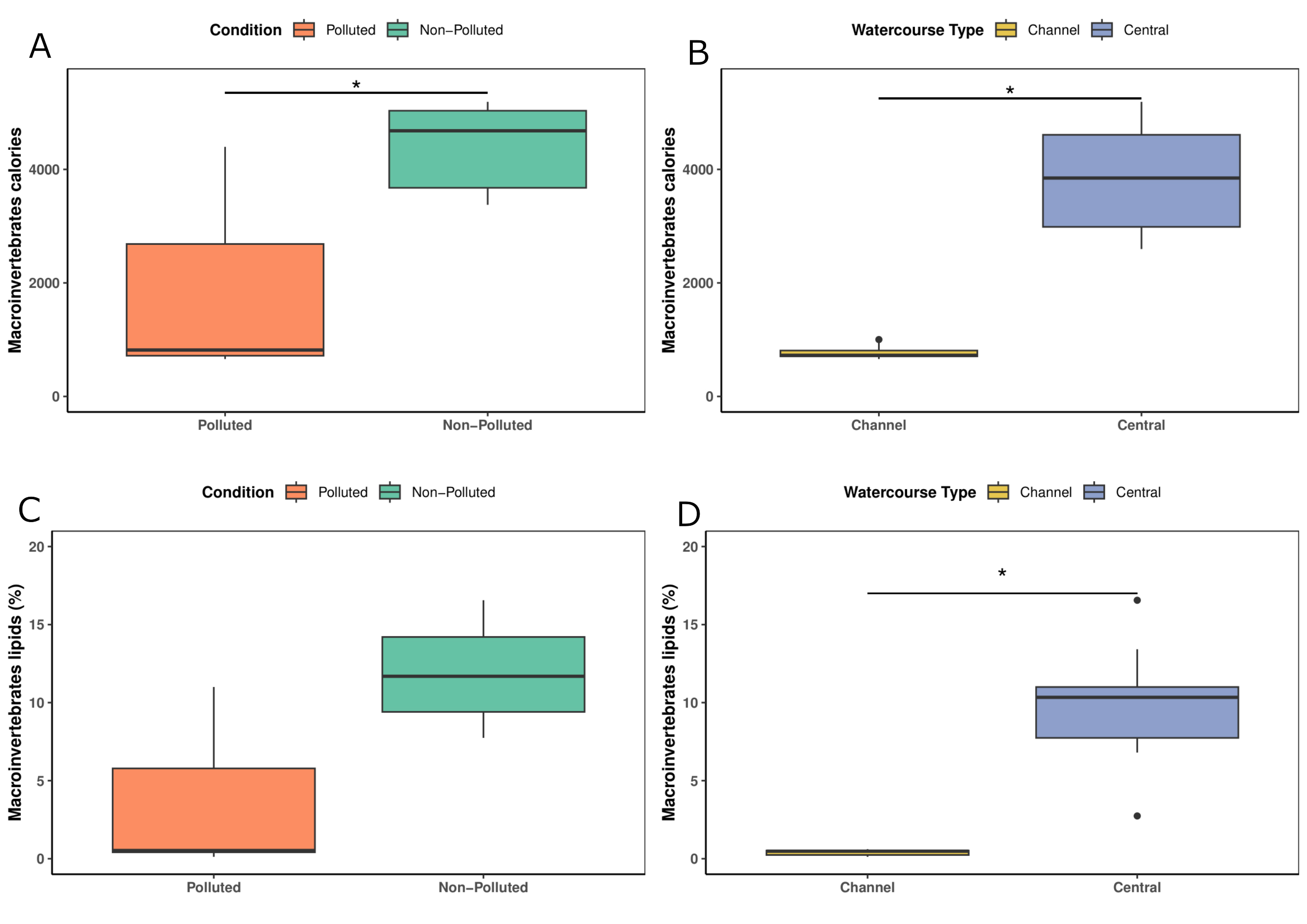
FIGURE 5| Boxplots of caloric content and lipid percentage of macroinvertebrates: A. Caloric content of macroinvertebrates in polluted and non-polluted sites, B. Caloric content of macroinvertebrates in channel-type and central channels, C. Lipid percentage of macroinvertebrates in polluted and non-polluted sites, and D. Lipid percentage of macroinvertebrates in channel-type and central channels. *Indicate significant differences.
Silverside SMI body condition factor. The scaled mass index showed similar values among populations (SFM: mean = 5.78, sd = 0.604; IM: mean = 5.62, sd = 0.262; MEL: mean = 5.73, sd = 0.228; PEL: 5.36, sd = 0.470) and no statistical differences were found among them (p > 0.05 among all comparisons). When we grouped populations considering pollution factor, we also found similar SMI values (non-poluted: mean = 5.78, sd = 0.604; polluted: mean = 5.57, sd = 0.354) and no statistical differences (p > 0.05). Similarly, when populations were grouped according to watercourse type, we obtained similar SMI values (central-channel-type: mean = 5.75, sd = 0.436; channel-type: mean = 5.49, sd = 0.378) and no statistical differences (p > 0.05).
The correlation between the average lipid and caloric content of macroinvertebrates with the average of condition factor for each site were not statistically significant (lipid content: p = 0.083; caloric content: caloric content: p = 0.083), showing no direct relationship between the quality of the macroinvertebrates and body condition in silverside.
Alimentary preference of B. microlepidotus. The Chesson index showed selection by the gastropod Physidae in the channel-type, central-channel-type, and polluted sites, while in the least polluted site there was selection by the Diptera Muscidae and Athericidae.
Stable isotope δ¹⁵N signal and trophic position of B. microlepidotus and baseline. The significant effects of pollution (F = -16.513; p < 0.0001) and watercourse type (F = -9.988; p < 0.0001) on the stable isotope δ¹⁵N signal were detected in silverside, although the effect of the pollution factor was much clearer, with individuals from the polluted sites exhibiting higher signal of the stable isotope δ¹⁵N than individuals from the non-polluted site (Figs. 6A, C). Regarding the baseline we can observe a similar pattern with significant effects of pollution (F = -22.67; p < 0.0001) and watercourse type (F = -11.66; p < 0.0001) with polluted and central-channel-type sites showing higher isotope δ¹⁵N signal than non-polluted or channel-type sites, respectively (Figs. 6B, D).
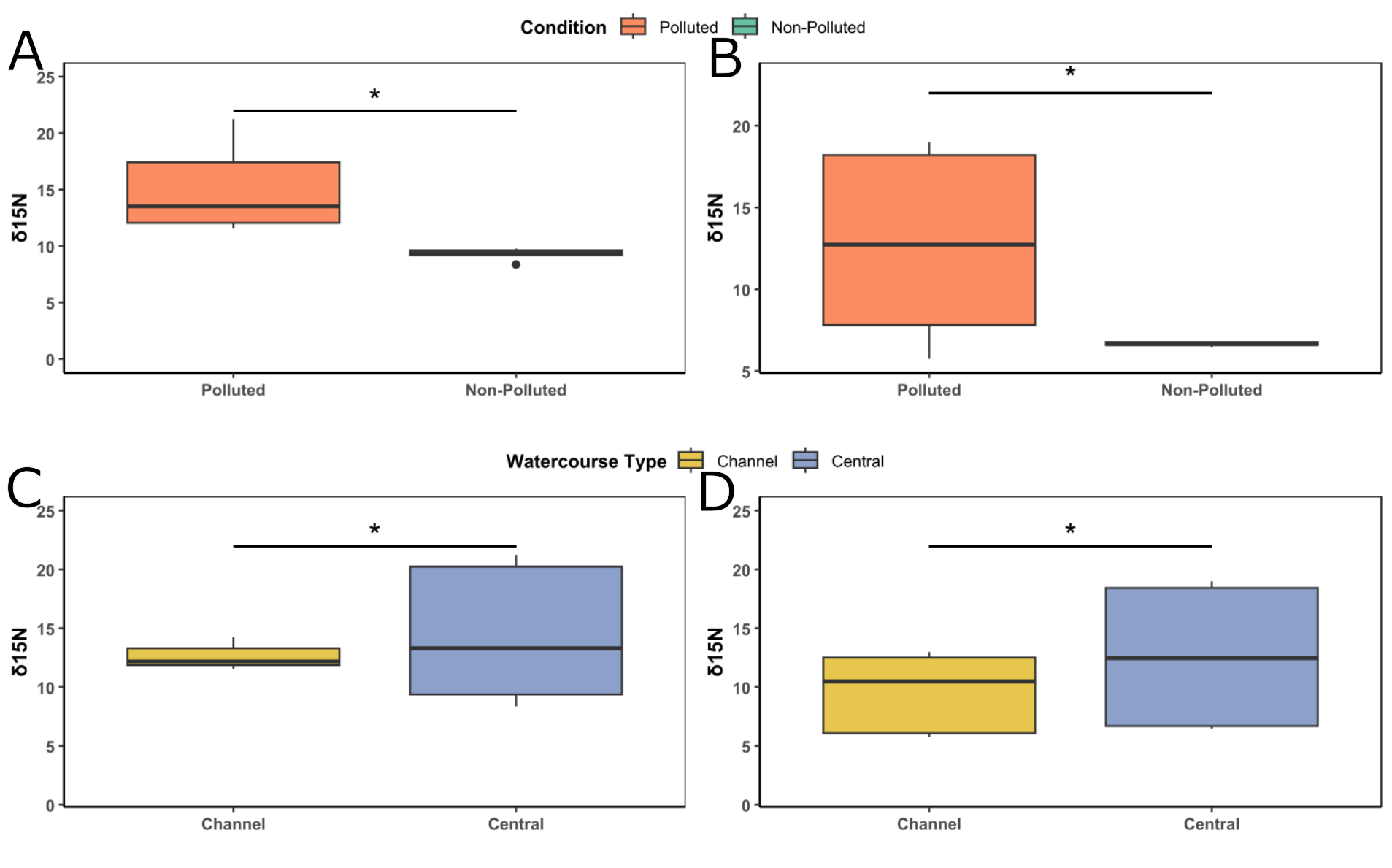
FIGURE 6| δ15N signal in muscle tissue of Basilichthys microlepidotus and baseline: A. δ15N content in silverside in polluted and non-polluted sites, B. δ15N content in silverside in channel and central watercourse types, C. δ15N content in baseline in polluted and non-polluted sites, D. δ15N content in baseline in channel and central watercourse types. *Indicate significant differences.
No effect of pollution was observed on trophic position, with non-polluted sites showing credibility intervals ranging between 2.565 and 2.991 and polluted sites ranging between 2.072 and 3.786, with a posterior mode of 2.774 and 2.705, respectively. Similarly, when watercourse type was considered, no effect was observed over trophic position with credibility intervals ranging between 2.044 and 4.406 for central-channel-type sites and 2.208 and 3.678 for channel-type sites, with a posterior mode of 2.731 and 2.877, respectively. Nevertheless, when δ¹⁵N and δ¹3C signals were considered, it is observed a consistent pattern related to the δ¹⁵N signal, with polluted sites showing in most cases a higher value, in both silverside (Fig. 7A) and the baseline (Fig. 7B). Regarding the δ¹3C signal, polluted sites showed a high variability. In the case of watercourse type the effects are not clear, with central-channel-type sites showing a very contrasting pattern.
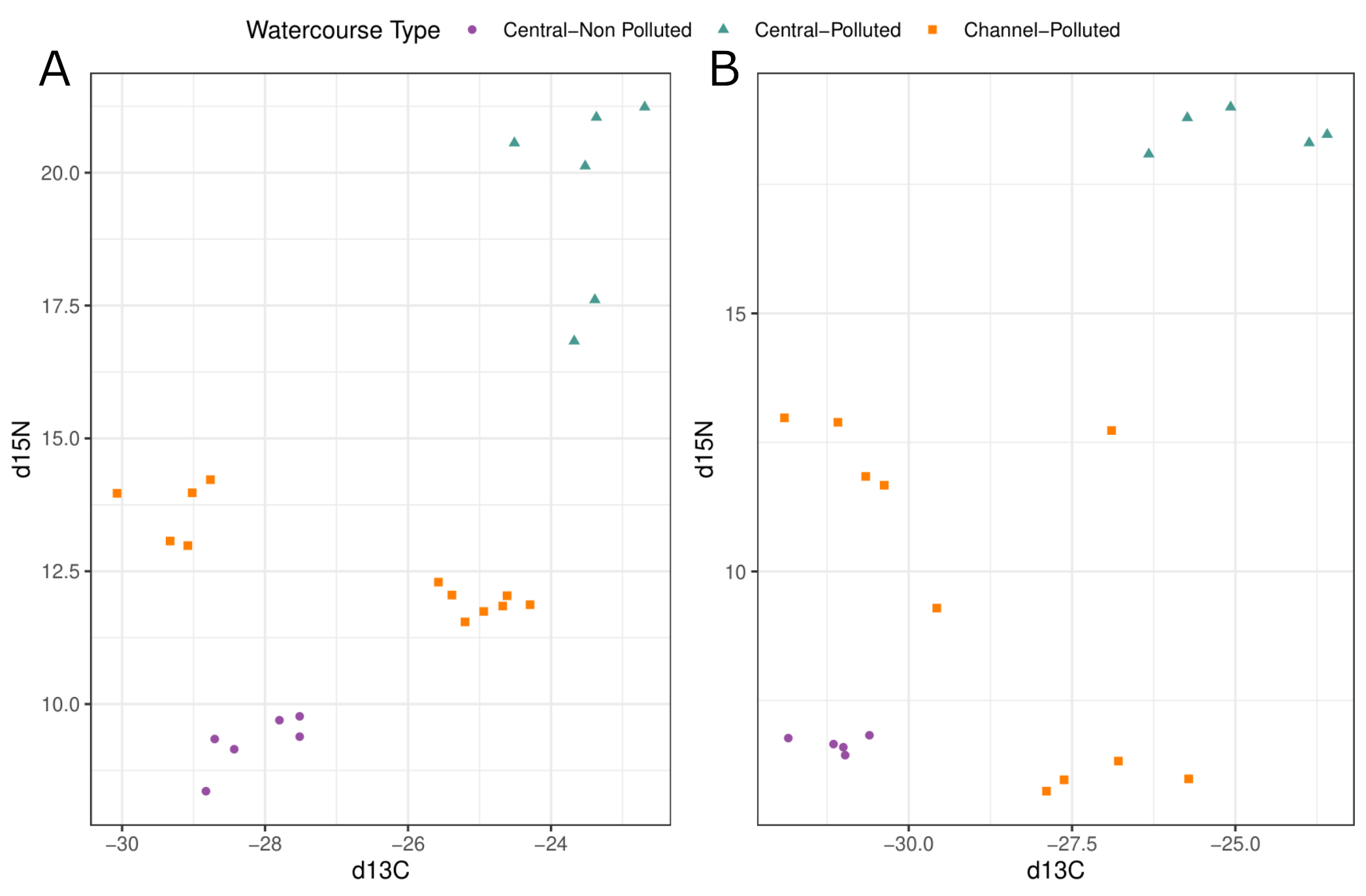
FIGURE 7| Biplot of δ15N and δ13C signals in Basilichthys microlepidotus and baseline. Silverside biplot (A) and baseline biplot (B).
Discussion
Our study suggests that water pollution and river channelization not only affect the composition of the macroinvertebrate community but also its nutritional quality as a food resource for fishes, although no changes in the body factor condition and silverside’s trophic position were observed.
The macroinvertebrate community in the Maipo River resembled patterns seen in polluted sites elsewhere, with high abundances of Oligochaeta, Nematoda, Chironomidae, and Gastropoda (Azrina et al., 2006; Theodoropoulos et al., 2015). In Chile, Córdova et al. (2009) reported elevated percentage of Chironomidae, Dugesiidae, Hydropsychidae, and gastropods in polluted areas of the Limache stream, while less-impacted sites hosted more Perlidae and Leptophlebiidae. In our study, Chironomidae were dominant across all sites, while sensitive groups like Ephemeroptera and Plecoptera were scarce, consistent with the Maipo River’s proximity to a large urban center. Notably, macroinvertebrate composition varied more by watercourse type than by pollution level. Channel-type channels exhibited a greater abundance of gastropods of the genus Littoridina, whereas central-channel types showed higher abundances of insects, particularly Chironomidae. Similarly, Habit et al. (1998) described distinct benthic macroinvertebrate communities between irrigation channels and the main channel of the Itata River in the south-central region of Chile. Our results suggest that, in the case of the Maipo River basin, watercourse type is a more reliable indicator than pollution level to assess macroinvertebrate community composition. Moreover, a similar pattern was observed in the stomach content analysis, where the different watercourse types showed variations in trophic niche breadth. Channel-type sites exhibited greater diversity in gut contents compared to the central channel. Regarding pollution, the stomach contents of silverside were similar under both conditions, indicating that despite differences in environmental macroinvertebrates availability between polluted and non-polluted sites, silverside consumed similar types of macroinvertebrates. Da Silva Penha et al. (2025) found similar patterns/results for the fish Hemiodus unimaculatus in the Xingu River where the species showed a stable pattern in the diet across different hydrological periods with different environmental offers. It is important to consider that B. microlepidotus has a broader diet including small invertebrates, insect larvae, detritus, and filamentous algae (Bahamondes et al., 1979), suggesting that a shift in diet could happen but not at macroinvertebrate level.
Not only differences in community structure were found, as silverside in polluted and channel-type sites showed access to food with lower caloric and lipid content compared to those in non-polluted and central-channel sites. Few studies have examined macroinvertebrate nutritional quality for fish. Cobo et al. (2000) highlighted the need to consider both the quantity and quality of macroinvertebrates to assess trophic impacts. They reported the caloric and lipid content of insects (Coleoptera, Trichoptera, and Diptera), with an average energy content of 5,426.58 cal/g and a lipid content of 22.62%. These results matched values from less-impacted sites in this study, likely due to greater insect abundance. In contrast, gastropods were dominant in channel and polluted areas. Hondolero et al. (2012) estimated the caloric content of a group of gastropods in the northern Bering and Chukchi Sea as approximately 766.7 cal/g, a value comparable to the caloric content observed in this study for channel-type and polluted sites (776.4 cal/g and 1627.4 cal/g, respectively). Heras et al. (1998) determined the lipid content of juveniles of the gastropod Pomacea canaliculata (Lamarck, 1828), reporting a value of 2% lipid content in dry weight, which closely matches the lipid content measured in macroinvertebrates from the polluted and channel-type sites in this study (3.03% and 0.39%, respectively).
Caloric and lipid content across sites reflected the dominant macroinvertebrate types, with gastropods providing lower nutritional value than insects. This may affect consumers like B. microlepidotus. Polluted sites showed reduced diversity and food quality, aligning with gene expression patterns found by Vega-Retter et al. (2018). Interestingly, channel sites had lower food quality but higher diversity than central channels, suggesting a complex relationship between macroinvertebrate diversity, nutritional value, and fish condition that warrants further study. Despite the differences observed in the nutritional quality of macroinvertebrates among sites, at a broader scale no effect was detected on the silverside diet composition between polluted and non-polluted areas and, similarly, no differences in the body condition factor were observed by pollution and watercourse type. These two last results mentioned could be associated with the selection behavior of B. microlepidotus in almost all sites by the gastropod Physidae. This suggests that Physidae would be an easy prey for B. microlepidotus, since Physidae is attached to rocks at the bottom of the channel and has low mobility (Hanson et al., 2010), thus increasing its probability of capture. Similarly to our results, Davis et al. (2012) used stomach content and stable isotope analyses to assess size-related dietary shifts in an Australian fish assemblage. All species showed marked ontogenetic changes in diet. While δ¹³C values generally agreed with stomach content data, concordance with δ¹⁵N-based trophic position estimates was low, likely due to omnivory, overlapping basal isotope sources, and uncertainty in isotopic fractionation rates. Interestingly, silverside inhabiting polluted sites appeared to actively select macroinvertebrates with lower nutritional content. A possible explanation for this pattern could be a shift in carbon sources within polluted environments, as suggested by the δ¹³C signal observed at those sites. A comparable phenomenon has been reported for native and exotic fish species in the rio das Velhas basin, Brazil, where a shift in carbon sources occurs in polluted areas, changing from filamentous algae and periphyton in cleaner sites to sewage-derived and fine particulate organic matter in polluted sites (de Carvalho et al., 2019). This could be in line with the broad feeding behavior of silverside, that could differ in other components besides macroinvertebrates in polluted sites. For example, Alves et al. (2016) showed that the silverside Atherinella brasiliensis presents a generalist feeding behavior changing its food sources in polluted environments including items not related to macroinvertebrates. Nevertheless, this plasticity in feeding behavior in polluted sites could have consequences, like liver damage (Santos et al., 2018). Future studies in the Maipo River basin should analyze different carbon sources to test this hypothesis.
Individuals of B. microlepidotus inhabiting polluted sites exhibited higher δ¹⁵N values compared to those from the non-polluted site. Similarly, de Carvalho et al. (2020) conducted a study across 16 sites within the rio das Velhas basin, a Brazilian river heavily impacted by anthropogenic activity, and found that δ¹⁵N values in both periphyton and catfishes reflected the concentrations of reactive nitrogen introduced by diverse human activity. Accordingly, our findings suggest that the polluted sites (IM, MEL, and PEL) are subject to a greater environmental nitrogen load. Additionally, the gastropods used as baseline showed the same pattern related to pollution factor. This is also supported by the findings of Briones-Alburquenque et al. (2025), who conducted research on three of the sites examined in the present study and reported higher δ¹⁵N values in the feeding functional groups of macroinvertebrates inhabiting the polluted sites compared to those from less-polluted sites. Despite this, the trophic position of B. microlepidotus was not affected either by pollution or watercourse type. The trophic position of an organism can be influenced by three main factors: the isotopic baseline of the environment, primarily associated with the incorporation of organic and nitrogenous contaminants, which tend to elevate the trophic position of all exposed organisms (Lee et al., 2018), the physiological or nutritional condition of the organism (Hobson et al., 1993; Doucett et al., 1999), and dietary habits, specifically the direct composition of the organism’s diet (Post, 2002; Caut et al., 2009). In this study we observe effects at macroinvertebrate communities related to pollution and watercourse type factors, and previous studies had clearly shown the chemical differences among sites (Vega-Retter et al., 2014; Cortés-Miranda et al., 2024), but this does not affect silverside at broader scale like trophic position or body condition factor. This could be explained by the carbon shift sources as was discussed above but also could be related to the δ¹⁵N enrichment observed in the baseline related to pollution. Nevertheless, it does not mean that the changes in macroinvertebrate structure and quality could not have an effect on other fish species not evaluated in this study.
Another factor to consider is that our sampling was conducted during a single season (spring). To assess whether the detected pattern remains consistent across different periods of the year, it is essential to replicate this study in the autumn, winter and summer. Additionally, not considering other potentially relevant factors like water temperature, climate, geomorphological factors could limit the interpretation of the results. However, it is important to note that the four sampling sites used in this study have been monitored by our team for over 10 years, with physicochemical data collected in the years both prior to and following sampling for this study indicating temporal stability in the level of contamination at these sites (Vega-Retter et al., 2014; Veliz et al., 2020; Cortés-Miranda et al., 2024). However, as we show in Cortés-Miranda et al. (2024) the pollution profile of the three polluted sites is not the same, with a clear separation of MEL from IM and PEL, which could be acting as an additional confounding factor, thereby hindering the detection of the effects of pollution. Furthermore, another study has confirmed our team’s characterization of the pollution in the basin (Soriano et al., 2024). Given the high reliability of our site characterization, our results appear to be consistent, but it would be necessary to consider other variables in future studies to confirm this.
Overall, this study suggests that anthropogenic activities such as water pollution and river channelization not only affect the composition of the macroinvertebrate community but also its nutritional quality as a food resource for fishes. Most macroinvertebrates indexes used to assess environmental conditions only consider changes in abundance and/or composition, but quality of macroinvertebrate as food is neglected (Sumudumali, Jayawardana, 2021; Ndatimana et al., 2023), even considering that previous studies emphasis this problem (Bertoli et al., 2021). Our results point out that change in macroinvertebrate community composition can be associated with change in their caloric and lipid content, which could have several impacts on aquatic ecosystems. The absence of an effect on the silverside’s trophic position is controversial but may reflect a dietary shift not directly related to the macroinvertebrate community. Finally, our findings indicate that anthropogenic disturbances may not necessarily reduce the total abundance of macroinvertebrates, but they can alter community composition and, consequently, the quality of this resource for higher trophic levels, an aspect often overlooked in environmental assessments. In our study, the consumer species was not affected; however, this outcome should be evaluated for other consumers that may have very different ecological requirements. Given that macroinvertebrates play a key role in linking primary producers and higher trophic levels, it is essential to consider this dimension in environmental impact assessments, for instance, by developing indices that incorporate this measure, to support appropriate conservation and management strategies for aquatic ecosystems and fish diversity.
Acknowledgments
The authors thank to Rachel Gaucci for helping in the field work.
References
Alves VEN, Patrício J, Dolbeth M, Pessanha A, Palma ART, Dantas EW et al.Do different degrees of human activity affect the diet of Brazilian silverside Atherinella brasiliensis? J Fish Biol. 2016; 89(2):1239–57. https://doi.org/10.1111/jfb.13023
Azrina MZ, Yap CK, Rahim Ismail A, Ismail A, Tan SG. Anthropogenic impacts on the distribution and biodiversity of benthic macroinvertebrates and water quality of the Langat River, Peninsular Malaysia. Ecotoxicol Environ Saf. 2006; 64(3):337–47. https://doi.org/10.1016/j.ecoenv.2005.04.003
Bahamondes I, Soto D, Vila I. Hábitos alimentarios de las especies de Atherinidae del Embalse Rapel. Medio Ambiente. 1979; 4:3–18.
Belcheva N, Istomina A, Dovzhenko N, Lishavskaya T, Chelomin V. Using heavy metal content and lipid peroxidation indicators in the tissues of the mussel Crenomytilus grayanus for pollution assessment after marine environmental remediation. Bull Environ Contam Toxicol. 2015; 95(4):481–87. https://doi.org/10.1007/s00128-015-1624-3
Bertoli M, Piazza G, Pastorino P, Prearo M, Cozzoli F, Vignes F et al.Macrobenthic invertebrate energy densities and ecological status in freshwater watercourses (Friuli Venezia-Giulia, Northeast Italy). Aquat Ecol. 2021; 55(2):501–18. https://doi.org/10.1007/s10452-021-09840-x
Braun DP, Phillips ID, Nanayakkara L, Wissel B. Diet characterization and a preliminary investigation into trophic niche placement for an endangered lake sturgeon (Acipenser fulvescens) population in the Saskatchewan River, SK, Canada. PLoS ONE. 2018; 13(11):e0206313. https://doi.org/10.1371/journal.pone.0206313
Briones-Alburquenque MP, Cortés-Miranda J, Vega-Retter C. Pollution effects on diversity and content of the stable isotope 15N in macroinvertebrates: a study of the impacts on the functional groups from the Maipo River, Chile. Mar Freshw Res. 2025; 76(8):MF24234. https://doi.org/10.1071/MF24234
Brito EF, Moulton TP, Souza ML, Bunn SE. Stable isotope analysis indicates microalgae as the predominant food source of fauna in a coastal forest stream, south-east Brazil. Austral Ecol. 2006; 31(5):623–33. https://doi.org/10.1111/j.1442-9993.2006.01610.x
de Carvalho DR, Alves CBM, Flecker AS, Sparks JP, Moreira MZ, Pompeu PS. Using δ15N of periphyton and fish to evaluate spatial and seasonal variation of anthropogenic nitrogen inputs in a polluted Brazilian river basin. Ecol Indic. 2020; 115. https://doi.org/10.1016/j.ecolind.2020.106372
de Carvalho DR, Flecker AS, Alves CBM, Sparks JP, Pompeu PS. Trophic responses to aquatic pollution of native and exotic livebearer fishes. Sci Total Environ. 2019; 681:503–15. https://doi.org/10.1016/j.scitotenv.2019.05.092
Caut S, Angulo E, Courchamp F. Variation in discrimination factors (∆15N and ∆13C): the effect of diet isotopic values and applications for diet reconstruction. J Appl Ecol. 2009; 46(2):443–53. https://doi.org/10.1111/j.1365-2664.2009.01620.x
Cobo F, Mera A, González M. Análisis químico y contenido energético de algunas familias de insectos holometábolos dulceacuícolas. Nova Acta Ci Compostelana Biol. 2000; 10:77–86.
Comte S, Vila I. Spawning of Basilichthys microlepidotus (Jenyns). J Fish Biol. 1992; 41(6):971–81. https://doi.org/10.1111/j.1095-8649.1992.tb02724.x
Cooper CM. Biological effects of agriculturally derived surface water pollutants on aquatic systems – a review. J. Environ Quality. 1993; 22(3):402–08. https://doi.org/10.2134/jeq1993.00472425002200030003x
Córdova S, Gaete H, Aránguiz F, Figueroa R. Evaluación de la calidad de las aguas del estero Limache (Chile central), mediante bioindicadores y bioensayos. Lat Am J Aquat Res. 2009; 37(2):199–209. https://doi.org/10.3856/vol37-issue2-fulltext-7
Cortés-Miranda J, Rojas-Hernández N, Muñoz G, Copaja S, Quezada-Romegialli C, Veliz D et al. Biomarker selection depends on gene function and organ: the case of the cytochrome P450 family genes in freshwater fish exposed to chronic pollution. PeerJ. 2024; 12. https://doi.org/10.7717/peerj.16925
Davis AM, Blanchette ML, Pusey BJ, Jardine TD, Pearson RG. Gut content and stable isotope analyses provide complementary understanding of ontogenetic dietary shifts and trophic relationships among fishes in a tropical river. Freshw Biol. 2012; 57(10):2156–72. https://doi.org/10.1111/j.1365-2427.2012.02858.x
Dirección General de Aguas (DGA). Diagnóstico y clasificación de los Cursos y cuerpos de agua según objetivos de calidad. Cuenca del Rio Maipo. Dirección General de Aguas de Chile. 2004.
Doi H, Akamatsu F, González AL. Starvation effects on nitrogen and carbon stable isotopes of animals: An insight from meta-analysis of fasting experiments. R Soc Open Sci. 2017; 4(8):170633. https://doi.org/10.1098/rsos.170633
Doucett R, Booth R, Power G, Mckinley R. Effects of the spawning migration on the nutritional status of anadromous Atlantic salmon (Salmo salar): insights from stable-isotope analysis. Can J Fish Aquat Sci. 1999; 56(11):2172–80. https://doi.org/10.1139/f99-147
Farías M, Charrier R, Carretier S, Martinod J, Fock A, Campbell D et al. Late Miocene high and rapid surface uplift and its erosional response in the Andes of central Chile (33° – 35°S). Tectonics. 2008; 27(1). https://doi.org/10.1029/2006TC002046
Fierro P, Bertrán C, Mercado M, Peña-Cortés F, Tapia J, Hauenstein E et al. Composición del paisaje como determinante de la diversidad y de grupos funcionales alimentarios de macroinvertebrados acuáticos en ríos de la Araucanía, Chile. Lat Am J Aquat Res. 2015; 43(1):186–200. https://doi.org/10.3856/vol43-issue1-fulltext-16
Fierro P, Valdovinos C, Vargas-Chacoff L, Bertrán C, Arismendi I. Macroinvertebrates and fishes as bioindicators of stream water pollution. Water Quality. In Tech. 2017. https://doi.org/10.5772/65084
Figueroa R, Valdovinos C, Araya E, Parra O. Macroinvertebrados bentónicos como indicadores de calidad de agua de ríos del sur de Chile. Rev Chil Hist Nat. 2003; 76(2):275–85. http://dx.doi.org/10.4067/S0716-078X2003000200012
Fry B. Stable isotope ecology. Springer Nature; 2008.
Fu L, Jiang Y, Ding J, Liu Q, Peng QZ, Kang MY. Impacts of land use and environmental factors on macroinvertebrate functional feeding groups in the Dongjiang River basin, southeast China. J Freshw Ecol. 2016; 31(1):21–35. https://doi.org/10.1080/02705060.2015.1017847
Habit E, Bertran C, Arevalo S, Victoriano P. Benthonic fauna of the Itata River and irrigation canals (Chile). Irrig Sci. 1998; 18:91–99. https://doi.org/10.1007/s002710050049
Habit E, Górski K, Vila I, Manosalva A, Díaz G, Toledo B et al. Efectos de las presiones antrópicas sobre la fauna nativa de peces y lampreas de Chile. Gayana. 2024; 88(1):76–104. Available from: https://gayana.cl/index.php/gn/article/view/328/171
Habit E, Victoriano P, Campos H. Ecología trófica y aspectos reproductivos de Trichomycterus areolatus (Pisces, Trichomycteridae)en ambientes lóticos artificiales. Rev Biol Trop. 2005; 53(1–2):195–210.
Hammer DAT, Ryan PD, Hammer Ø, Harper DAT. Past: paleontological statistics software package for education and data analysis. Paleontol Electron. 2001; 4(1):1–09.
Hanson P, Springer M, Ramirez A. Capítulo 1: Introducción a los grupos de macroinvertebrados acuáticos. Revista de biología tropical 2010; 58: 3-37.
Heras H, Garin CF, Pollero RJ. Biochemical composition and energy sources during embryo development and in early juveniles of the snail Pomacea canaliculata (Mollusca: Gastropoda). J Exp Zool. 1998; 280(6):375–83. https://doi.org/10.1002/(SICI)1097-010X(19980415)280:6%3C375::AID-JEZ1%3E3.0.CO;2-K
Hertz E, Trudel M, Cox MK, Mazumder A. Effects of fasting and nutritional restriction on the isotopic ratios of nitrogen and carbon: a meta-analysis. Ecol Evol. 2015; 5(21):4829–39. https://doi.org/10.1002/ece3.1738
Hobson KA, Alisauskas RT, Clark RG. Stable-nitrogen isotope enrichment in avian tissues due to fasting and nutritional stress: implications for isotopic analyses of diet. The Condor. 1993; 95(2):388–94. https://doi.org/10.2307/1369361
Hondolero D, Bluhm BA, Iken K. Caloric content of dominant benthic species from the northern Bering and Chukchi Seas: historical comparisons and the effects of preservation. Polar Biol. 2012; 35(4):637–44. https://doi.org/10.1007/s00300-011-1107-x
Koetsier P, McCauley LMM. An irrigation canal as a lotic mesocosm examining the relationsh. West N Am Nat. 2015; 75(3):259–70. https://doi.org/10.3398/064.075.0303
Kohler SL. Competition and the structure of a benthic stream community. Ecol Monogr. 1992; 62(2):165–88. https://doi.org/10.2307/2937092
Kokotović I, Kolar V, Rožman M, Bočkor L, Vitecek S, Previšić A. Wastewater and warming effects on aquatic invertebrates: experimental insights into multi-level biodiversity consequences. Water Res. 2024; 267:122496. https://doi.org/10.1016/j.watres.2024.122496
Krebs CJ. Niche measures and resource preferences. In: Krebs CJ, editor. Ecological methodology. 3 ed. Vancouver: University of British Columbia; 2014. p.596–653.
Lake JL, Serbst JR, Kuhn A, Smucker NJ, Edwards P, Libby A et al. Use of stable isotopes in benthic organic material as a baseline for estimating fish trophic. Can J Fish Aquat Sci. 2019; 76(7):1227–37. https://doi.org/10.1139/cjfas-2017-0381
Lee KY, Graham L, Spooner DE, Xenopoulos MA. Tracing anthropogenic inputs in stream foods webs with stable carbon and nitrogen isotope systematics along an agricultural gradient. PLoS ONE. 2018; 13(7):e0200312. https://doi.org/10.1371/journal.pone.0200312
Letourneur Y, Fey P, Dierking J, Galzin R, Parravicini V. Challenging trophic position assessments in complex ecosystems: calculation method, choice of baseline, trophic enrichment factors, season and feeding guild do matter: a case study from Marquesas Islands coral reefs. Ecol Evol. 2024; 14(7):e11620. https://doi.org/10.1002/ece3.11620
Minagawa M, Wada E. Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochimica et cosmochimica acta. 48; 1135-1140.
Muñoz Donoso BM. Cambios en la riqueza íctica del rio Maipo. Santiago: Universidad de Chile; 2007. Available from: https://repositorio.uchile.cl/handle/2250/187633
Ndatimana G, Nantege D, Arimoro FO. A review of the application of the macroinvertebrate-based multimetric indices (MMIs) for water quality monitoring in lakes. Environ Sci Pollut Res. 2023; 30(29):73098–115. https://doi.org/10.1007/s11356-023-27559-0
Palma A. Guía para la identificación de invertebrados acuáticos. 1 ed. 2013.
Peig J, Green AJ. New perspectives for estimating body condition from mass/length data: the scaled mass index as an alternative method. Oikos. 2009; 118(12):1883–91. https://doi.org/10.1111/j.1600-0706.2009.17643.x
Philips N. Stream biomonitoring using species traits. Water Atmos. 2004; 12(4):8–09.
Pielou EC. The measurement of diversity in different types of biological collections. J Theor Biol. 1966; 13(C):131–44. https://doi.org/10.1016/0022-5193(66)90013-0
Post DM. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology. 2002; 83(3):703–18. https://doi.org/10.2307/3071875
Qiu L, Ji F, Qiu Y, Xie H, Li G, Shen J. Water-level fluctuationcontrol of the trophic structure of a Yangtze River oxbow. Biology. 2023; 12(10):1359. https://doi.org/10.3390/biology12101359
Quezada-Romegialli C, Jackson AL, Hayden B, Kahilainen KK, Lopes C, Harrod C. tRophicPosition, an r package for the Bayesian estimation of trophic position from consumer stable isotope ratios. Methods Ecol Evol. 2018; 9(6):1592–99. https://doi.org/10.1111/2041-210X.13009
R Development Core Team. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2024. Available from: https://www.r-project.org/
Rau GH, Sweeney RE, Kaplan IR, Mearns AJ, Young DR. Notes and discussions differences in animal 13C, 15N and D abundance between a polluted and an unpolluted coastal site: likely indicators of sewage uptake by a marine food web. Estuar Coast Shelf S. 1981; 13(6):701–07. https://doi.org/10.1016/S0302-3524(81)80051-5
Redding TA, Midlen AB. Estudio de la producción piscícola en los canales de riego. FAO Documento Técnico de Pesca. No 317. Rome. FAO; 1992. Available from: https://openknowledge.fao.org/handle/20.500.14283/t0401s
Santos GS, Piancini LDS, Tincani FH, Klingelfus T, Yamamoto FY, Ribeiro CAO et al. Liver damage in two neotropical fish species from a polluted estuarine area. Ecotoxicol Environ Contam. 2018; 13(1):39–50. https://doi.org/10.5132/eec.2018.01.05
Sayer CA, Fernando E, Jimenez RR, Macfarlane NBW, Rapacciuolo G, Böhm M et al. One-quarter of freshwater fauna threatened with extinction. Nature. 2025; 638(8049):138–45. https://doi.org/10.1038/s41586-024-08375-z
Scharnweber K, Scholz C, Schippenbeil V, Milano S, Hühn D. Effects of mining activities on fish communities and food web dynamics in a lowland river. Ecol Evol. 2024; 14(3):e1111. https://doi.org/10.1002/ece3.11111
Shannon C, Weaver W. A mathematical model of communication. Urbana, IL: University of Illinois Press. 1949; 11:11–20.
da Silva Penha IC, Jardim AA, Prata EG, Seabra LB, Gusmão RR, Ferreira GT et al. Feeding ecology of a detritivorous fish in controlled flood pulse area in the Amazon revealed by stomach content and stable isotopes analyses. Aquat Sci. 2025; 87(19). https://doi.org/10.1007/s00027-024-01147-x
Simaika JP, Samways MJ. Comparative assessment of indices of freshwater habitat conditions using different invertebrate taxon sets. Ecol Indic. 2011; 11(2):370–78. https://doi.org/10.1016/j.ecolind.2010.06.005
Simpson EH. Measurement of diversity. Nature. 1949; 163(4148):688. https://doi.org/10.1038/163688a0
Smith VH, Tilman GD, Nekola JC. Eutrophication: impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environ Pollut. 1999; 100(1–3):179–96.
Smucker NJ, Kuhn A, Cruz-Quinones CJ, Serbst JR, Lake JL. Stable isotopes of algae and macroinvertebrates in streams respond to watershed urbanization, inform management goals, and indicate food web relationships. Ecol Indic. 2018; 90:295–304. https://doi.org/10.1016/j.ecolind.2018.03.024
Soriano Y, Carmona E, Renovell J, Picó Y, Brack W, Krauss M et al. Co-occurrence and spatial distribution of organic micropollutants in surface waters of the River Aconcagua and Maipo basins in Central Chile. Sci Total Environ. 2024; 954:176314. https://doi.org/10.1016/j.scitotenv.2024.176314
Soxhlet F. Die gewichtsanalytische Bestimmung des Milchfettes. Polytech J. 1879; 232:461–65.
Steffy LY, Kilham SS. Elevated δ15N in stream biota in areas with septic tank systems in an urban watershed. Ecol Appl. 2004; 14(3):637–41. https://doi.org/10.1890/03-5148
Stephens RB, Shipley ON, Moll RJ. Meta-analysis and critical review of trophic discrimination factors (∆13C and ∆15N): importance of tissue, trophic level and diet source. Funct Ecol. 2023; 37(9):2535–48. https://doi.org/10.1111/1365-2435.14403
Sumudumali RGI, Jayawardana JMCK. A review of biological monitoring of aquatic ecosystems approaches: with special reference to macroinvertebrates and pesticide pollution. Environ Manage. 2021; 67(2):263–76. https://doi.org/10.1007/s00267-020-01423-0
Theodoropoulos C, Aspridis D, Iliopoulou-Georgudaki J. The influence of land use on freshwater macroinvertebrates in a regulated and temporary Mediterranean river network. Hydrobiologia. 2015; 751(1):201–13. https://doi.org/10.1007/s10750-015-2187-3
Timofeyev MA, Shatilina ZM, Kolesnichenko AV, Bedulina DS, Kolesnichenko VV, Pflugmacher S et al. Natural organic matter (NOM) induces oxidative stress in freshwater amphipods Gammarus lacustris Sars and Gammarus tigrinus (Sexton). Sci Total Environ. 2006; 366(2–3):673–81. https://doi.org/10.1016/j.scitotenv.2006.02.003
Travnichek VH, Maceina MJ. Comparison of flow regulation effects on fish assemblages in shallow and deep water habitats in the Tallapoosa River, Alabama. J Freshw Ecol. 1994; 9(3):207–16. https://doi.org/10.1080/02705060.1994.9664888
Tripp-Valdez A, Arreguin-Sanchez F. The use of stable isotopes and stomach contents to identify dietary components of the spotted rose snapper, Lutjanus guttatus (Steindachner, 1869), off the eastern coast of the Southern Gulf of California. J Fish Aquat Sci. 2009; 4:274–84. https://doi.org/10.3923/jfas.2009.274.284
Tupinambás TH, Callisto M, Santos GB. Benthic macroinvertebrate assemblages structure in two headwater streams, south-eastern Brazil. Rev Bras Zool. 2007; 24(4):887–97. https://doi.org/10.1590/S0101-81752007000400005
Vega-Retter C, Muñoz-Rojas P, Vila I, Copaja S, Véliz D. Genetic effects of living in a highly polluted environment: the case of the silverside Basilichthys microlepidotus (Jenyns) (Teleostei: atherinopsidae) in the Maipo River basin, central Chile. Popul Ecol. 2014; 56(4):569–79. https://doi.org/10.1007/s10144-014-0444-3
Vega-Retter C, Rojas-Hernandez N, Vila I, Espejo R, Loyola DE, Copaja S et al. Differential gene expression revealed with RNA-Seq and parallel genotype selection of the ornithine decarboxylase gene in fish inhabiting polluted areas. Sci Rep. 2018; 8(1):4820. https://doi.org/10.1038/s41598-018-23182-z
Véliz D, Catalán L, Pardo R, Acuña P, Díaz A, Poulin E et al. The genus Basilichthys (Teleostei: Atherinopsidae) revisited along its Chilean distribution range (21° to 40° S) using variation in morphology and mtDNA. Rev Chil Hist Nat. 2012; 85(1):49–59. https://doi.org/10.4067/S0716-078X2012000100004
Veliz D, Rojas-Hernández N, Copaja SV, Vega-Retter C. Temporal changes in gene expression and genotype frequency of the ornithine decarboxylase gene in native silverside Basilichthys microlepidotus: impact of wastewater reduction due to implementation of public policies. Evol Appl. 2020; 13(6):1183–94. https://doi.org/10.1111/eva.13000
Vila I, Veloso A, Schlatter R, Ramírez C. Macrofitas y vertebrados de los sistemas lımnicos de Chile. Santiago: Editorial Universitaria; 2006.
Wilkinson JL, Boxall ABA, Kolpin DW, Leung KMY, Lai RWS, Galban-Malag C et al. Pharmaceutical pollution of the world’s rivers. PNAS. 2022; 119(8):e2113947119. https://doi.org/10.1073/pnas.2113947119
World Water Assessment Programme. Wastewater: the untapped resource: the United Nations world water development report 2017. United Nations Education, Scientific and Cultural Organization; 2017.
Authors

![]() Matías Briones1,
Matías Briones1, ![]() Jorge Cortés-Miranda1,
Jorge Cortés-Miranda1, ![]() Claudio Veloso1,
Claudio Veloso1, ![]() David Véliz1,2 and
David Véliz1,2 and ![]() Caren Vega-Retter1
Caren Vega-Retter1 ![]()
[1] Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Región Metropolitana, Chile. (MB) matias.bioamb@gmail.com, (JCM) jorge.cortes.m@ug.uchile.cl, (CV) clveloso@uchile.cl, (DV) dveliz@uchile.cl, (CVR) carenvega@uchile.cl (corresponding author).
[2] Centro de Ecología y Manejo Sustentable de Islas Oceánicas (ESMOI), Larrondo 1281, Coquimbo, Chile
Authors’ Contribution 

Matías Briones: Formal analysis, Investigation, Methodology, Writing-original draft.
Jorge Cortés-Miranda: Formal analysis, Methodology, Visualization, Writing-review and editing.
Claudio Veloso: Methodology, Resources, Writing-review and editing.
David Véliz: Conceptualization, Resources, Supervision, Writing-original draft.
Caren Vega-Retter: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing-original draft.
Ethical Statement
All the fish and macroinvertebrates sampled were manipulated according to the approved protocols by the Ethics Committee of the Universidad de Chile and complied with the Chilean laws (Resolución Exenta number 3078 Subsecretaria de Pesca).
Competing Interests
The author declares no competing interests.
Data availability statement
Data is available at figshare repository 10.6084/m9.figshare.29318942.
AI statement
ChatGPT was used in this article only for English language editing.
Funding
This study was funded by Fondo Nacional de Desarrollo Científico y Tecnológico – FONDECYT N°11150213 and FONDECYT N°1250534.
Supplementary Material
Supplementary material SUP
How to cite this article
Briones M, Cortés-Miranda J, Veloso C, Véliz D, Vega-Retter C. Effects of pollution and watercourse type on trophic structure: the silverside Basilichthys microlepidotus and its food resources in the Maipo River basin as a model. Neotrop Ichthyol. 2026; 24(2):e250141. https://doi.org/10.1590/1982-0224-2025-0141
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2025 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted February 6, 2026
Accepted February 6, 2026
![]() Submitted August 12, 2025
Submitted August 12, 2025
![]() Epub June 19, 2026
Epub June 19, 2026