![]() Vinícius de Andrade Urbano1
Vinícius de Andrade Urbano1 ![]() ,
, ![]() Driele Delanira-Santos1,2,
Driele Delanira-Santos1,2, ![]() Matheus Maximilian Ratz Scoarize1,2,3 and
Matheus Maximilian Ratz Scoarize1,2,3 and ![]() Evanilde Benedito1,2
Evanilde Benedito1,2
PDF: EN XML: EN | Supplementary: S1 S2 | Cite this article
Abstract
Landscape change caused by hydropower dam construction and invasion of agriculture in the riparian zone has been a catalyst for alterations in fish food web dynamics. The objectives of this study were to investigate the trophic state and land use of three rivers, one directly impacted by a dam; the trophic structure of four fish species of four trophic guilds; and the relationship between the trophic state and land use with trophic structure. We found a greater total allochthonous contribution in the dam-regulated river in contrast to its tributaries, along with less natural cover in the riparian zone, but a high percentage of natural vegetation on the islands. Additionally, we observed greater contribution of the riverine sources for all three rivers, independent of the trophic state and natural cover of the river. The trophic position was greater for the fish in the river in a protected area and lower in the rivers with a high percentage of agropastoral activities in the riparian areas. These findings show the importance of the conservation of the riverine zones mainly in dam-regulated rivers, because this vegetation provides the main energy source by fish.
Keywords: Land use change, Oligotrophication, Stable isotopes, Wetland.
Mudanças na paisagem causadas pela construção de barragens hidrelétricas e invasão da agricultura na região ribeirinha têm catalisado alterações na dinâmica da cadeia alimentar de peixes. Os objetivos deste estudo foram investigar o estado trófico e o uso da terra de três rios, um diretamente impactado por uma barragem; a estrutura trófica de quatro espécies de peixes de quatro guildas tróficas; e a relação entre o estado trófico e o uso da terra com a estrutura trófica. Encontramos maior contribuição alóctone total no rio regulado pela barragem em contraste com seus tributários, juntamente com menos cobertura natural na região ribeirinha, mas uma alta porcentagem de vegetação natural nas ilhas. Além disso, observamos uma maior contribuição das fontes ribeirinhas para todos os três rios, independentemente do estado trófico e da cobertura natural do rio. A posição trófica foi maior para os peixes no rio em uma área protegida e menor nos rios com uma alta porcentagem de atividades agropastoris nas áreas ribeirinhas. Esses achados mostram a importância da conservação das zonas ribeirinhas, principalmente em rios regulados por barragens, pois essa vegetação fornece a principal fonte de energia dos peixes.
Palavras-chave: Áreas úmidas, Mudanças de uso da terra,Isótopo estável, Oligotrofização.
Introduction
Floodplain systems present some of the most complex ecosystem dynamics (Petsch et al., 2023), with high heterogeneity in their chemical and ecological characteristics (Rocha, 2011; Tessler et al., 2017). These systems thereby provide numerous ecosystem services to the environment and society (Petsch et al., 2023). These ecosystems are dependent on the surrounding terrestrial vegetation due to periodic flooding and the subsequent input of allochthonous resources, a dynamic known as the flood pulse (Junk et al., 1989). However, in the last decades, these environments have lost their natural features due to river regulation by hydropower dams implementation (Petsch et al., 2023).
The construction of hydropower dams has been identified as one of the biggest threats to the biodiversity of rivers (Hoeinghaus et al., 2009; Murphy et al., 2019) and wetlands (Agostinho et al., 2008). These activities lead to fragmentation and consequent loss of natural habitats through land use change threatening freshwater biodiversity throughout the world (Isbell et al., 2023). The establishment of hydropower dams also blocks the migratory routes of commercially important fish species (Agostinho et al., 2007). As also, alters the flood and dry regimes in floodplains (Agostinho et al., 2008; Okawa, 2010; Costa et al., 2012) due to the regulation of hydrological levels (Agostinho et al., 2007) disrupting the natural flood pulse (Junk et al., 1989). Additionally, hydropower dams retain organic matter and nutrients (Agostinho et al., 2008) and decrease water turbidity downstream (Agostinho et al., 2008), resulting in a more oligotrophic river environment (Duda et al., 2011).
The retention of nutrients and organic matter by dam cascades and the reduction or absence of periodic flooding in floodplains can lead to a potential decrease in available energy sources. The retention of nutrients by the dam limits autochthonous primary production (Agostinho et al., 2008). Simultaneously, the absence of the flood pulse causes a decline in the contribution of allochthonous organic matter (Junk et al., 1989). Consequently, there may be a reduction in fish species richness, abundance, and biomass (Lobón-Cerviá et al., 2015; Freitas et al., 2018; Arantes et al., 2019), which has economic impacts on families relying on commercial and subsistence fishing (Oczkowski et al., 2009).
In environments with low primary autochthonous production, consumers tend to depend more on allochthonous energy sources (McWilliam-Hughes et al., 2009). Riparian vegetation, in particular, serves as a significant source of energy in the food webs of oligotrophic environments (Correa, Winemiller, 2018). Zheng et al. (2018) found a high contribution of allochthonous energy sources to fisheries biomass in downstream environments affected by dams, in contrast to environments without such impacts. Therefore, it is essential to consider land-use practices around aquatic environments, as the contribution of allochthonous energy sources tends to decline as the natural land cover decreases (Wang et al., 2014).
Land use change is the main driver of biodiversity loss (Isbell et al., 2023), particularly the expansion of agricultural and pastoral activities, which resulted in a decrease in natural land cover (Foley et al., 2005). As a result, there is a decline in consumer biodiversity given that these organisms rely on natural land cover for their habitat (Wang et al., 2014), especially in floodplain areas (Junk et al., 1989). Consumers influenced by anthropogenic activities often exhibit shifts in their trophic positions compared with those in undisturbed environments (Carvalho et al., 2015). Conserved environments, characterized by high resource availability, play an important role in sustaining complex food webs (Durán et al., 2016), in contrast to environments impacted by human activities (Xiang et al., 2023). Therefore, understanding the trophic dynamics of consumers under the influence of hydropower and land-use impacts, particularly in conserved environments, is crucial for the development of effective mitigation measures to address these impacts.
Studies have showed the importance of allochthonous energy sources for consumers in natural oligotrophic rivers (e.g., Correa, Winemiller, 2018; Zheng et al., 2018; Carvalho et al., 2023), especially in environments with high availability of allochthonous energy sources (Correa, Winemiller, 2018). However, little is known about the dynamics of the allochthonous energy source to artificially oligotrophic rivers. To identify the trophic structure of aquatic environments, carbon and nitrogen stable isotope analyses are useful (McWilliam-Hughes et al., 2009). The δ13C values in consumers are typically similar to their energy sources (Fry, 2006), making them a good differentiator between different energy sources (Manetta, Benedito-Cecilio, 2003). On the other hand, the δ15N values exhibit a discrimination of approximately 3.4% per trophic level (Post, 2002), and thus provide valuable information for estimating trophic positions (McCutchan Jr. et al., 2003).
In this study, we aimed (i) to investigate the trophic status and land use in the riparian vegetation areas of three rivers within a Neotropical floodplain, including one river directly affected by a dam cascade; (ii) to analyze the contribution of autochthonous and allochthonous carbon sources and the trophic positions of consumers (four trophic guilds of fish) in each of the three environments; (iii) to determine if there are differences in the contribution of riverine and pelagic energy sources for consumers; and (iv) to examine the relationships between the trophic status of the rivers and land use with energy sources as well as trophic positions. We tested two hypotheses (i) there is a greater contribution from allochthonous sources for a river affected by the dam cascade, indicating a higher reliance on riverine regions for sustenance due to a decrease of autochthonous primary production compared with other rivers; and (ii) fish in a dam-regulated river have lower trophic position due to a decrease in biodiversity caused by dam regulation, decrease in trophic links, compared with rivers without this impact.
Material and methods
Study area. The upper Paraná River floodplain (PIAP) is situated within the biodiversity hotspot recognized as the Atlantic Forest (Ribeiro et al., 2009) and forms part of the La Plata River basin, which ranks as the fifth largest river system globally (Pochat, 2011). The PIAP encompasses one river impacted by a dam cascade: Paraná River (I-01), and two control rivers (i.e., free of dam regulation): Ivinhema River (C-01) and Baía River (C-02). The I-01 experienced oligotrophication (Agostinho et al., 2008) subsequent to the construction of the Porto Primavera Dam in 1998 (Agostinho et al., 2007; Torrecilha, 2008), which significantly altered the river dynamics, leading to a decline in sedentary and migratory fish species (Agostinho et al., 2007). This dam is 10,186.20 m long, resulting in the flooding of an area of 2,250 km² (equivalent to or 225,000 ha) (CESP, 2024). This dam is preceded by numerous other large dams upstream (Agostinho et al., 2004). In addition to the impacts caused by the cascade of reservoirs, anthropogenic influences from urban and rural areas along the river’s banks further exacerbate the situation (Agostinho et al., 2007). However, its islands show a high conservation level due to their status as a protected area for sustainable use: an Environmental Protected Area (EPA) of the I-01 islands and floodplain. This designation allows most commercial uses within its borders (i.e., it is not as restrictive as the national parks), but it safeguards the islands in the three main rivers of the wetlands.
The C-02 experiences indirect impacts from urban areas, leading to the release of pollutants into its tributaries, along with surface runoff from agricultural activities in their watershed (Pott et al., 2014). Additionally, this river is partially obstructed by the Porto Primavera Dam (Granzotti et al., 2018). Conversely, the C-01 benefits from partial protection through the establishment of the Parque Estadual das Várzeas do Rio Ivinhema (Rio Ivinhema State Park), created in 1998 as a compensatory measure for the implementation of the Porto Primavera Dam (Torrecilha, 2008). This area boasts conservation (Carvalho, 2019) and represents the least affected stretch of the Porto Primavera Dam’s influence on the I-01 (Ragonha et al., 2014) because it is not obstructed by the dam (Granzotti et al., 2018).
In each environment, sampling was collected from the I-01, C-01 and C-02 as well as the lagoons connected to the rivers (see Tab. S1). In the I-01, the sampling occurred at 22°45’39.96”S 53°15’7.44”W, the Pau Véio blind channel (22°44’50.76”S 53°15’11.16”W), and the Garças Lagoon (22°43’27.18”S 53°13’04.56”W). In the Ivinhema River, the sampling occurred at 22°47’59.64”S 53°32’21.3”W and the Patos Lagoon (22°49’33.66”S 53°33’09.9”W). In the C-02, the sampling occurred at 22°43’23.16”S 53°17’25.5”W and the Guaraná Lagoon (22°43’16.68”S 53°18’09.24”W) (Fig. 1).

FIGURE 1| Study area. I-01: P01, Paraná River; P02, Garças Lagoon; and P03, Pau Véio blind channel. C-02: B01, Baía River and B02, Guaraná Lagoon. C-01: I01, Ivinhema River and I02, Patos Lagoon.
Land use. The Land use assessment of the riparian vegetation area followed the guidelines outlined in the Brazilian Forest Code (Federal Law n. 12.651/2012), the primary legislation in Brazil that protects riparian vegetation. This code establishes specific dimensions for permanent preservation areas (PPAs) along riverbanks, which serve as buffers for riparian vegetation (Da Cruz et al., 2022). Accordingly, the assessment considered a 100-m buffer for rivers with a channel width of up to 200 m (in this study, the C-01 and C-02), and a 500-m buffer for rivers with a width exceeding 600 m (in this study, the I-01) (Brasil, 2012). The land-use assessment was conducted by using the Quantum Geographic Information System (QGIS). Polygons were created to cover each river and 100-m buffers for the C-01 and C-02 and 500-m buffers for the I-01 in the last lentic system between the Porto Primavera Dam and the former Sete Quedas Fall. Subsequently, a polygon also covered all islands of the I-01 islands and floodplain. High-resolution (30 × 30 m) land-use images were obtained from MapBiomas (Souza et al., 2020; https://mapbiomas.org/en). These images were then reclassified into the following land-use categories: forest, wetlands, agriculture, pasture, mosaic (a combination of pasture and agriculture), and urban areas. The land-use map obtained from MapBiomas was overlaid with the previously created buffers using the “cut out the mask layer” function, and the percentage of land use within each buffer was calculated using the “Landscape Ecology” tool.
Sampling. The sampling was conducted quarterly between July 2009 and August 2010 as part of the project “A Planície de Inundação do alto rio Paraná,” a long-term ecological research (LTER) initiative (PELD/CNPq nº 441356/2020-6). Specimens of four trophic guilds were collected: the herbivorous characin, Schizodon borellii (Boulenger, 1900); the detritivorous Curimbata, Prochilodus lineatus (Valenciennes, 1837); the piscivorous spotted sorubim, Pseudoplatystoma corruscans (Spix & Agassiz, 1829); and the omnivore Granulated catfish, Pterodoras granulosus (Valenciennes, 1821). The fish were identified based on the exemplars deposited in the Fish Collection of Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (Nupélia) of the Universidade Estadual de Maringá (UEM), Maringá (NUPE 2010030801, NUPE 91073001, NUPE 2009091703, and NUPE 2005110801, respectively). Each specimen was collected, and its potential energy sources were investigated, including C3 and C4 aquatic macrophytes, phytoplankton, biofilm, and riparian vegetation. The energy sources were initially categorized into autochthonous sources (aquatic macrophytes, phytoplankton, and biofilm) and allochthonous sources (riparian vegetation), and subsequently classified into riverine sources (riparian vegetation, aquatic macrophytes, and biofilm) and pelagic sources (phytoplankton). This classification is based on Thorp, Delong (1994) used in riverine productivity model, where they classified all the energy sources produced in allochthonous area (i.e., riparian vegetation) and those produced in the shore region of the aquatic environments (i.e., macrophytes, periphyton and particulate organic matter) as riverine sources. While pelagic sources are the energy sources produced in the pelagic region of the river (Thorp, Delong, 1994).
The primary consumer, S. borellii, exhibits an herbivorous feeding habit (Ferretti, 1996; Hahn et al., 2002); it primarily consumes terrestrial vegetation and aquatic macrophytes (Ferretti, 1996). P. lineatus is classified as a detritivore species, mainly feeding on debris (Fugi et al., 1996), while P. granulosus is an omnivorous fish that consumes both autochthonous sources such as fish and plants, as well as allochthonous sources including vegetation and insects (Agostinho et al., 2003, 2009a). Pseudoplatystoma corruscans, a piscivore top predator (Hahn et al., 2002), is considered an umbrella species (Agostinho et al., 2007) and is currently at risk of extinction (ICMBio, 2018). It is a migratory fish, and its reproduction depends on flood periods (Agostinho et al., 2004). It utilizes lagoons as nursery habitats and relies on flooding to connect the river to the lagoons (Agostinho et al., 2007). Therefore, dam impacts have had a negative effect on the life cycle of this fish (Agostinho et al., 2008). Among the main impacts on fish, the suppression of natural barriers stands out, allowing the invasion of exotic species and the alteration of the limnological conditions of modified ecosystems (Ruaro et al., 2020). These impacts influence the body condition of many species due to the effect of dam operations on the flood pulse, which promotes seasonal entry of food resources to native species (Espínola et al., 2012).
Fish were collected using stationary nets with different mesh sizes (2–12 cm between the nodes), which were deployed for a 24 h period and checked every 8 h. Following the sampling, the fish were transported and euthanized in accordance with the American Veterinary Medical Association (AVMA) guidelines (Underwood et al., 2013). Only adult fish were used in this study to minimize ontogenetic effects on isotopic analyses. From each individual, a muscle sample (approximately 2 cm2) was extracted near the base of the dorsal fin insertion. To estimate the contribution of energy sources, five samples of the most abundant species of riparian vegetation and aquatic macrophytes were collected from each study site. Biofilm samples were obtained by gently scraping the petioles of the aquatic macrophyte Pontederia azurea Sw. using a blade wrapped in aluminum foil. Particulate organic matter (POM) samples were collected by obtaining 500 ml of surface water from the rivers using polyethylene containers, and, phytoplankton was gathered with a 53-μm mesh size plankton net. Samples were allowed to settle and the sedimentary bodies were subsequently removed from the samples to spearing phytoplankton to debris as employed by Santana et al. (2009), Alves et al. (2017), and Urbano et al. (2022). Both phytoplankton and periphyton samples were filtered through glass fiber filters (Whatman GFC) with a 47 mm diameter, which had been pre-burned at 400ºC for 4 h.
Turbidity and water transparency (measured in meters) were assessed using a portable potentiometer and a Secchi disc, respectively. In the Basic Limnology Laboratory (Laboratório de Limnologia Básica) of the Research Center for Limnology, Ichthyology, and Aquaculture (Nupelia/UEM), water samples from the sampling points were analyzed for the concentrations of total nitrogen (TN), total phosphorus (TP), nitrate ions (NO3–), ammonium ions (NH4+), phosphate ions (PO43-), and chlorophyll a (Chla). Water samples were stored on ice and transported to the nearest field station, where they were filtered through GF 52-C membranes within 10 hours of sampling and immediately frozen at -20°C for subsequent analyses of dissolved nutrients. Water was also frozen at -20°C prior to filtering to measure total nitrogen and total phosphorus. The persulfate method was employed to analyze total nitrogen, converting all nitrogenous compounds to nitrate (Reis, Zagatto, 1978). Using a flow-injection system, the ion was quantified with a spectrophotometer after reducing nitrite in the presence of cadmium (Giné et al., 1980). The indophenol blue method was used to quantify ammonium, and the readings were taken with a spectrophotometer (Mackereth et al., 1978). According to the method outlined by Golterman et al. (1978) total and reactive dissolved phosphorus (P) were measured with a spectrophotometer. The membranes were utilized for chlorophyll quantification, employing the extraction method with acetone and subsequent spectrophotometric reading, as described by Golterman et al. (1978).
The samples were dried at 60°C for 72 h (Lopes et al., 2009). Afterward, the samples were ground into a fine and homogeneous powder using a mill. The powdered samples were sent to the University of California Davis Stable Isotope Facility (Davis, CA, USA) for the determination of carbon and nitrogen isotopic values. The Vienna PeeDee Belemnite (VPDB) was used as the standard reference for δ13C, while atmospheric δ15N was used as the standard reference for nitrogen. The analyses were conducted using a mass spectrophotometer, following the method described by Lajtha, Michener (1994):
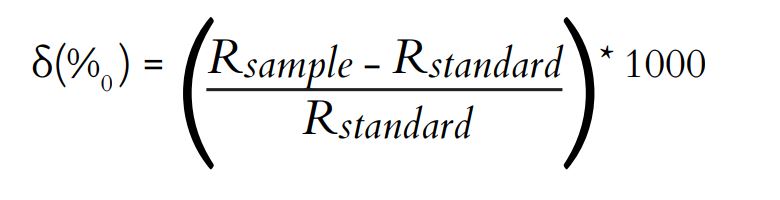
where R is 13C: 12C or 15N: 14N.
Data analysis. The trophic state was determined by using the formula proposed by Lamparellii (2004), which utilizes the annual mean concentrations of Chl a and TP. The equation is as follows:

where TSI(Chla) is the chlorophyll trophic state index, TSI(P) is the phosphor trophic state index, and TSI is the trophic state index. The results are classified as followed: ultraoligotrophic for TSI ≤ 47, oligotrophic for 47 < TSI ≤ 52, mesotrophic for 52 < TSI ≤ 59, eutrophic for 59 < TSI ≤ 63, supereutrophic for 63 < TSI ≤ 67, and hypereutrophic for TSI > 67 (Lamparellii, 2004).
To differentiate the rivers in terms of the abiotic variables, a principal component analysis (PCA) was conducted with the vegan package (Oksanen et al., 2019) in R (R Development Core Team, 2023) using the dissimilarity matrix and Euclidean distances (Legendre, Legendre, 1998). To identify the differences in abiotic variation and the abiotic composition between the rivers, a permutational multivariate analysis of variance (PERMANOVA; Anderson, 2001) was performed to evaluate the difference in the abiotic composition of rivers using the vegan package (Oksanen et al., 2019) in R (R Development Core Team, 2023). Because the abiotic variables were not normally distributed, they were subjected to a Kruskal-Wallis test to determine differences between the rivers.
A total of 344 samples were collected between June 2009 and June 2010, comprising 162 fish samples and 182 primary producer samples. Isotopic values can vary with time (Fry, 2006). Therefore, we performed a PERMANOVA to assess whether there was isotopic variation between the individuals of each population due to collection period. For each of the four species from each of the three rivers we performed a separate PERMANOVA using the δ13C and δ15N values, followed by Bonferroni adjustment of the p value (i.e., 0.05/5). Given no significant differences were found, we grouped individuals collected between June 2009 and June 2010 by species according to the river they were sampled. If there was a significant interaction in the PERMANOVA, pairwise comparisons were performed by using the pairwise.adonis function in the vegan package in R (R Development Core Team, 2023). The same was performed with values of δ13C and δ15N of each basal energy source (i.e., aquatic macrophytes, periphyton, phytoplankton, POM and riparian vegetation) collected in different months between June 2009 and June 2010 in each river or energy sources collected in different environments from each river (i.e., river channel or lagoons connected to the rivers). There were no significant differences in δ13C and δ15N values of each fish species or of each basal energy source (collected in different months), and they were grouped to conduct stable isotope analyses (i.e., trophic position calculation and energy source analyses) (see Tab. S1).
To evaluate whether the data source could explain the consumer data, simulated mixing polygons (Smith et al., 2013) were run using various trophic discrimination factors (TDF) to identify the best TDF for each consumer species in each environment. For S. borellii, the TDF for herbivore fish were tested (Canseco et al., 2021); for P. lineatus, the species-specific TDF were tested (Sacramento et al., 2016); for P. granulosus, the TDF for omnivore fish were tested (McCucthan et al., 2003); and for P. corruscans, the species-specific TDF (Manetta et al., 2023) and the piscivore fish TDF (Cansenco et al., 2021) were tested. In addition, the universal TDF proposed by Post (2002), Cansenco et al. (2021), Vanderklift, Ponsard (2003), and Caut et al. (2009) were tested for each consumer. The TDF that explained best the data were those proposed by Post (2002). After identifying the best TDF for the data, the simulated mixing polygons (Smith et al., 2013) were run again, and some samples of P. lineatus collected in December from C-01 and March in C-02 and, one consumer of I-01 was an outlier (see Tab. S1), these values could not be explained by the energy source data and thus were removed following the criteria suggested by Smith et al. (2013) (see the plot of simulated mixing polygons in Fig. S2).
First, trophic position was calculated using the mean and standard deviation of all baselines, incorporating the TDF proposed by Post (2002) of 3.42 for δ15N. This initial TP was used to correct the TDF used in the energy source analyzes (see below). To estimate TP with greater precision, we use the weighted average of δ15N from allochthonous and autochthonous energy sources according to the contribution of these energy sources to each consumer in each environment, generating specific baseline values of average δ15N for each consumer in each environment, according to the formula from Post (2002):

where: TP = trophic position; δ15Nconsumer = value of δ15N to each consumer; δ15Nbaseline1 = mean of δ15N to allochthonous energy source; δ15Nbaseline2 = mean of δ15N to autochthonous energy source; α = proportion of the contribution of allochthonous energy source; 1 – α = proportion of the contribution of autochthonous energy source.
To determine the proportion of energy sources contributing to consumer biomass, the stable isotope mixing model was applied using the SIMMr package (Parnell et al., 2013) in R (R Core Team 2023). The model incorporated the δ15N and δ13C values of consumers and energy sources, as well as the TDF adjusted based on the calculated trophic position (calculated using the mean of δ15N to all energy source) for each consumer in each environment. SIMMr utilizes the Just Another Gibbs Sampler (JAGS) program to execute the Bayesian stable isotope mixing model. The model is run using the simmr_out and sim_mcmc functions; it employs Markov chain Monte Carlo (MCMC) for stochastic simulation to generate numerous random assumptions of energy source contributions to determine the best proportion of contributions for the given data. Subsequently, the compare_sources function (Parnell et al., 2013) was utilized to assess the probability of the sum of all autochthonous sources (i.e., the sum of contribution of phytoplankton, periphyton, POM and C3 and C4 aquatic macrophytes) contributing more than allochthonous sources (i.e., Riparian vegetation) and the probability of the sum of all riverine sources (i.e., the sum of contribution of riparian vegetation, periphyton, POM and C3 and C4 aquatic macrophytes) contributing more than pelagic sources (i.e., Phytoplankton).
To assess whether there were differences in the allochthonous contribution among the rivers or differences in the riverine contribution and difference in the TP, a two-way analysis of variance (ANOVA) or the Kruskal-Wallis test was conducted, depending on whether the data fulfilled the normality and homoscedasticity assumptions based on the Shapiro-Wilk test and Levene’s test, respectively. The ANOVA or Kruskal-Wallis was conducted using the allochthonous contribution to all consumers in each river and the values calculated of TP to each consumer of each species between each river. Furthermore, to determine whether there were differences in the contribution of autochthonous and allochthonous sources as well as riverine and pelagic sources within each environment, a t-test was performed for each environment (assuming that the data met the normality and homoscedasticity based on the Shapiro-Wilk test and Levene’s test, respectively), using the value of contribution of autochthonous and allochthonous or riverine and pelagic to each consumer in each river.
Results
There is a high percentage of native vegetation (forest and wetlands) along the margins of the I-01 islands (96%), as well as along the margins of the C-01 (93%) and C-02 (88%) rivers, in contrast to the I-01 (75%). Along the I-01, there is a significant presence of agropastoral activities (e.g., agriculture and pasture) in its margins, in contrast to the C-01 and C-02 (Tab. 1).
TABLE 1 | Percentage (%) land use of the riparian forest area along the margins of the I-01, C-01 and C-02 and on the I-01 island. I-01 = Paraná River, C-01 = Ivinhema River, C-02 = Baía River.
| Forest | Wetland | Pasture | Mosaic | Urban | Agriculture |
C-01 | 20.6 | 72.1 | 3.9 | 2.5 | 0.0 | 1.0 |
C-02 | 3.0 | 85.2 | 2.3 | 9.2 | 0.0 | <0.1 |
I-01 | 17.1 | 56.7 | 16.4 | 6.7 | 0.8 | 2.3 |
I-01 Island | 14.0 | 82.4 | 0.3 | 3.0 | 0.8 | 0.1 |
The first two axes of the PCA explained 65.4% of the abiotic variation. The first axis separated the I-01 from the other two rivers; the I-01 was associated with Secchi disk, alkalinity, and NO3–, while the other rivers were associated with nutrients, Chla, and turbidity. The PERMANOVA revealed a significant difference in abiotic composition between rivers (df = 2, F = 7.28, p = 0.002, R2 = 0.43) (Fig. 2). We observed lower nutrient concentrations, turbidity, and Chla in the I-01 compared with the C-01 and C-02. Conversely, the alkalinity and transparency were higher in the I-01 compared with the C-01 and C-02. Additionally, we found that the I-01 is ultraoligotrophic, the C-01 is mesotrophic, and the C-02 is eutrophic (Tab. 2).

FIGURE 2| Principal component analysis of the abiotic variables with ellipses indicating a 95% credibility interval. Secchi: Secchi disk (m); Turb.: turbidity (NTU); Alc: alkalinity; Chl: chlorophyll a (µg/L); TN: total nitrogen (mg/L); NO3-: nitrate ion (mg/L); TP: total phosphorus (µg/L); NH4+: ammonium ion (µg/L); PO3-4: phosphate ion (µg/L).
TABLE 2 | Abiotic variables from I-01 (Paraná River), C-01 (Ivinhema River) and C-02 (Baía River), sampled in February, June, September, and December 2009. Alk: alkalinity (mg L-1), Chl.: chlorophyll a (mg L-1), E: eutrophic, M: mesotrophic, NH4+: ammonium ion (µg L-1), NO3–: nitrate ion (mg L-1), PO43-: phosphate ion (mg L-1), TN: total nitrogen (mg L-1), TP: total phosphorus (mg L-1), Trans.: transparency (m), TS: trophic state; TSI: trophic state index, Turb.: turbidity (NTU), U: ultraoligotrophic.
River | TS | TSI | Chl. | TP | TN | Trans. | Turb. | NH4+ | NO3– | PO43- | PH | DO | ºC | Alk. |
I-01 | U | 44 | 0.4 | 10.1 | 815.0 | 5.7 | 1.1 | 6.9 | 163.9 | 5.1 | 7.1 | 7.5 | 25.6 | 459.1 |
C-01 | M | 53 | 1.3 | 44.2 | 921.5 | 0.7 | 22.1 | 6.1 | 125.0 | 17.0 | 6.7 | 6.2 | 25.1 | 411.4 |
C-02 | E | 60 | 3.8 | 42.7 | 1145.5 | 1.0 | 5.5 | 24.4 | 8.4 | 10.3 | 6.2 | 5.3 | 26.6 | 214.3 |
We observed the lowest mean TP for the herbivore (I-01: 2.20 ± 0.67, C-01: 1.79 ± 0.69 and C-02: 1.73 ± 0.59) and detritivore (I-01: 2.16 ± 0.69, C-01: 2.35 ± 0.71 and C-02: 1.93 ± 0.41) consumers, and highest mean TP for the omnivore (I-01: 2.74 ± 0.74, C-01: 2.69 ± 0.59 and C-02: 1.96 ± 0.74) and piscivore (I-01: 2.75 ± 0.29, C-01: 2.79 ± 0.64 and C-02: 2.22 ± 0.31) consumers in all rivers (Tab. 3). For the C-01, we found a significant difference in the TP for the herbivore consumers compared with the TP for the detritivore, omnivore and piscivore consumers. When comparing the environments, we found the highest TP in the C-01 (range 1.79–2.79) and I-01 (range 2.15–2.75) followed by the C-02 (range 1.73–2.22) (Figs. 3–4).
TABLE 3 | Mean ± standard deviation of the consumer and producer δ13C and δ15N values at each site. Herbivore: Schizodon borellii; detritivore: Prochilodus lineatus; omnivore: Pterodoras granulosus, and piscivore: Pseudoplatystoma corruscans. n = number of individuals. I-01 = Paraná River, C-01 = Ivinhema River, C-02 = Baía River.
|
| I-01 | C-01 | C-02 | |||||
| n | δ13C | δ15N | n | δ13C | δ15N | n | δ13C | δ15N |
Herbivore | 21 | -28.14±3.65 | 9.71±2.35 | 17 | -26.03±3.46 | 9.18±2.35 | 18 | -25.59±3.74 | 8.72±2.15 |
Detritivore | 7 | -27.00±2.26 | 10.09±2.34 | 6 | -29.40±3.19 | 9.22±0.66 | 13 | -27.93±4.23 | 9.30±1.44 |
Omnivore | 13 | -25.79±1.85 | 13.23±1.27 | 9 | -27.30±1.66 | 11.95±1.75 | 5 | -27.17±1.87 | 10.06±2.51 |
Piscivore | 13 | -27.31±1.08 | 11.37±1.00 | 11 | -26.56±2.92 | 11.89±2.20 | 19 | -27.55±2.00 | 9.91±1.03 |
Phytoplankton | 8 | -25.13±1.15 | 4.09±2.78 | 8 | -25.99±0.56 | 3.50±0.96 | 7 | -28.56±0.53 | 2.22±0.75 |
Biofilm | 4 | -15.41±0.77 | 6.45±0.53 | 8 | -26.40±0.86 | 4.23±1.70 | 8 | -27.53±2.61 | 3.74±1.87 |
C3 A. Macrophytes | 12 | -25.74±2.49 | 6.88±0.64 | 54 | -29.29±0.80 | 4.93±2.61 | 39 | -28.01±1.57 | 8.22±3.04 |
C4 A. Macrophytes | 4 | -12.29±0.11 | 10.86±0.10 | 4 | -10.83±0.13 | 6.94±0.43 | 4 | -11.26±0.33 | 12.29±0.65 |
R. Vegetation | 12 | -31.73±1.35 | 4.10±1.34 | 26 | -29.87±1.26 | 2.92±2.04 | 8 | -28.89±1.14 | 4.14±2.72 |

FIGURE 3| Biplot with the mean and standard deviation of the δ13C and δ15N values for the energy sources: AM = aquatic macrophytes, BF = biofilm, PH = phytoplankton, and RV = riparian vegetation. The δ13C and δ15N values are for the consumers at each site. C-01 = Ivinhema River, C-02 = Baía River, I-01 = Paraná River.

FIGURE 4| Trophic position for each fish species in each studied river. Consumers in the same environment with the same letters (a or b) indicate the lack of a significant difference. Consumers of the same species with the same symbol (* or #) indicate the lack of a significant difference. The black point in the center of each violin is the mean of the trophic position. I-01 = Paraná River, C-01 = Ivinhema River, C-02 = Baía River.
For the I-01, we observed a greater total contribution from riparian vegetation (40.7%) compared with the C-01 (29.0%) and the C-02 (20.8%). The C-02 had a higher phytoplankton contribution (24–50%) than other energy sources. For the C-01, we found a higher contribution from both riparian vegetation and phytoplankton (19–35%) than from other energy sources. For all rivers, we observed a low contribution from aquatic macrophyte C3 and C4 and biofilm (3–26%) (Figs. 5–6, Tab. S3).

FIGURE 5| Proportion of the contribution of the energy sources for each fish species at each site. AM C3 = aquatic macrophyte C3, AM C4 = aquatic macrophyte C4, BF = biofilm, PH = phytoplanktonand RV = riparian vegetation. C-01 = Ivinhema River, C-02 = Baía River, I-01 = Paraná River.

FIGURE 6| The t-test results for the autochthonous and allochthonous contributions as well as the pelagic and riverine contributions at each site. A significant pairwise interaction is represented by the subscript letters (ab). I-01 = Paraná River, C-01 = Ivinhema River, C-02 = Baía River.
The contribution of autochthonous energy sources in the C-01 and C-02 was greater than the contribution of allochthonous energy sources. However, contributions among autochthonous and allochthonous energy sources were not different for the I-01 (Fig. 6). We observed a relatively high probability that the riverine contribution is greater than the pelagic contribution, as well as a relatively high probability of a greater contribution from autochthonous sources compared with allochthonous sources (Fig. 7; Tab. 4). Furthermore, the contribution of riparian vegetation and riverine energy sources for each fish species was not significantly different between the subsystem (Gl = 2, F = 3.71, p = 0.06; and Gl = 2, F = 1.06 and p = 0.38, respectively), but the mean contribution of allochthonous sources to all fish in the I-01 was two times greater than the allochthonous contribution in the C-02 and 1.4 times greater than the allochthonous contribution in the C-01 (Fig. 5).

FIGURE 7| The arrow width is proportional to the contribution of the energy sources (phytoplankton, riparian vegetation, and aquatic macrophytes and biofilm) to the mesotrophic C-01, the eutrophic C-02, and the ultraoligotrophic I-01. AM = aquatic macrophytes, PH = phytoplankton, PP = periphyton, and RV = riparian vegetation. C-01 = Ivinhema River, C-02 = Baía River, I-01 = Paraná River.
TABLE 4 | Results of the SIMMr comparison, showing the probability (%) that the riverine source will be greater than the pelagic source (riverine × pelagic) and the probability (%) that the autochthonous contribution will be greater than the allochthonous contribution (autochthonous × allochthonous) for each fish species at each site Herbivore: Schizodon borellii; detritivore: Prochilodus lineatus; omnivore: Pterodoras granulosus, and piscivore: Pseudoplatystoma corruscans. I-01 = Paraná River, C-01 = Ivinhema River, C-02 = Baía River.
|
| Herbivore | Detritivore | Omnivore | Piscivore |
Riverine x Pelagic | I-01 | 33 | 86 | 76 | 89 |
C-01 | 45 | 95 | 86 | 64 | |
C-02 | 28 | 18 | 77 | 23 | |
Autochthonous x Allochthonous | I-01 | 99 | 62 | 92 | 84 |
C-01 | 98 | 55 | 96 | 96 | |
C-02 | 98 | 97 | 90 | 99 |
Discussion
We partially accepted our first hypothesis that the river impacted by dam has a greater contribution of allochthonous carbon sources and that the riverine region is the main carbon source in both rivers: While the allochthonous carbon source contribution was not different between the rivers, the mean of the allochthonous carbon source contribution was higher for the I-01. This finding highlights the importance of natural vegetation for sustaining fish populations and emphasizes the need for conservation efforts in the riverine regions of these rivers. However, we rejected our second hypothesis that the fish in the river impacted by dam would occupy a lower trophic position; our results showed that the C-02 presented the lowest trophic position. A lower trophic position in C-02 (a eutrophic environment) may be attributed to the impacts of land use in its watershed (see Tab. 1), as observed in other studies (Cabana, Rasmussen, 1996; Carvalho et al., 2015).
The greater contribution of allochthonous sources of energy in the I-01 (ultraoligotrophic river) relative to the C-01 and the C-02 (mesotrophic and eutrophic rivers, respectively) highlights the importance of riparian forest as a carbon source for sustaining oligotrophic environments, as observed in previous studies (Correa, Winemiller, 2018; Champagne et al., 2022). Despite the greater allochthonous contribution in the ultraoligotrophic I-01, the environment along the margins of this river had approximately 70% natural cover, whereas the C-01 and C-02 had around 90% natural cover. This suggests that the high percentage of vegetation on the islands in the I-01 could be contributing to the proportion of riparian vegetation utilized by consumers. This potentiality highlights the importance of conserving natural vegetation along the river margins, particularly, in environments that have undergone oligotrophication due to dam construction. The change in the availability of food energy source, particularly from riverine sources, due to the increase in agropastoral activities and absence of natural flood pulse may contribute to decreases in fish biomass in the I-01. Therefore, it is crucial to address land use conservation to mitigate the impact on fish populations in the I-01.
An increase in autochthonous production would be expected with the increase in the order of rivers and consequently an increase in the autochthonous proportion of energy (Carvalho et al., 2023), however our results showed a lower autochthonous proportion of energy in a higher order river. Therefore, it is likely that the process of oligotrophication of the I-01 due to the dam cascade (Agostinho et al., 2002) has altered the energetic dynamics of this environment. Consequently, this environment is unable to support dense fish populations, denoted by a decline in commercial fish catches (Agostinho et al., 2009b), as well as a decrease in fish biomass observed in the years following dam operation (Costa et al., 2012). This process is similar to that observed in the Nile River after the construction of the Aswan Dam in 1965, where fisheries in the river delta collapsed due to decreased availability of nutrients for primary production (Oczkowski et al., 2009). In the I-01, the unnatural trophic state induced by anthropogenic pressures may help explain the reduction in fish biomass in this environment. Allochthonous carbon can support dense fish populations of large, naturally oligotrophic river (Correa, Winemiller, 2018). However, the aquatic community in these environments has a long evolutionary period of adaptation, unlike the I-01 where the oligotrophic state is only a few decades old due to the dam cascade (Agostinho et al., 2007).
The significant autochthonous contribution of the carbon energy source in the I-01 helps explaining why the C-01 and C-02 show greater fish biomass/abundance than the I-01 (Fernandes et al., 2009) because the I-01 is an ultraoligotrophic environment and has a low autochthonous primary production, perhaps not enough to support a greater fish biomass/abundance. The oligotrophication of the I-01 (Agostinho et al., 2002) has occurred due to an increase in nutrient and organic matter retention by the dam, reducing the availability of nutrients downstream (Kobayashi et al., 2008) and, consequently, decreasing autochthonous primary production (Piana et al., 2017). This phenomenon is supported by the lower mean annual concentration of Chla (a proxy for autochthonous primary production; Stadig et al., 2020) in the I-01 compared with the C-01 and C-02, as well as the lower concentration of limiting nutrients for primary production (e.g., phosphorus; Roberto et al., 2009). Therefore, the dam in the I-01 has led to a decrease in the main energy sources of this environment.
Despite a greater total allochthonous contribution for fish in the ultraoligotrophic I-01 compared with the mesotrophic C-01 and the eutrophic C-02, the herbivore species (S. borellii) showed an allochthonous contribution of about 20%. This may reflect the specific feeding habits of this species (Ferretti et al., 1996), as the allochthonous contribution was similar in all rivers, while the autochthonous contribution was around 80%. Therefore, herbivore species may not be the best bioindicators for distinguishing between autochthonous and allochthonous carbon sources, as their feeding habits influence their dietary preferences. Conversely, detritivore, omnivorous, and carnivorous species reflect a broader spectrum of the food web due to their higher trophic position.
Our results are consistent with previous studies that found a significant contribution of riverine energy sources to bottom dwellers fishes (Lopes et al., 2015) and zooplankton (Urbano et al., 2022) in the PIAP, as well as for fish of various guilds in the C-02 (Manetta et al., 2003). The greater riverine contribution raises concerns regarding environmental conservation, particularly for the I-01, which has a higher percentage of anthropic activities along its riverbanks. The presence of agricultural and livestock activities along the river margins reduces the area available for natural primary production, thereby increasing the proportion of energy sources originating from anthropic activities (Carvalho et al., 2015; Effert-Fanta et al., 2023). Increases in anthropic energy sources can destabilize the trophic structure of an aquatic ecosystem, favoring generalist species (Carvalho et al., 2020), altering the availability of natural basal resources, and affecting fish abundance (Effert-Fanta et al., 2023). In contrast, the C-01 and C-02 maintain a high percentage of riparian vegetation conservation, ensuring the stability of the riverine regions and serving as carbon sources for these environments. The importance of riverine energy sources further emphasizes the significance of the environmental protected area of the I-01 islands and floodplain; this designation maintains a high percentage of native vegetation. This vegetation can help support the energy sources for fish even when agriculture is prominent in the riparian areas (Champagne et al., 2022).
Trophic positions of consumers did not show a clear relation with the trophic state of the rivers or the land use in the riparian vegetation area. The C-01 had the highest consumer trophic position, possibly reflecting the high level of environmental conservation in that environment, as has been observed for other sites (Carvalho et al., 2015; Effert-Fanta et al., 2023). This result suggests that high availability of energy sources can sustain food webs with higher trophic levels, similarly to what Wang et al. (2016) observed. Despite the dense occupation of the watershed by agricultural and livestock activities (Urbano et al., unpublished data), the high percentage of native vegetation along the river margins, as well as the presence of PA, may act as buffers against the impacts of land use on aquatic consumers. On the other hand, both the C-02 and I-01 showed a lower trophic position for all analyzed consumers, possibly reflecting the effects of watershed use by agricultural and livestock activities (Pott et al., 2014). Rivers influenced by agricultural and pastural activities tend to exhibit a decrease in the consumer trophic positions (Carvalho et al., 2015) due to the increased input of anthropic energy sources into the food webs (Carvalho et al., 2015), which can alter the calculated trophic position values (Manetta et al., 2003). Furthermore, the high human land cover for the C-02 (Pott et al., 2014) and I-01 (Urbano et al., unpublished data) River basins may cause nitrogen pollution originating from human activities (i.e., fertilizers and sewage), resulting in incorrectly calculated trophic position values due to altered δ15N discrimination (Carvalho et al., 2021). Additionally, we believe that the lower trophic position found in the C-02 may be attributed to its lower order compared with the I-01, making the effects of land use more pronounced.
In addition to our findings, other studies have supported the notion that the riverine region is the main energy source for sustaining the biomass of that environment (Manetta et al., 2003; Lopes et al., 2015; Urbano et al., 2022). Furthermore, suppression of native vegetation by agropastoral activities is positively correlated with a decrease in energy sources for the fish community (Wang et al., 2014). Therefore, the conservation of biodiversity in wetlands relies on the preservation of land–water ecotones to ensure trophic stability. It is crucial to protect riparian vegetation and riverine areas, as they play a vital role in maintaining the health and integrity of aquatic communities that are impacted by anthropogenic factors such as dams and land-use changes.
It is evident that there are irregularities in the protected area, as the riparian area vegetation designation for the I-01 is not fully enforced given the high percentage of human activities in the river margins. Brazilian law neglects the floodplain areas (Brasil, 2012), creating loopholes for deforestation of the riverine vegetation (Grasel et al., 2019) in favor of agribusiness (Metzger et al., 2010). The integrity of the wetland is an essential aspect of protected areas that strive for the sustainable use of resources; however, the protection provided by the law does not uniformly apply to these wetlands. We suggest that local governments and the watershed committee collaborate to propose and implement measures to restore the irregularly protected areas of the I-01 riparian area vegetation in the coming years. It is important to take action to ensure the effective conservation and restoration of these vital ecosystems.
Acknowledgments
This work was supported by the Fundação Araucária de Apoio ao Desenvolvimento Científico e Tecnológico do Estado do Paraná (FA); Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq – Programa de Pesquisas Ecológicas de Longa Duração (PELD) sítio 6; PIAP – Planície de inundação do alto rio Paraná, 141691/2020–4 to MMRS, 308522/2021-4 to EB); Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES – 88881.690087/2022–01 to MMRS, 88882.344471/2019–01 to DDS).
References
Agostinho AA, Bonecker CC, Gomes LC. Effects of water quantity on connectivity: the case of the upper Paraná River floodplain. Ecohydrol Hydrobiol. 2009b; 9(1):99–113. https://doi.org/10.2478/v10104-009-0040-x
Agostinho AA, Gomes LC, Suzuki HI, Júlio Jr. HF. Migratory fishes of the upper Paraná River basin, Brazil. In: Carolsfeld J, Harvey B, Roos C, Baer A (Editors). Migratory fishes of south America: biology, fisheries and conservation status. Ottawa: World Fisheries Trust: The World Bank: International Development Research Centre; 2003. p.19–98.
Agostinho AA, Gomes LC, Veríssimo S, Okada EK. Flood regime, dam regulation and fish in the upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Rev Fish Biol Fisher. 2004; 14:11–19. https://doi.org/10.1007/s11160-004-3551-y
Agostinho A, Pelicice F, Gomes L. Dams and the fish fauna of the neotropical region: impacts and management related to diversity and fisheries. Braz J Biol. 2008; 68:1119–32. https://doi.org/10.1590/s1519-69842008000500019
Agostinho AA, Pelicice FM, Petry AC, Gomes LC, Júlio Jr. HF. Fish diversity in the upper Paraná River basin: habitats, fisheries, management and conservation. Aquat Ecosyst Health Manag. 2007; 10(2):174–86. https://doi.org/10.1080/14634980701341719
Agostinho AA, Thomaz SM, Nakatani K. A planície de inundação do Alto rio Paraná – Site 6. In: Seeliger U, Cordazzo C, Barbosa FA (Editors). Os sites e o programa brasileiro de pesquisas ecológicas de longa duração. Belo Horizonte: UFMG; Programa PELD. 2002. p.101–24.
Agostinho CS, Marques EE, Oliveira RJ, Braz PS. Feeding ecology of Pterodoras granulosus (Siluriformes, Doradidae) in the Lajeado reservoir, Tocantins, Brazil. Iheringia Sér Zool. 2009a; 99(3):301–06. https://doi.org/10.1590/s0073-47212009000300012
Agostinho AA, Bonecker CC, Gomes LC. Effects of water quantity on connectivity: the case of the upper Paraná River floodplain. Ecohydrol Hydrobiol. 2009; 9(1):99–113. https://doi.org/10.2478/v10104-009-0040-x
Alves GHZ, Hoeinghaus DJ, Manetta GI, Benedito E. Dry season limnological conditions and basin geology exhibit complex relationships with δ13C and δ15N of carbon sources in four Neotropical floodplains. PLoS ONE. 2017; 12(3):e0174499. https://doi.org/10.1371/journal.pone.0174499
Anderson MJ. A new method for non‐parametric multivariate analysis of variance. Austral Ecol. 2001; 26(1):32–46. https://doi.org/10.1111/j.1442-9993.2001.01070.pp.x
Arantes CC, Winemiller KO, Asher A, Castello L, Hess LL, Petrere Jr. M et al. Floodplain land cover affects biomass distribution of fish functional diversity in the Amazon River. Sci Rep. 2019; 9:16684. https://doi.org/10.1038/s41598-019-52243-0
Brasil. Lei Nº 12.651, de 25 de maio de 2012. Código Florestal. Brasília; 2012.
Cabana G, Rasmussen JB. Comparison of aquatic food chains using nitrogen isotopes. P Natl A Sci India A. 1996; 93(20):10844–47. https://doi.org/10.1073/pnas.93.20.10844
Canseco JA, Niklitschek EJ, Harrod C. Variabilidade nos fatores de discriminação trófica δ13C e δ15N para peixes teleósteos: uma meta-análise de temperatura e efeitos dietéticos. Rev Fish Biol Fish. 2022; 32(2):313–29. https://doi.org/10.1007/s11160-021-09689-1
Carvalho DR, Castro D, Callisto M, Moreira MZ, Pompeu PS. Isotopic variation in five species of stream fishes under the influence of different land uses. J Fish Biol. 2015; 87(3):559–78. https://doi.org/10.1111/jfb.12734
Carvalho DR, Alves CBM, Flecker AS, Sparks JP, Moreira MZ, Pompeu PS. Using δ15N of periphyton and fish to evaluate spatial and seasonal variation of anthropogenic nitrogen inputs in a polluted Brazilian river basin. Ecol Indic. 2020; 115:106372. https://doi.org/10.1016/j.ecolind.2020.106372
Carvalho DR, Alves CBM, Pompeu PS. Trophic structure of fish assemblages from oligotrophic tropical rivers: evidence of growing assimilation of autochthonous primary producers with the increase in river dimensions. Aquatic Ecology. 2023; 57(2):405–19. https://doi.org/10.1007/s10452-023-10018-w
Carvalho EM, Pereira NS, Ansilago M, Guimarães FJ. Estudo parcial do plano de manejo do parque estadual das Várzeas do Rio Ivinhema como subsídio para ações estratégicas. Braz J Dev. 2019; 5(9):14740–60. https://doi.org/10.34117/bjdv5n9-3151
Caut S, Angulo E, Courchamp F. Variation in discrimination factors (∆15N and ∆13C): the effect of diet isotopic values and applications for diet reconstruction. J Appl Ecol. 2009; 46(2):443–53. https://doi.org/10.1111/j.1365-2664.2009.01620.x
Companhia Energética de São Paulo (CESP). “Porto Primavera”. 2024. Available from: https://www.cesp.com.br/porto-primavera/
Champagne EJ, Guzzo MM, Gutgesell MK, McCann KS. Riparian buffers maintain aquatic trophic structure in agricultural landscapes. Biol Lett. 2022; 18(3):20210598. https://doi.org/10.1098/rsbl.2021.0598
Correa SB, Winemiller K. Terrestrial-aquatic trophic linkages support fish production in a tropical oligotrophic river. Oecologia. 2018; 186(4):1069–78. https://doi.org/10.1007/s00442-018-4093-7
Costa RS, Okada EK, Agostinho AA, Gomes LC. Variação temporal no rendimento e composição específica da pesca artesanal do alto rio Paraná, PR-Brasil: os efeitos crônicos dos barramentos. Bol Inst Pesca. 2012; 38(3):199–213.
Da Cruz DC, Ferreira GC, Ribeiro SS, Schwartz G, Monteiro A. Priority areas for restoration in permanent preservation areas of rural properties in the Brazilian Amazon. Land Use Policy. 2022, 115:106030. https://doi.org/10.1016/j.landusepol.2022.106030
Duda JJ, Coe HJ, Morley SA, Kloehn KK. Establishing spatial trends in water chemistry and stable isotopes (δ15N and δ13C) in the Elwha River prior to dam removal and salmon recolonization. River Res Appl. 2011; 27(10):1169–81. https://doi.org/10.1002/rra.1413
Durán AP, Inger R, Cantú‐Salazar L, Gaston KJ. Species richness representation within protected areas is associated with multiple interacting spatial features. Divers Distrib. 2016; 22(3):300–08. https://doi.org/10.1111/ddi.12404
Effert‐Fanta EL, Chow SL, Wahl DH. Effects of riparian forest and agricultural land use on stream fish diet and trophic position. Ecol Freshw Fish. 2023; 32(2):291–304. https://doi.org/10.1111/eff.12686
Espínola LA, Júlio Junior HF, Benedito E. Invasive non-native species of fish in upper Paraná River Basin, Brazil: variations of caloric content in Cichla kelberi. Neotrop Ichthyol. 2012, 10: 401-408. https://doi.org/10.1590/S1679-62252012005000002
Fernandes R, Agostinho AA, Ferreira EA, Pavanelli CS, Suzuki HI, Lima DP et al. Effects of the hydrological regime on the ichthyofauna of riverine environments of the upper Paraná River floodplain. Braz J Biol. 2009; 69(2):669–80. https://doi.org/10.1590/s1519-69842009000300021
Ferretti CML, Andrian IF, Torrente G. Dieta de duas espécies de Schizodon (Characiformes, Anostomidae) na planície de inundação do alto rio Paraná, e sua relação com aspectos morfológicos. Bol Inst Pesca. 1996; 23:171–86.
Foley JA, DeFries R, Asner GP, Barford C, Bonan G, Carpenter SR et al. Global consequences of land use. Science. 2005; 309(5734):570–74. http://doi.org/10.1126/science.1111772
Freitas CEC, Laurenson L, Yamamoto KC, Forsberg BR, Petrere Jr. M, Arantes C et al. Fish species richness is associated with the availability of landscape components across seasons in the Amazonian floodplain. PeerJ. 2018; 6:e5080. https://doi.org/10.7717/peerj.5080
Fry B. Stable isotope ecology. Springer Science & Business Media; 2006.
Fugi R, Hahn NS, Agostinho AA. Feeding styles of five species of bottom-feeding fishes of the high Paraná River. Environ Biol Fish. 1996; 46:297–307. https://doi.org/10.1007/BF00005006
Giné MF, Zagatto EAG, Reis BF. Simultaneous determination of nitrate and nitrite by flow injection analysis. Anal Chim Acta. 1980; 114:191–97. https://doi.org/10.1016/S0003-2670(01)84290-2
Golterman HL, Clymo RS, Ohmstad MAM. Methods for physical and chemical analysis of freshwater. Blackwell Scientifc. 1978.
Grasel D, Fearnside PM, Vitule JRS, Bozelli RL, Mormul RP, Rodrigues RR et al. Brazilian wetlands on the brink. Biodivers Conserv. 2019; 28(1):255–57. https://doi.org/10.1007/s10531-018-1666-z
Hahn NS, Fugi R, Peretti D, Russo Mr, Loureiro-Crippa VE. Estrutura trófica da ictiofauna da planície de inundação do alto rio Paraná. A planície de inundação do Alto rio Paraná. Maringá: Area de Pesquisas Ecológicas de Longa Duração, Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura-Nupelia, Universidade Estadual de Maringá. 2002; 123–26.
Hoeinghaus DJ, Agostinho AA, Gomes LC, Pelicice FM, Okada EK, Latini JD et al. Effects of river impoundment on ecosystem services of large tropical rivers: embodied energy and market value of artisanal fisheries. Conserv Biol. 2009; 23(5):1222–31. https://doi.org/10.1111/j.1523-1739.2009.01248.x
Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio). Livro vermelho da fauna Brasileira ameaçada de extinção. Vol. 1. Brasília: ICMBio/MMA; 2018. Available from: https://www.icmbio.gov.br/portal/images/stories/comunicacao/publicacoes/publicacoes-diversas/livro_vermelho_2018_vol1.pdf
Isbell F, Balvanera P, Mori AS, He J-S, Bullock JM, Regmi GR et al. Expert perspectives on global biodiversity loss and its drivers and impacts on people. Front Ecol Environ. 2023; 21(2):94–103. https://doi.org/10.1002/fee.2536
Junk WJ, Bayley PB, Sparks RE. The flood pulse concept in river-floodplain systems. Canadian special publication of fisherries and aquatic sciences. 1989; 106:110–27.
Kobayashi JT, Thomaz SM, Pelicice FM. Phosphorus as a limiting factor for Eichhornia crassipes growth in the upper Paraná River floodplain. Wetlands. 2008; 28:905–13. https://doi.org/10.1672/07-89.1
Lajtha K, Michener RH. Stable isotopes in ecology and environmental science. Malden: Blackwell Scientific Publications; 1994.
Lamparelli MC. Graus de trofia em corpos d’água do estado de São Paulo: avaliação dos métodos de monitoramento [PhD Thesis]. São Paulo: Universidade de São Paulo; 2004.
Legendre P, Legendre L. Numerical ecology. Elsevier, Amsterdam; 1998.
Lobón-Cerviá J, Hess LL, Melack JM, Araujo-Lima CARM. The importance of forest cover for fish richness and abundance on the Amazon floodplain. Hydrobiologia. 2015; 750(1):245–55. https://doi.org/10.1007/s10750-014-2040-0
Lopes CA, Manetta GI, Figueiredo BR, Martinelli LA, Benedito E. Carbon from littoral producers is the major source of energy for bottom-feeding fish in a tropical floodplain. Environ Biol Fish. 2015; 98(4):1081–88. https://doi.org/10.1007/s10641-014-0343-7
Mackereth F. Water analysis: some revised methods for limnologists. Freshw Biol Assoc Sci Pub. 1978; 36:117.
Manetta GI, Benedito-Cecilio E, Martinelli M. Carbon sources and trophic position of the main species of fishes of Baía River, Paraná River floodplain, Brazil. Braz J Biol. 2003; 63(2):283–90. https://doi.org/10.1590/s1519-69842003000200013
Manetta GI, Benedito-Cecilio E. Aplicação da técnica de isótopos estáveis na estimativa da taxa de turnover em estudos ecológicos: uma síntese. Acta Sci Biol Sci. 2003; 25(1):121–29. https://doi.org/10.4025/actascibiolsci.v25i1.2090
Manetta GI, Scoarize MMR, Delanira-Santos D, Sacramento PA, Urbano VA, Benedito E. Diet-tissue discrimination factors of three neotropical freshwater fishes and a comparison of the trophic position. J Liminol. 2023; 82. https://doi.org/10.4081/jlimnol.2023.2159
McCutchan JH, Lewis Jr. WM, Kendall C, McGrath CC. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos. 2003; 102(2):378–90. https://doi.org/10.1034/j.1600-0706.2003.12098.x
McWilliam-Hughes SM, Jardine TD, Cunjak RA. Instream C sources for primary consumers in two temperate, oligotrophic rivers: possible evidence of bryophytes as a food source. J N Am Benthol Soc. 2009; 28(3):733–43. https://doi.org/10.1899/08-103.1
Metzger JP, Lewinsohn TM, Joly CA, Verdade LM, Martinelli LA, Rodrigues RR. Brazilian law: full speed in reverse? Science. 2010; 329(5989):276–77. https://doi.org/10.1126/science.329.5989.276-b
Michener RH. Stable isotope ratios and tracers in marine aquatic food web. Stable isotopes in ecology and environmental science. 1994.
Murphy CA, Arismendi I, Taylor GA, Johnson SL. Evidence for lasting alterations to aquatic food webs with short-duration reservoir draining. PLoS ONE. 2019; 14(2):e0211870. https://doi.org/10.1371/journal.pone.0211870
Oczkowski AJ, Nixon SW, Granger SL, El-Sayed AM, McKinney RA. Anthropogenic enhancement of Egypt’s Mediterranean fishery. P Natl A Sci India A. 2009; 106(5):1364–67. https://doi.org/10.1073/pnas.0812568106
Okawa C. Caracterização do regime de fluxos a jusante da usina hidrelétrica Porto Primavera no rio Paraná: estação Porto São José. Rev Tecnol. 2010; 19(1):67–74. https://doi.org/10.4025/revtecnol.v19i1.9059
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D et al. Imports MASS. Package ‘vegan’. Community ecology package. 2019.
Parnell AC, Phillips DL, Bearhop S, Semmens BX, Ward EJ, Moore JW et al. Bayesian stable isotope mixing models. Environmetrics. 2013; 24(6):387–99. https://doi.org/10.1002/env.2221
Petsch DK, Cionek VDM, Thomaz SM, Santos NCL. Ecosystem services provided by river-floodplain ecosystems. Hydrobiologia. 2023; 850:2563–84. https://doi.org/10.1007/s10750-022-04916-7
Piana PA, Cardoso BF, Dias J, Gomes LC, Agostinho AA, Miranda LE. Using long-term data to predict fish abundance: the case of Prochilodus lineatus (Characiformes, Prochilodontidae) in the intensely regulated upper Paraná River. Neotrop Ichthyol. 2017; 15(3):e160029. https://doi.org/10.1590/1982-0224-20160029
Pochat V. International agreements, institutions and projects in La Plata River basin. Int J Water Resour Dev. 2011; 27(3):497–510. https://doi.org/10.1080/07900627.2011.597830
Post DM. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology. 2002; 83(3):703–18. https://doi.org/10.1890/0012-9658(2002)083[0703:usitet]2.0.co;2
Pott A, Silva JSV, Gomes EL. Características da bacia hidrográfica do rio Ivinhema. Rev GeoPantanal. 2014; 9(16):109–24.
R Development Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2023. Available from: https://www.r-project.org/
Ragonha FH, Tramonte RP, Takeda AM. Fauna de macroinvertebrados no Parque Estadual das Várzeas do rio Ivinhema-MS. Arq MUDI. 2014; 18(1):28–42.
Reis BF, Zagatto EAG. A new device for improving sensitivity and stabilization in flow-injection analysis. Anal Chim Acta. 1978; 97(2):427–31.
Ribeiro MC, Metzger JP, Martensen AC, Ponzoni FJ, Hirota MM. The Brazilian Atlantic Forest: how much is left, and how is the remaining forest distributed? Implications for conservation. Biol Conserv. 2009; 142(6):1141–53. https://doi.org/10.1016/j.biocon.2009.02.021
Roberto MC, Santana NF, Thomaz SM. Limnology in the upper Paraná River floodplain: large-scale spatial and temporal patterns, and the influence of reservoirs. Braz J Biol. 2009; 69:717–25. https://doi.org/10.1590/s1519-69842009000300025
Rocha PC. Sistemas rio-planície de inundação: geomorfologia e conectividade hidrodinâmica. Cad Prudentino Geogr. 2011; 1(33):50–67.
Ruaro R, Gubiani EA, Hughes RM, Mormul RP. Global trends and challenges in multimetric indices of biological condition. Ecol Indic. 2020; 110:105862. https://doi.org/10.1016/j.ecolind.2019.105862
Sacramento PA, Manetta GI, Benedito E. Diet‐tissue discrimination factors (∆13C and ∆15N) and turnover rate in somatic tissues of a neotropical detritivorous fish on C3 and C4 diets. J Fish Biol. 2016; 89(1):213–19. https://doi.org/10.1111/jfb.12859
Santana ARA, Lansac-Tôha FA, Benedito E. Variability of δ13C and δ15N in three zooplankton species from the Upper Paraná River floodplain. Zoologia. 2009; 26(4):725–32. https://doi.org/10.1590/S1984-46702009000400017
Souza Jr. CM, Shimbo JZ, Rosa MR, Parente LL, Alencar AA, Rudorff BFT et al. Reconstructing three decades of land use and land cover changes in Brazilian biomes with Landsat archive and earth engine. Remote Sens. 2020; 12(17):2735. https://doi.org/10.3390/rs12172735
Smith JA, Mazumder D, Suthers IM, Taylor MD. Para ajustar ou não ajustar: avaliando modelos de mistura de isótopos estáveis usando polígonos de mistura simulados. Methods Ecol Evol. 2013; 4(7):612–18. https://doi.org/10.1111/2041-210X.12048
Stadig MH, Collingsworth PD, Lesht BM, Höök TO. Spatially heterogeneous trends in nearshore and offshore chlorophyll a concentration in lakes Michigan and Huron (1998–2013). Freshw Biol. 2020; 65(3):366–78. https://doi.org/10.1111/fwb.13430
Tessler M, Brugler MR, DeSalle R, Hersch R, Velho LFM, Segovia BT et al. A global eDNA comparison of freshwater bacterioplankton assemblages focusing on large-river floodplain lakes of Brazil. Microb Ecol. 2017; 73:61–74. https://doi.org/10.1007/s00248-016-0834-5
Torrecilha S. Plano de manejo Parque Estadual das Várzeas do Rio Ivinhema, Sylvia Torrecilha. 2008. Available from: https://www.imasul.ms.gov.br/wp-content/uploads/2015/06/Parque-Estadual-das-V%C3%A1rzeas-do-Rio-Ivinhema-PEVRI.pdf
Urbano VA, Delanira-Santos D, Benedito E. The role of cladocerans in green and brown food web coupling. Iheringia Sér Zool. 2022; 112:e2022022. https://doi.org/10.1590/1678-4766e2022022
Vanderklift MA, Ponsard S. Sources of variation in consumer-diet?15N enrichment: a meta-analysis. Oecologia, 2003; 136(2):169–82. https://doi.org/10.1007/s00442-003-1270-z
Wang J, Gu B, Huang J, Han X, Lin G, Zheng F, Li Y. Terrestrial contributions to the aquatic food web in the middle Yangtze River. PLoS ONE. 2014; 9(7):e102473. https://doi.org/10.1371/journal.pone.0102473
Wang Y, Xiao X, Yu X, Xu J, Cai Y, Lei G. Resource availability determines food chain length in Chinese subtropical rivers. Aquat Ecol. 2016; 50(2):187–95. https://doi.org/10.1007/s10452-016-9567-2
Xiang T, Dong X, Ju T, Shi L, GrFenouillet G. Anthropogenic activities and environmental filtering have reshaped freshwater fish biodiversity patterns in China over the past 120 years. J Environ Manage. 2023; 344:118374. https://doi.org/10.1016/j.jenvman.2023.118374
Authors

![]() Vinícius de Andrade Urbano1
Vinícius de Andrade Urbano1 ![]() ,
, ![]() Driele Delanira-Santos1,2,
Driele Delanira-Santos1,2, ![]() Matheus Maximilian Ratz Scoarize1,2,3 and
Matheus Maximilian Ratz Scoarize1,2,3 and ![]() Evanilde Benedito1,2
Evanilde Benedito1,2
[1] Departamento de Biologia, Universidade Estadual de Maringá, Av. Colombo, 5790, 87020-900 Maringá, PR, Brazil. (VAU) viniciusaurbano@gmail.com (corresponding author).
[2] Programa de Pós-Graduação em Ecologia de Ambientes Aquáticos, Universidade Estadual de Maringá, Av. Colombo, 5790, 87020-900 Maringá, PR, Brazil. (MMRS) maxscoarize@gmail.com, (DDS) drieledelanira@gmail.com, (EB) eva@nupelia.uem.br.
[3] Australian Rivers Institute (ARI), Griffith University, Australia.
Authors’ Contribution 

Vinícius de Andrade Urbano: Conceptualization, Formal analysis, Investigation, Methodology, Writing-original draft, Writing-review and editing.
Driele Delanira-Santos: Conceptualization, Formal analysis, Investigation, Methodology, Supervision.
Matheus Maximilian Ratz Scoarize: Methodology, Writing-original draft, Writing-review and editing.
Evanilde Benedito: Conceptualization, Methodology, Resources, Supervision, Writing-original draft, Writing-review and editing.
Ethical Statement
This project has received research permits from the federal (ICMBio nº 52596–5) and state (IMASUL nº 71/000666/2021) levels, as well as ethical approval from the Universidade Estadual de Maringá (UEM) ethics committee (CEUA nº 1420221018).
Competing Interests
The author declares no competing interests.
How to cite this article
Urbano VA, Delanira-Santos D, Scoarize MMR, Benedito E. Dams and agricultural lands affect energy sources and the trophic position of fish in a floodplain. Neotrop Ichthyol. 2024; 22(3):e230084. https://doi.org/10.1590/1982-0224-2023-0084
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2024 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted August 27, 2024 by Caroline Arantes
Accepted August 27, 2024 by Caroline Arantes
![]() Submitted July 20, 2023
Submitted July 20, 2023
![]() Epub October 18, 2024
Epub October 18, 2024