![]() Thiago Henrique Pedroso1
Thiago Henrique Pedroso1 ![]() ,
, ![]() Gabriel de Carvalho Deprá1 and
Gabriel de Carvalho Deprá1 and ![]() Carla Simone Pavanelli1,2
Carla Simone Pavanelli1,2
PDF: EN XML: EN | Cite this article
Abstract
Five species of Bryconamericus are known from the upper rio Paraná basin: ‘B.’ aff. iheringii, ‘B.’ coeruleus, B. stramineus, ‘B.’ turiuba and the non-native B. exodon. The new species can be easily distinguished from them by its body depth (27.8–31.8% SL) and the teeth aligned in the outer row of the premaxilla (except ‘B.’ aff. iheringii). The new species is distributed in tributaries to the Piquiri, Ivaí and Tibagi rivers, at altitudes between 498 and 900 m a.s.l. The geographic distribution of the new species is similar to the congener ‘B.’ coeruleus. Other species with similar distribution are Apareiodon vladii and Planaltina kaingang, although they only occur in the Piquiri and Ivaí river basins. Furthermore, this work records the new species with the presence of breeding tubercles. These tubercles are located on the dorsal and lateral portions of the head and on the posterior margin of the scales of mature males; and in females, when present, they are smaller and restricted to the posterior margin of the scales. Comments are made on the presence of breeding tubercles in other groups of fishes, and also on some dimorphic characters present in the new species.
Keywords: Drainage rearrangement, Gonadal stage, Sexual dimorphism, Taxonomy.
Cinco espécies de Bryconamericus são conhecidas da bacia do alto rio Paraná: ‘B.’aff. iheringii, ‘B.’ coeruleus, B. stramineus, ‘B.’ turiuba e a não nativa B. exodon. A nova espécie pode ser facilmente diferenciada delas pela altura do corpo (27,8–31,8% CP) e pelos dentes alinhados na série externa do pré-maxilar. A nova espécie está distribuída em afluentes dos rios Piquiri, Ivaí e Tibagi, em altitudes entre 498 e 900 m acima do nível do mar. A distribuição geográfica da nova espécie é semelhante à da congênere ‘B.’ coeruleus. Outras espécies com distribuição semelhante são Apareiodon vladii e Planaltina kaingang, embora ocorram apenas nas bacias dos rios Piquiri e Ivaí. Além disso, este trabalho registra uma nova espécie com a presença de tubérculos nupciais. Estes tubérculos estão localizados nas porções dorsal e lateral da cabeça e na margem posterior das escamas de machos sexualmente maduros; e nas fêmeas, quando presentes, são menores e restritos à margem posterior das escamas. Comentários são apresentados sobre a presença de tubérculos nupciais em outros grupos de peixes, e também sobre alguns caracteres dimórficos presentes na nova espécie.
Palavras-chave: Dimorfismo sexual, Estágio gonadal, Rearranjo de drenagem, Taxonomia.
Introduction
Bryconamericus Eigenmann, 1907 is one of the most species-rich genera in Stevardiinae, containing 54 valid species (Fricke et al.,2023), distributed in a variety of freshwater ecosystems from Central America to northern Argentina, on both sides of the Andean cordilleras (Vari, Siebert, 1990; Jerep, Shibatta, 2017). The polyphyletic nature of Bryconamericus has been hypothesized by Eigenmann (1927), Fink (1976), Vari, Siebert (1990), and Malabarba, Kindel (1995), and corroborated by several phylogenetic studies (Thomaz et al.,2015; García-Melo et al.,2019; Mirande, 2019). However, the phylogenetic position of many species currently included in the genus is still poorly understood.
Mirande(2019) recovered a clade called “Nantis clade” that includes several species from Southern South America: ‘B.’ iheringii (Boulenger, 1887), ‘B.’ rubropictus (Berg, 1901), ‘B.’ uporas Casciotta, Almirón & Azpelicueta, 2004 and ‘B.’ ikaa Casciotta, Almirón & Azpelicueta, 2004, although the clade probably includes other species from the Southern Neotropics, which are morphologically similar to ‘B.’ iheringii. This clade appears as sister-group to “Hypobrycon clade” and Odontostoechus lethostigmus Gomes, 1947. In turn, the clade that includes the type-species Bryconamericus exodon Eigenmann, 1907 appears relatively distant from the “Nantis clade”, confirming the polyphyly of the genus and showing the uncertain phylogenetic position of most species currently assigned to Bryconamericus.Species from Northern South America, on the other hand, are more likely to be related to other genera, such as Attonitus Vari & Ortega, 2000, Ceratobranchia Eigenmann, 1914, Eretmobrycon Fink, 1976, Hemibrycon Günther, 1864, and Knodus Eigenmann, 1911(THP, pers. obs.).
As presently understood, Bryconamericus does not have a morphological diagnosis. Thus, in the absence of phylogenetic evidence supporting the inclusion of a new species in the genus, that inclusion must be based on the possession of the combination of characters proposed by Eigenmann (1927), and modified by Vari, Siebert (1990): premaxilla with two rows of teeth, the inner row with four teeth, which are larger than those in the external row, zero to six teeth distributed along the anterior margin of the maxilla, complete lateral line, caudal fin naked, single tooth row on the dentary, setiform gill rakers, absence of a glandular pouch on the caudal fin of males, and third infraorbital expanded and in contact with the preopercle. Recent sampling efforts in the upper rio Paraná basin have yielded specimens fitting the aforementioned definition, which do not correspond to any other described species and are herein formally described as a new species of Bryconamericus.
Material and methods
Morphometric and meristic data were taken on the left side of the specimens, following Fink, Weitzman (1974), except for the gill rakers count, which was made on the right side of the fish. Osteological data were obtained as in Deprá et al. (2021:4, tab. 2) from specimens cleared and stained (c&s) as in Taylor, Van Dyke (1985). The nomenclature of types of vertebrae were also obtained as in Deprá et al. (2021:4, fig. 1). Ribs were counted starting from the fifth vertebra, i.e., ribs modified into Weberian apparatus ossicles were not counted (e.g., the rib of vertebra 4 contributes to the formation of the os suspensorium; see Grande, Young, 2004). Sexual dimorphism traits were confirmed by the analysis of gonadal development state performed in 16 specimens through a small cut on the ventral region of the body. In the description, the frequency of each meristic count is given in parentheses, and the counts of the holotype are marked by an asterisk. Morphometric data are presented as percentages of standard length (SL), except subunits of the head as percentages of the head length (HL). Institutional abbreviations followed Sabaj (2022). For the species of which specimens were not available for analysis, the respective original description or a redescription were used as a mean of comparison: Berg (1901) and Braga (2000), for ‘B.’ rubropictus; Bizerril, Peres-Neto (1995), for ‘B.’ ornaticeps; Miquelarena, Aquino (1999), for ‘B.’ eigenmanni; Miquelarena et al. (2002), for‘B.’ mennii, and Mirande et al. (2004), for N. indefessus (= ‘B.’ indefessus). For the other species, of which specimens were available for comparison, the material is listed under “Comparative material examined”.

FIGURE 1| Bryconamericus misei. A. NUP 24255, holotype, 54.8 mm SL, male, unnamed stream, tributary to the rio Capivara, rio Piquiri basin, Campina do Simão, Paraná State, Brazil; B. NUP 24149, paratype, 54.9 mm SL, female. C. NUP 24155, paratype, 26.4 mm SL.
Results
Bryconamericus misei, new species
urn:lsid:zoobank.org:act:2CAF55C7-5B39-4C05-9318-8C69B783565D
(Figs. 1–4; Tab. 1)
Bryconamericus aff. iheringii (non Boulenger, 1887). —Cavalli et al., 2018:5 (checklist of species of Piquiri River; citation).
Holotype. NUP 24255, 54.8 mm SL, male, Brazil, Paraná State, Campina do Simão, unnamed stream, tributary to the rio Capivara, rio Piquiri basin, 25°05’43.02”S 51°49’29.00” W, 14 Jun 2019, F. T. Mise.
Paratypes. All from the Paraná State, upper rio Paraná basin, Brazil.MZUEL 18401, 5, 42.3–46.0 mm SL, Ortigueira, rio Apucarana, tributary to the rio Tibagi, 24°05’26.82”S 51°00’54.66”W, 21 Sep 2017, I. Vitture, A. Souza, R. Ono & N. Narezzi. MZUEL 18438, 3, 26.4–42.4 mm SL, córrego Água Boa, tributary to the rio Apucarana, tributary to the rio Tibagi, 23°57’40.43”S 51°01’17.46”W, 14 Nov 2017, I. Vitture, E. Santana, A. Souza & R. Ono. NUP 15777, 5, 41.9–53.7 mm SL, Goioxim, rio Bonito, tributary to the rio Piquiri, 25°04’25.80”S 52°04’05.79”W, 26 Jan 2014, W. J. Graça, W. M. Domingues, F. A. Teixeira & R. J. Graça. NUP 16042, 3, 36.3–45.0 mm SL, Luiziana, rio Laranjeiras, tributary to the rio Ivaí, 24°20’26.85”S 52°11’42.50”W, 28 Jan 2014, W. J. Graça, W. M. Domingues, F. A. Teixeira & R. J. Graça. NUP 18274, 11, 35.9–47.8 mm SL, same locality as the holotype, 3 Oct 2015, W. J. Graça, F. A. Teixeira, W. M. Domingues & A. Frota. NUP 18277, 9, 27.0–38.2 mm SL, Campina do Simão, unnamed stream, tributary to the rio Capivara, rio Piquiri basin, 25°08’38.49”S 51°49’29.01”W, 3 Oct 2015, W. J. Graça, F. A. Teixeira, W. M. Domingues & A. Frota. NUP 24149, 3, 18.4–54.9 mm SL, same locality as the holotype, 7 Dec 2018, F. T. Mise. NUP 24150, 8, 25.8–49.8 mm SL, same locality as the holotype, 14 Jun 2019, F. T. Mise. NUP 24151, 3, 44.0–56.9 mm SL, same locality as the holotype, 16 Oct 2018, F. T. Mise. NUP 24152, 1, 39.1 mm SL, Campina do Simão, unnamed stream, tributary to the rio Capivara, rio Piquiri basin, 25°08’38.49”S 51°49’29.01”W, 22 Mar 2019, F. T. Mise. NUP 24153, 2, 37.8–50.2 mm SL, same locality as NUP 24152, 30 Nov 2019, F. T. Mise. NUP 24154, 7, 36.5–45.4 mm SL, same locality as the holotype, 30 Nov 2019, F. T. Mise. NUP 24155, 18, 20.6–44.7 mm SL, same locality as NUP 24152, 14 Jun 2019, F. T. Mise.
TABLE 1 | Morphometric data of Bryconamericus misei. N = number of specimens; SD = Standard deviation.
Characters | Holotype | Males | Females | ||||||
N | Range | Mean | SD | N | Range | Mean | SD | ||
Standard length (mm) | 54.8 | 16 | 39.1–50.2 | 43.4 |
| 13 | 40.7–56.9 | 46.5 |
|
Percents of standard length | |||||||||
Depth at dorsal fin origin | 29.4 | 16 | 28.9–31.8 | 30.3 | 0.7 | 13 | 27.8–31.7 | 30.0 | 1.0 |
Snout to dorsal fin origin | 53.1 | 16 | 52.7–56.0 | 54.0 | 1.0 | 13 | 53.0–56.5 | 54.7 | 1.0 |
Snout to pelvic fin origin | 46.9 | 16 | 45.0–48.6 | 47.0 | 1.1 | 13 | 46.5–49.0 | 47.7 | 0.8 |
Snout to anal fin origin | 63.5 | 16 | 61.9–66.1 | 63.8 | 1.3 | 13 | 63.8–67.3 | 65.4 | 1.1 |
Caudal peduncle depth | 12.4 | 16 | 11.1–13.3 | 12.2 | 0.5 | 13 | 11.1–12.2 | 11.7 | 0.3 |
Caudal peduncle length | 15.0 | 16 | 14.3–17.3 | 15.8 | 0.9 | 13 | 14.3–17.1 | 15.5 | 0.7 |
Pectoral fin length | 21.7 | 16 | 20.0–22.9 | 21.5 | 0.7 | 13 | 18.8–21.3 | 20.2 | 0.9 |
Pelvic fin length | 15.5 | 16 | 14.4–17.7 | 16.5 | 0.8 | 13 | 13.9–15.7 | 14.8 | 0.6 |
Dorsal fin length | 21.7 | 16 | 20.9–25.9 | 23.5 | 1.2 | 13 | 20.2–23.0 | 22.1 | 0.9 |
Dorsal fin base length | 13.0 | 16 | 11.5–14.5 | 13.1 | 0.7 | 13 | 12.3–13.9 | 12.8 | 0.4 |
Anal fin length | 17.3 | 16 | 16.9–20.5 | 18.6 | 1.0 | 13 | 15.8–18.8 | 17.2 | 0.7 |
Anal fin base length | 25.4 | 16 | 21.2–26.6 | 24.6 | 1.3 | 13 | 20.8–23.9 | 22.6 | 0.9 |
Dorsal fin origin to caudal fin base | 55.3 | 16 | 49.1–53.2 | 51.1 | 1.3 | 13 | 48.5–54.3 | 50.7 | 2.0 |
Orbit to dorsal fin origin | 40.5 | 16 | 39.1–42.6 | 40.7 | 0.9 | 13 | 40.1–43.3 | 41.9 | 0.8 |
Head length | 26.6 | 16 | 25.6–28.0 | 26.7 | 0.6 | 13 | 25.2–28.3 | 26.3 | 1.0 |
Percents of head length | |||||||||
Orbit diameter | 29.5 | 16 | 29.7–33.3 | 31.5 | 1.0 | 13 | 28.1–33.9 | 30.7 | 1.6 |
Snout length | 24.7 | 16 | 22.9–25.7 | 24.2 | 0.8 | 13 | 22.0–27.8 | 25.4 | 1.5 |
Interorbital width | 31.5 | 16 | 30.4–34.3 | 31.8 | 1.1 | 13 | 30.7–33.6 | 31.7 | 0.9 |
Upper jaw length | 38.4 | 16 | 35.4–41.6 | 38.3 | 1.4 | 13 | 36.5–40.2 | 38.6 | 1.3 |
Diagnosis. Among the Stevardiinae genera endemic to the Southern Neotropics, Bryconamericus misei can be distinguished from species of Glandulocaudini, Bryconamericus sensu stricto (i.e., B. exodon and B. stramineus Eigenmann, 1908; Mirande, 2019), some species of Diapoma Cope, 1894, Hysteronotus Eigenmann, 1911, Lepidocharax Ferreira, Menezes & Quagio-Grassiotto, 2011, Piabarchus Myers, 1928, Piabina Reinhardt, 1867, Planaltina Böhlke, 1954,and Pseudocorynopoma Perugia, 1891 by having only a few, unmodified scales on the very base of the caudal fin (vs. presence of modified scales associated with glandular tissue on the ventral caudal-fin lobe of males and females of Planaltina and Diapoma pyrrhopteryx Menezes & Weitzman, 2011, D. speculiferum Cope, 1894, D. terofali (Géry, 1964), and D. thauma Menezes & Weitzman, 2011 and of males of Hysteronotus, and Pseudocorynopoma; presence of modified scales associated with glandular tissue on the dorsal caudal-fin lobe of males of Glandulocaudini; and non-modified scales covering more than one third of each caudal-fin lobe in Lepidocharax [also Knodus]); anal-fin origin approximately at a vertical through the dorsal-fin terminus (vs. anterior to a vertical through the middle of the dorsal-fin base in Glandulocaudini [except Lophiobrycon Castro, Ribeiro, Benine & Melo, 2003], Hysteronotus, Lepidocharax, Piabarchus, Planaltina, and Pseudocorynopoma); 8 total pelvic-fin rays (a single specimen with 7; vs. 7 in Diapoma, Lepidocharax, and Planaltina); retrognathous mouth (vs. isognathous or prognathous in Glandulocaudini, Diapoma, Hysteronotus, Lepidocharax, Planaltina, and Pseudocorynopoma; and teeth in the outer premaxillary series arranged in a regular line (vs. not aligned in Bryconamericus sensu stricto and Piabina [also in Creagrutus Günther, 1864 and ‘B.’ coeruleus Jerep & Shibatta, 2017, ‘B.’ mennii Miquelarena, Protogino, Filiberto & López, 2002, ‘B.’ turiuba Langeani, Lucena, Pedrini & Tarelho-Pereira, 2005, and Knodus moenkhausii (Eigenmann & Kennedy, 1903)]).
Another lineage of Stevardiinae endemic to Southern Neotropics includes the nominal genera Hypobrycon Malabarba & Malabarba, 1994, Nantis Mirande, Aguilera & Azpelicueta, 2006 and Odontostoechus Gomes, 1947, in addition to ‘B.’ iheringii and several similar species currently assigned to ‘Bryconamericus’ (Mirande, 2019; B. misei appears to belong in that lineage).Bryconamericus misei is distinguished from all members of that lineage, except for ‘B.’ ecai da Silva, 2004, ‘B.’ eigenmanni (Evermann & Kendall, 1906), ‘B.’ ikaa Casciotta, Almirón & Azpelicueta, 2004, ‘B.’ sylvicola Braga, 1998 and ‘B.’ ytu Almirón, Azpelicueta & Casciotta, 2004 by having a vertically elongated black humeral spot, followed by a second inconspicuous spot vs. a single vertically extended spot in Nantis indefessus Mirande, Aguilera & Azpelicueta, 2004, Hypobrycon, ‘B.’ agna Azpelicueta & Almirón, 2001, ‘B.’ microcephalus (Miranda Ribeiro, 1908), ‘B.’ ornaticeps Bizerril & Perez-Neto, 1995, ‘B.’ patriciae da Silva, 2004, ‘B.’ rubropictus (Berg, 1901), and ‘B.’ tenuis Bizerril & Auraujo, 1992; a single, rounded humeral spot in Odontostoechus lethostigmus, ‘B.’ lambari Malabarba & Kindel, 1995, ‘B.’ pyahu Azpelicueta, Casciotta & Almirón, 2003 and ‘B.’ uporas Casciotta, Azpelicueta & Almirón, 2002; Bryconamericus misei differs from ‘B.’ ecai, ‘B.’ eigenmanni, ‘B.’ iheringii, ‘B.’aff. iheringii from the upper rio Paraná (Frota et al., 2016; Reis et al., 2020),‘B.’ ikaa, ‘B.’ sylvicola and ‘B.’ ytu, as well from ‘B.’ microcephalus by having a body depth of 27.8–31.8% SL vs. 33.1–36.9% in ‘B.’ ecai, 33.7–42.3% in ‘B.’ iheringii, 34.2–39.3% in ‘B.’aff. iheringii (Tab. 2), 33.7–36.4% in ‘B.’ ikaa, 23.5–27.7% in ‘B.’ microcephalus, 36.1–40.7% in ‘B.’ sylvicola and 34.6–37.9% in ‘B.’ ytu.
TABLE 2 | Morphometric data of ‘Bryconamericus’ iheringii from the Laguna dos Patos in comparison with ‘B.’aff. iheringii from the upper rio Paraná basin. The diagnostic characters between these two species and B. misei are highlighted in bold. N = number of specimens.
Characters | ‘B.’ iheringii Laguna dos Patos | ‘B.’ iheringii upper rio Paraná | ||||
Range | N | Mean | Range | N | Mean | |
Standard length (mm) | 43.3–61.2 | 30 | 50.6 | 44.2–58.2 | 20 | 51.0 |
Percents of standard length | ||||||
Depth at dorsal fin origin | 33.7–42.3 | 30 | 36.2 | 34.2–39.3 | 20 | 36.9 |
Snout to dorsal fin origin | 52.3–58.1 | 30 | 55.7 | 53.0–57.0 | 20 | 55.4 |
Snout to pelvic fin origin | 47.6–52.2 | 30 | 49.5 | 46.9–51.5 | 20 | 49.5 |
Snout to anal fin origin | 65.2–70.4 | 30 | 67.5 | 64.8–70.4 | 20 | 67.6 |
Dorsal fin length | 23.5–27.6 | 30 | 25.0 | 20.9–26.6 | 20 | 24.5 |
Anal fin base length | 21.6–27.8 | 30 | 24.9 | 22.1–26.6 | 20 | 24.4 |
Orbit to dorsal fin origin | 39.5–45.3 | 30 | 42.4 | 40.0–44.7 | 20 | 42.7 |
Head length | 24.3–26.9 | 30 | 25.3 | 23.6–26.5 | 20 | 25.0 |
Percents of head length | ||||||
Orbit diameter | 33.6–39.7 | 30 | 37.0 | 34.8–40.9 | 20 | 37.1 |
Interorbital width | 29.3–33.7 | 30 | 31.5 | 29.9–36.6 | 20 | 32.4 |
In addition, Bryconamericus misei differs from ‘B.’ eigenmanni, and‘B.’ iheringii by having 13–16 total external gill rakers on first branchial arch vs. 17–21; from ‘B.’ pyahu by having 6–7 gill rakers on epibranchial vs. 4–5 in ‘B.’ pyahu; from adults of Odontostoechus lethostigmus by having two rows of teeth on the premaxilla vs. a single row; from Nantis indefessus by having four teeth in the inner row of premaxilla vs. five; from ‘B.’ ornaticeps and ‘B.’ sylvicola by having 16–19 branched anal-fin rays vs. 14–15 in ‘B.’ ornaticeps and22–25 in ‘B.’ sylvicola; from ‘B.’ agna and ‘B.’ uporas by having 3–5 cusps on the inner-series premaxillary teeth vs. 7; from ‘B.’ tenuis, by having distally compressed teeth vs. massive teeth; from Hypobrycon, by having dentary teeth positioned anterodorsally vs. teeth positioned along the anterior margin of dentary (compare Fig. 2D with fig. 1 in Malabarba, Malabarba, 1994); from ‘B.’ agna and H. poi Almirón, Casciotta, Azpelicueta & Cione, 2001 by having 8–11 teeth on dentary vs. 6–7; from ‘B.’ microcephalus, ‘B.’ iheringii and from ‘B.’aff. iheringii from the upper rio Paraná by having a lower orbital diameter (28.1–33.9% HL vs. 38.4–41.6% in ‘B.’ microcephalus, 33.6–39.7% in ‘B.’ iheringii and 34.8–40.9% in ‘B.’aff. iheringii; Tab. 2).
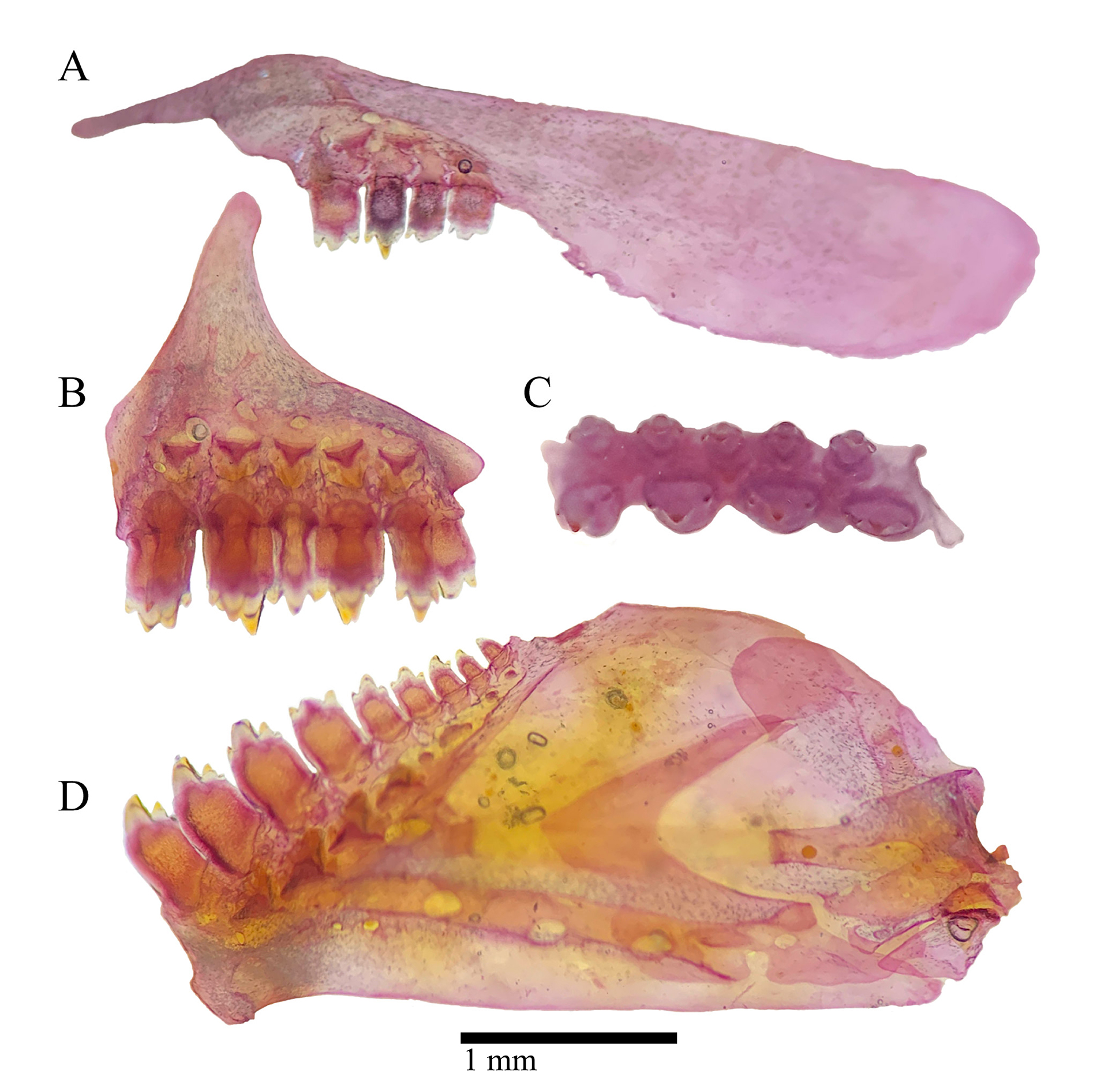
FIGURE 2| Left jaws of Bryconamericus misei, NUP 24155, paratype, 44.7 mm SL. A. Maxilla. B. Premaxilla in anterior view. C. Ventral view of premaxilla. D. Dentary in lateral view.
Description. Morphometric data presented in Tab. 1. Dorsal profile of head convex from tip of snout to vertical through anterior border of nostrils, slightly convex from nostrils to dorsal-fin origin; straight from that point to adipose-fin origin, slightly concave along caudal peduncle. Ventral profile convex from tip of dentary to anal-fin origin; straight from that point to end of anal-fin, slightly concave along caudal peduncle.
Mouth slightly retrognathous, always positioned at level of ventral border of orbit or below (Fig. 3). Posterior tip of maxilla exceeding vertical through anterior limit of orbit. Outer premaxillary tooth row with three(3), four*(17) or five(10) tricuspid teeth; inner row with four*(30) tri to pentacuspid teeth; maxilla with two(2), three(12), four(14), five*(1) or six(1) tricuspid teeth; dentary with four large tri to tetracuspid teeth anteriorly, and four(7), five(17), six(2) or seven*(2) smaller teeth gradually decreasing in size posteriorly. External gill rakers on first arch 6*(13) or 7(17) on upper limb and 7(3), 8(17) or 9*(10) on lower limb, counted in entire specimens.

FIGURE 3| Head shape and sexual dimorphism in Bryconamericus misei. A. NUP 24150, holotype, 54.8 mm SL. Male with breeding tubercles on dorsal and lateral portions of the head and posterior margin of the scales. B. NUP 24149, paratype, 54.9 mm SL. Female without tubercles.
Scales cycloid. Lateral line completely pored, with 36(3), 37(13), 38*(11) or 39(1) perforated scales. Longitudinal scale rows between lateral line and dorsal-fin origin 4½(1), 5(2), 5½*(27) or 6½(1); longitudinal scale rows between lateral line and pelvic-fin origin 4(3), 5*(23) or 6(6); single row of 4(1), 5(4), 6(6), 7(8), 8(5) or 9*(5) scales on base of anteriormost anal-fin rays; circumpeduncular scales 14*(30); axillary scale present on pelvic-fin insertion.
Pectoral-fin rays I,9(1), i,10(1), i,10,i(11), i,11(2), i,11,i*(10), i,12(2) or i,12,i(3); tip of pectoral-fin not reaching pelvic-fin origin; pelvic-fin rays i,5,i(1), i,6,i(24) or i,7*(5); dorsal-fin rays ii,8*(30); first unbranched ray about one-half length of second unbranched ray; first branched ray longer than second unbranched ray; distal margin of dorsal-fin slightly rounded; anal-fin rays iii,16(3), 17*(19), 18(7) or 19(1); anal-fin insertion at vertical through insertion of last dorsal-fin ray or posterior to it; adipose-fin present, its insertion posterior to or at vertical through insertion of last anal-fin ray; caudal-fin principal rays i,17,i*(30); caudal-fin lobes rounded, equally sized.
Dorsal procurrent caudal-fin rays 12(2); ventral procurrent caudal-fin rays 11(2); total vertebrae 37(2); abdominal vertebrae 18(2); caudal vertebrae 19(2); fifth to seventeenth vertebrae type A; eighteenth vertebra type B; nineteenth to thirty-seventh vertebra type D; ribs 13(2), on fifth through seventeenth vertebra; supraneurals 5(2), between fourth and ninth vertebra; dorsal-fin pterygiophores 9(2), between eleventh through nineteenth vertebra; anal-fin pterygiophores 17(2), between nineteenth through twenty-seventh vertebra; two rows of gill rakers in arches 1–4, one row in fifth arch (only external); external rakers on first arch 17 (0, 1, 6, 0, 8, 0, 2); internal rakers on first arch 10 (0, 0, 6, 0, 4, 0, 0); external rakers on second arch 15 (0, 1, 5, 1, 6, 0, 2); internal rakers on second arch 12 (0, 0, 6, 0, 6, 0, 0); external rakers on third arch 14 (0, 1, 5, 1, 6, 0, 1); internal rakers on third arch 13 (0, 0, 5, 0, 8, 0, 0); external rakers on fourth arch 14 (0, 0, 6, 0, 8, 0, 0); internal rakers on fourth arch 7 (0, 0, 0, 0, 7, 0, 0); rakers on fifth arch 8 (only external).
Coloration in alcohol. Ground color yellow to dark-brown (Fig. 1). Dorsal portion of head with dark brown coloration from tip of snout to posterior margin of supraoccipital, extending posteriorly as dorsal band to end of caudal peduncle; great concentration of melanophores around orbit and along posterior margin of maxilla; fewer melanophores scattered on infraorbitals, interopercle and opercle, and anterior portion of lower jaw; melanophores also on distal margin of scales, more concentrated on scales above lateral line, forming reticulated pattern; vertically elongated black humeral spot across second to fourth lateral line scale, reaching three scale rows above and one scale row below lateral line, tapering downward. Second, inconspicuous humeral spot separated from first spot by two scales; dark midlateral stripe from second humeral spot to end of caudal peduncle; caudal-fin with narrower stripe along median rays and melanophores concentrated on middle portion of lobes; dorsal and anal fins with melanophores concentrated on distal border; first unbranched dorsal-fin ray completely covered by melanophores; pectoral, pelvic and adipose fins with few scattered melanophores.
Sexual dimorphism. Pelvic-fin slightly longer and more rounded in males (Tab. 1), covering completely urogenital opening in ventral view, and usually reaching anal-fin origin; pelvic-fin slightly shorter and more pointed in females, not covering urogenital opening in ventral view (Figs. 4C–D). Distal border of anal-fin straight in males, slightly concave in females (Figs. 4A–B). Anal-fin base slightly longer in males, pre-anal distance slightly longer in females (Tab. 1). Sexually mature males with bony hooks on anal and pelvic fins; anal-fin hooks on last unbranched ray through fifth to ninth branched ray; pelvic-fin hooks on all rays except first; bony hooks absent in females. Sexually mature males with breeding tubercles along dorsal and lateral portion of head, and on distal border of scales (Fig. 3A). Tubercles absent in females or, if present, few and concentrated only on distal border of scales.
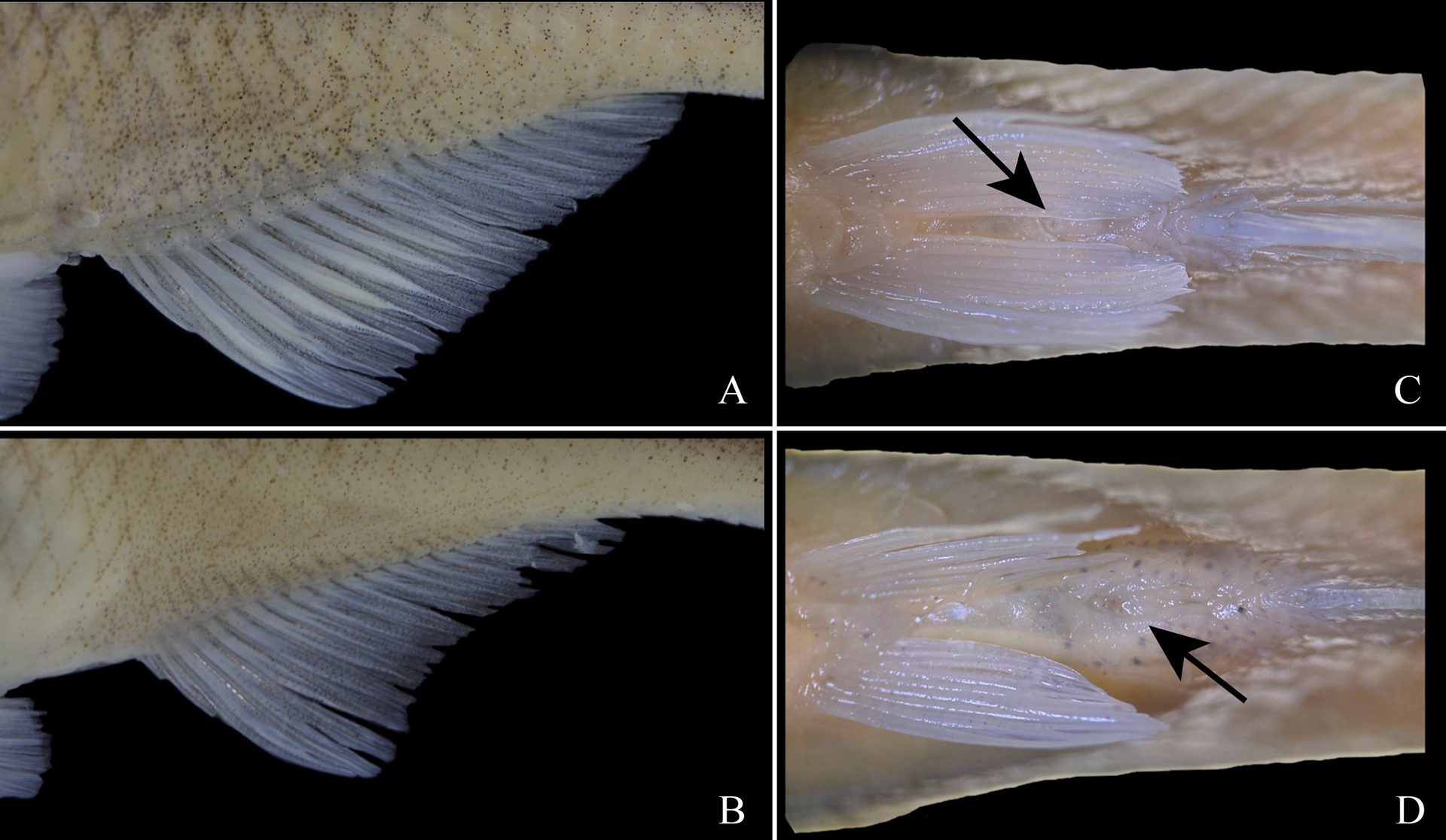
FIGURE 4| Sexual dimorphism in Bryconamericus misei. A. In males, the distal margin of the anal-fin is straight. B. In females, the distal margin of the anal-fin is slightly concave. C. In males, the pelvic-fin reaches distinctly past the urogenital opening. D. In females, the pelvic-fin reached at most the urogenital opening. A, C. NUP 24150, holotype, 54.8 mm SL. B, D. NUP 24149, paratype, 54.9 mm SL.
Geographical distribution. Bryconamericus misei is known from the rio Bonito and from two small tributaries to the rio Capivara, in the rio Piquiri basin; from the rio Laranjeiras, a tributary to the rio Formoso, in the rio Ivaí basin; and from the rio Apucarana, a tributary to the rio Tibagi (Fig. 5).
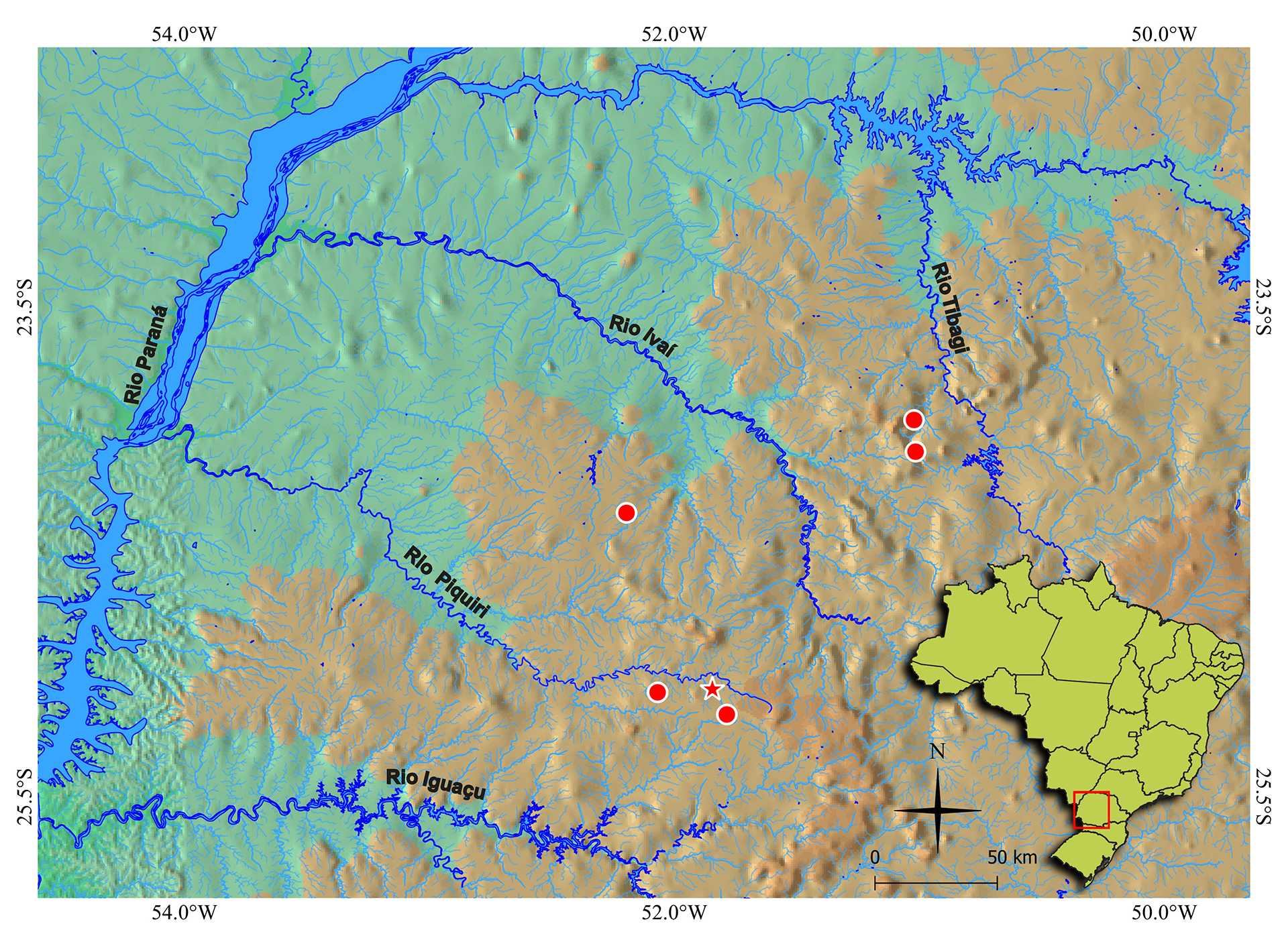
FIGURE 5| Geographic distribution of Bryconamericus misei (red marks). The star represents the type-locality. The red rectangle on detail indicates the position of the map in relation to Brazilian borders.
Ecological notes. At the sampling localities of Bryconamericus misei, the two tributaries to the rio Capivara, rio Piquiri basin (Figs. 6A–B) are about 1–2 m wide and 0.2 m deep, and lie about 800–900 m a.s.l. One of these (Fig. 6A) is the type-locality. The rio Laranjeiras, rio Ivaí basin (Fig. 6C) is about 4 m wide and 0.3 m deep, and lies about 498 m a.s.l. at the sampling location. The bottom of both streams is composed of small stones and pebbles, and some stretches with sandy bottom. The vegetation is mainly shrubs with some grasses, and stretches of the river are protected by canopies of small trees.

FIGURE 6| Sampling sites of Bryconamericus misei. A. Type-locality. B. Another unnamed stream, tributary to the rio Capivara, rio Piquiri basin. C. Rio Laranjeiras, a tributary to the rio Formoso, rio Ivaí basin.
Etymology. The specific name misei is a patronymic, given in honor of Fábio Teruo Mise, for his contributions to the ichthyological education of THP and for collecting part of the type-specimens of Bryconamericus misei. A noun in a genitive case.
Conservation status. Bryconamericus misei is known from tributaries to the Piquiri, Ivaí and Tibagi rivers and was collected from six different sites distributed along these basins. No threats to this species have been detected, therefore it has been classified as Least Concern (LC) according to the International Union for Conservation of Nature (IUCN) criteria and categories (IUCN Standards and Petitions Committee, 2022).
Discussion
Ideally, the classification of any given species should be supported by a phylogenetic analysis, preferably including molecular data. However, that is not always possible, even for newly described species, and we have to use pre-cladistic classificatory schemes, such as that of Eigenmann (1927), which allow us to assign a species to a given genus based on combination of character states. Given that the phylogenetic position of Bryconamericus misei is still unknown, its generic allocation is justified by the possession of the character combination established by Eigenmann (1927) and modified by Vari, Siebert (1990) for Bryconamericus.
The subterminal mouth with aligned teeth in the outer row of the premaxilla and dentary teeth decreasing more regularly in size in an anteroposterior sense places Bryconamericus misei apart from Bryconamericus sensu stricto, and closer to ‘B.’ agna, ‘B.’ ecai, ‘B.’ eigenmanni, ‘B.’ iheringii, ‘B.’ ikaa, ‘B.’ lambari, ‘B.’ patriciae, ‘B.’ rubropictus, ‘B.’ sylvicola, ‘B.’ uporas and ‘B.’ ytu, as well as to Hypobrycon and Odontostoechus. The phylogenetic relatedness of these species is confirmed by Mirande (2019), who shows that the “Nantis clade”, “Hypobrycon clade”, Odontostoechus lethostigmus and ‘B.’ microcephalus (hereafter referred to collectively as the “Odontostoechus clade”) are relatively distant from the clade containing the type-species Bryconamericus exodon. This relationship is also reflected in morphology, as the species included in the “Odontostoechus clade” differ radically from Bryconamericus exodon in many aspects, such as the general shape of the body and the pattern of dentition. Thus, species in the “Odontostoechus clade” must be transferred to another genus (or other genera) eventually. Odontostoechus is an eligible genus-level name for that assemblage, although the maintenance of Hypobrycon and Nantis as valid genera cannot be rejected as of yet. Nonetheless, much of its alpha taxonomy and phylogeny of the “Odontostoechus clade” needs clarification before such an arrangement involving the description of additional genera can be proposed.
Most of the species from the Northern South America that remain in Bryconamericus due to the lack of proper phylogenetic investigation differ from B. misei and similar species by a clearly terminal mouth. The few exceptions are ‘B.’ bolivianus Pearson, 1924, ‘B.’ grosvenori Eigenmann, 1927 (synonym of ‘B.’ bolivianus), and ‘B.’ pinnavittatus Dagosta & Netto-Ferreira, 2015. ‘Bryconamericus’ bolivianus has 15–16 total anal-fin rays and teeth of the outer premaxillary row about as large as those in the inner series (Pearson, 1924), which, along with its general appearance and geographic distribution suggests that in reality it could be a species of Ceratobranchia. ‘Bryconamericus’ grosvenori is very similar to female Attonitus irisae Vari & Ortega, 2000 (both species are described from Peru), with whom it shares similar dentition (especially the slender, tricuspid teeth), number of anal-fin rays and scales, and coloration. Dagosta, Netto-Ferreira (2015) investigated the phylogenetic position of ‘B.’ pinnavittatus based on morphological characters, recovering the species as sister to Bryconacidnus pectinatus Vari & Siebert, 1990. Whereas the allocation of the latter in Bryconacidnus Myers, 1929 by Thomaz et al. (2015) is questionable, there is no evidence that B. pectinatus or ‘B.’ pinnavittatus is closely related to B. exodon or other species from the Southern Neotropics.
Five species of Bryconamericus have been recorded in the area of occurrence of Bryconamericus misei: ‘B.’aff. iheringii (Fig. 7), ‘B.’ coeruleus, B. aff. stramineus,‘B.’ turiuba and the non-native B. exodon. Bryconamericus misei can be easily distinguished from all those species by the body depth (27.8–31.8% SL), from all but ‘B.’aff. iheringii by teeth aligned in the outer row of premaxilla, and still differs from ‘B.’aff. iheringii by having a smaller orbit diameter (28.1–33.9 vs. 34.8–40.9% HL in ‘B.’aff. iheringii), and also by having breeding tubercles in sexually mature males, a character absent in individuals attributed to ‘B.’aff. iheringii. The fact that B. misei was overlooked by previous inventories of the Ivaí and Piquiri ichthyofauna (Frota et al., 2016; Reis et al., 2020) reflects its overall similarity with ‘B.’aff. iheringii, but also the fact that even in heavily sampled driver basins undescribed species may remain unknown until in-depth taxonomic studies are performed.

FIGURE 7 | Lateral view of ‘Bryconamericus’aff. iheringii from the upper rio Paraná. A. NUP 16083, 55.9 mm SL, male, rio das Antas, rio Branco do Ivaí, 24°12’36”S 51°22’15”W, Paraná State, Brazil; B. NUP 16083, 54.6 mm SL, female; C. NUP 1441, 24.8 mm SL, rio Abelha, 23°36’01”S 52°27’30”W, Jussara, Paraná State, Brazil.
Among the other congeners, the species most similar to Bryconamericus misei are ‘B.’ microcephalus. That speciesis known from the Ribeira do Iguape basin, which has a high degree of isolation relative to drainages in the Inland Slope of State of Paraná (Reis et al., 2020). Indeed, the Ivaí, Piquiri and Tibagi sub-ecoregions share only 6.5–6.9% of their ichthyofaunas with the Ribeira de Iguape ecoregion (Reis et al., 2020:478–79, tabs. 4–5). It is likely that ‘B.’ microcephalus and B. misei are closely related, due to their phenotypic similarity and biogeographical proximity, but phylogenetic approaches are needed to provide a more conclusive answer in this regard. Furthermore, ‘B.’ microcephalus differs from B. misei by having a smaller body depth (23.5–27.7 vs. 27.8–31.8% SL in B. misei), a larger orbital diameter (38.4–41.6 vs. 28.1–33.9% HL in B. misei), and by having unpigmented fins (Bizerril, Peres-Neto, 1995), while B. misei has pigments in all fins.
The geographic distribution of Bryconamericus misei, restricted to the rio Piquiri, Ivaí and Tibagi basins, is similar to ‘B.’ coeruleus, which may indicate that those basins have been exchanging fauna via drainage rearrangement. Other species with a similar distribution are Apareiodon vladii Pavanelli, 2006 and Planaltina kaingang Deprá, Graça, Pavanelli, Avelino & Oliveira, 2018 (Reis et al., 2020), although they only occur in the Piquiri and Ivaí basins.
Comments on breeding tubercles and sexual dimorphism. Breeding tubercles are epidermal structures perceived as tiny white bumps on the body surface of preserved fish (Wiley, Collette, 1970). Their function is still not completely clear, but according to the same authors, the tubercles may facilitate body contact between the sexes during spawning and stimulate females during breeding.
In Bryconamericus misei, the tubercles occur in both sexes, but in different sizes and body regions. Males have well-developed tubercles on the dorsal and lateral portions of the head, and slightly less developed ones on the distal border of the scales, both on the dorsal and ventral portions of the body. In females, when present, the tubercles are smaller and restricted to the distal border of the scales, i.e., they are absent from the head. No tubercles were found on the fins in either sex. Tubercles and fin hooks in B. misei were found only in sexually mature specimens over 34 mm SL captured during the reproductive period, whereas sexually dimorphic character states related to the shape of the anal and pelvic fins were observed in adult specimens at gonadal resting stage and in juvenile specimens (smaller than 34 mm SL). Males in gonadal resting were collected in March and August. This indicates that tubercles and fin hooks are reabsorbed at the end of the reproductive period, as in Knodus nuptialis Menezes & Marinho, 2019.
The presence of breeding tubercles has been discussed in Wiley, Collette (1970) and revised in Menezes, Marinho (2019), who presented an updated table with the species that have the tubercles. According to these latter authors, the presence and distribution of tubercles may vary intra and interspecifically. Two other species of the “Odontostoechus clade” are described as having breeding tubercles: ‘B’. microcephalus and Hypobrycon poi. These two species have tubercles on the fins of sexually mature males, instead of on the head and scales. This tubercle distribution is similar to that found in the Stethaprioninae Deuterodon iguape Eigenmann, 1907, but differs from that of Bryconamericus misei and other characids, such as Creagrutus guanes Torres-Mejia & Vari, 2005, Knodus nuptialis, Astyanax scabripinnis (Jenyns, 1842) (see Menezes, Marinho, 2019), and Astyanax rupestris Zanata, Burger & Camelier, 2018, which have tubercles on the head and scales in both sexes, and Eretmobrycon emperador (Eigenmann & Ogle, 1907) (see Meek, Hildebrand, 1916), Astyanax aramburui Protogino, Miquelarena & López, 2006 and Piabina thomasi (Fowler, 1940) (see Protogino et al.,2006), which have tubercles on the head and scales, but only in male specimens. The presence of breeding tubercles was also recorded in other groups of fishes, in some species of Parodontidae, Lebiasinidae and Distichodontidae (Pavanelli, Britski, 2003; Vari, Ferraris, 2004; Pavanelli, 2006; Netto-Ferreira et al., 2011). This difference in the distribution of tubercles among different groups of fishes suggests that these structures evolved independently in each of these groups, as already mentioned by Wiley, Collette (1970).
Other characters associated with sexual dimorphism are a small difference in pre-anal distance, which is on average slightly greater in females, and greater anal-fin base length and pelvic-fin length in males. Regarding the pre-anal distance and anal-fin base length, there seems to be a trade-off between the abdominal and caudal regions, which is probably associated with an increase in the size of the abdominal cavity in females to fit the oocytes during the reproductive period. Other female characids seem to obtain an increase in mass devoted to reproduction through higher standard length or body depth (Menezes, Marinho, 2019; Teixeira et al.,2020).
Comparative material examined. Rio Uruguai basin. ‘Bryconamericus’ agna: NUP 24157, 7, 40.7–51.9 mm SL. ‘Bryconamericus’ patriciae: MCP 19615, 1, 47.6 mm SL, holotype; MCP 50013, 5, 60.5–70.8 mm SL. ‘Bryconamericus’ uporas: MCP 50912, 7, 33.9–52.4 mm SL. ‘Bryconamericus’ ytu: UFRGS 28406, 12 of 38, 41.2–72.0 mm SL. Hypobrycon leptorhynchus: MCP 18862, 1, 38.0 mm SL, holotype; NUP 18115, 1, 44.3 mm SL. Hypobrycon maromba: MCP 15757, 1, 43.0 mm SL, holotype. Hypobrycon poi: MCP 28164, 1, 51.6 mm SL, paratype. Laguna dos Patos basin. ‘Bryconamericus’ ecai: MCP 19608, 1, 60.6 mm SL, holotype. MCP 17494, 76, 53.8–76.2 mm SL, paratypes. ‘Bryconamericus’ iheringii: UFRGS 23692, 20 of 55, 44.9–57.2 mm SL; UFRGS 18191, 30 of 128, 43.3–61.2 mm SL. ‘Bryconamericus’ lambari: MCP 15448, 1, 55.9 mm SL, holotype; MCP 26057, 25 of 60, 40.2–50.4 mm SL. Upper rio Paraná basin. ‘Bryconamericus’aff. iheringii: NUP 1441, 10, 46.5–58.2 mm SL; NUP 16083, 10, 44.2–54.4 mm SL; NUP 20513, 77, 23.5–48.7 mm SL. ‘Bryconamericus’ coeruleus: NUP 3092, 8, 58.5 mm SL, paratypes; NUP 24156, 12, 48.5–63.3 mm SL. Bryconamericus exodon: NUP 10338, 11, 26.6–37.5 mm SL; NUP 16800, 10, 27.9–40.1 mm SL. ‘Bryconamericus’ turiuba: MCP 29414, 1, 61.1 mm SL, holotype; MCP 29073, 14, 40.4–54.3 mm SL, paratypes. Rio Iguaçu basin. ‘Bryconamericus’ ikaa: NUP 11845, 1, 49.7 mm SL; NUP 4128, 29, 28.9–51.4 mm SL; NUP 10789, 10, 36.1–51.5 mm SL. ‘Bryconamericus’ pyahu: NUP 12089, 1, 42.7 mm SL; NUP 7307, 3, 35.7–42.8 mm SL; NUP 7316, 2, 39.0 mm SL. Ribeira de Iguape basin. ‘Bryconamericus’ microcephalus: MZUSP 80013; NUP 20163, 5, 32.2–50.7 mm SL; NUP 20171, 3, 44.5–47.2 mm SL; NUP 17420, 5, 27.2–44.2 mm SL; NUP 17413, 10, 44.5–53.2 mm SL. ‘Bryconamericus’ tenuis: MZUSP 80226.
Acknowledgments
We would like to thank Fábio Teruo Mise for collecting and donating the material described here, and we would also like to thank Weferson J. Graça, Francisco A. Teixeira, Wladimir M. Domingues, Rodrigo J. Graça and Augusto Frota (NUP) for collecting material and for providing photos of the sampling sites. We are thankful to Fernando C. Jerep, Oscar A. Shibatta, and José L. O. Birindelli (MZUEL), for the loan of material and for their hospitality during the visit to the collection. We would also like to thank Marcos Mirande (Fundación Miguel Lillo) and another anonymous reviewer who contributed greatly to improving the manuscript. THP is grateful to the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES Proc. #88887.627795/2021–00). GCD and CSP are grateful to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq grant #151115/2022–2 and #308777/2019-0, respectively).
References
Berg C. Communicaciones ictiológicas. Comun Mus Nac B Aires. 1901; 1(9):293–311.
Bizerril CRSF, Peres-Neto PR. Redescription of Bryconamericus microcephalus (Ribeiro, 1908) and description of a new species of Bryconamericus (Characidae, Tetragonopterinae) from eastern Brazil. Comun Mus Ciên Tecnol PUCRS, Sér Zool. 1995; 8:13–25.
Braga L. Redescription of Bryconamericus rubropictus (Berg) n. comb. (Ostariophysi: Characidae) and reference to its secondary sexual dimorphism. Rev Mus Arg Cienc Nat Bernardino Rivadavia. 2000; 2(2):145–50.
Cavalli D, Frota A, Lira AD, Gubiani EA, Margarido VP, Graça WJ. Update on the ichthyofauna of the Piquiri River basin, Paraná, Brazil: a conservation priority area. Biota Neotrop. 2018; 18(2):e20170350. http://dx.doi.org/10.1590/1676-0611-BN-2017-0350
Dagosta FCP, Netto-Ferreira AL. New species of Bryconamericus Eigenmann (Characiformes: Characidae) from the rio Teles Pires, rio Tapajós basin, central Brazil. Zootaxa. 2015; 3911(3):433–42. https://doi.org/10.11646/zootaxa.3911.3.9
Deprá GC, Ota RR, Vitorino Júnior OB, Ferreira KM. Two new species of Knodus (Characidae: Steverdiinae) from the upper rio Tocantins basin, with evidence of ontogenetic meristic changes. Neotrop Ichthyol. 2021; 19(1): e200106. https://doi.org/10.1590/1982-0224-2020-0106
Eigenmann CH. The American Characidae. Mem Mus Comp Zool. 1927; 43:358–95.
Fink WL. A new genus and species of characid fish from the Bayano River basin, Panamá (Pisces: Cypriniformes). Proc Biol Soc Wash. 1976; 88(30):331–44.
Fink WL, Weitzman SH. The so-called cheirodontin fishes of Central America with descriptions of two new species (Pisces: Characidae). Smithson Contr Zool. 1974; (172):1–46. https://doi.org/10.5479/si.00810282.172
Fricke R, Eschmeyer WN, Van der Laan R. Eschmeyer’s catalog of fishes: Genera, species, references [Internet]. San Francisco: California Academy of Science; 2023. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Frota A, Deprá GC, Petenucci LM, Graça WJ. Inventory of the fish fauna from Ivaí River basin, Paraná State, Brazil. Biota Neotrop. 2016; 16(3):e20150151. https://doi.org/10.1590/1676-0611-BN-2015-0151
García-Melo JE, Oliveira C, Silva GJC, Ochoa-Orrego LE, Pereira LHG, Maldonado-Ocampo JA. Species delimitation of Neotropical characins (Stevardiinae): Implications for taxonomy of complex groups. PLoS ONE. 2019; 14(6):e0216786. https://doi.org/10.1371/journal.pone.0216786
Grande T, Young B. The ontogeny and homology of the Weberian apparatus in the zebrafish Danio rerio (Ostariophysi: Cypriniformes). Zool J Linn. 2004; 140(2):241–54. https://doi.org/10.1111/j.1096-3642.2003.00097.x
Jerep FC, Shibatta OA. A new species of Bryconamericus (Characidae: Stevardiinae: Diapomini) from the upper rio Paraná basin, Brazil. Neotrop Ichthyol. 2017; 15(3):e170028. https://doi.org/10.1590/1982-0224-20170028
Malabarba MCSL, Malabarba LR. Hypobrycon maromba, a new genus and species of Characiform fish from the upper rio Uruguay, Brazil (Ostariophysi: Characidae). Ichthyol Explor Freshw. 1994; 5(1):19–24.
Malabarba LR, Kindel A. A new species of the genus Bryconamericus Eigenmann, 1907 from Southern Brazil (Ostariophysi: Characidae). Proc Biol Soc Wash. 1995; 108(4):679–86.
Meek SE, Hildebrand SF. The fishes of the fresh waters of Panama. Field Mus Nat Hist. Zool Ser. 1916; 10(15):217–374.
Menezes NA, Marinho MMF. A new species of Knodus Eigenmann (Characiformes: Characidae: Stevardiinae) with comments on nuptial tubercles ang gill gland in characiform fishes. PLoS ONE. 2019; 14(7):e0217915. https://doi.org/10.1371/journal.pone.0217915
Miquelarena AMLC, Protogino RF, HL López. A new species of Bryconamericus (Characiformes: Characidae) from the Cuña-Pirú creek in northeastern Argentina, with comments on accompanying fishes. Aqua. 2002; 6(2):69–82.
Mirande JM, Aguilera G, Azpelicueta MM. A new genus and species of small characid (Ostariophysi, Characidae) from the upper río Bermejo basin, northwestern Argentina. Rev Suisse Zool. 2004; 111(4):715–28.
Mirande JM. Morphology, molecules and the phylogeny of Characidae (Teleostei, Characiformes). Cladistics. 2019; 35(3):282–300. https://doi.org/10.1111/cla.12345
Netto-Ferreira AL, Oyakawa OT, Zuanon J, Nolasco JC. Lebiasina yepezi, a new Lebiasininae (Characiformes: Lebiasinidae) from the Serra Parima-Tapirapecó mountains. Neotrop Ichthyol. 2011; 9(4):767–75. https://doi.org/10.1590/S1679-62252011000400008
Pavanelli CS. New species of Apareiodon (Teleostei: Characiformes: Parodontidae) from the rio Piquiri, upper rio Paraná basin, Brazil. Copeia. 2006; 2006(1)1:89–95. https://doi.org/10.1643/0045-8511(2006)006[0089:NSOATC]2.0.CO;2
Pavanelli CS, Britski HA. Apareiodon Eigenmann, 1916 (Teleostei, Characiformes), from the Tocantins-Araguaia basin, with description of three new species. Copeia. 2003; 2003(2):337–48. https://doi.org/10.1643/0045-8511(2003)003[0337:AETCFT]2.0.CO;2
Pearson NE. The fishes of the eastern slope of the Andes. I. The fishes of the rio Beni basin, Bolivia, collected by the Mulford expedition. Indiana University Studies. 1924; 11(64):1–83.
Protogino LC, Amalia MM, López HL. A new species of Astyanax (Teleostei: Characiformes: Characidae), with breeding tubercles, from the Paraná and Uruguay river basins. Zootaxa. 2006; 1297(1):1–16. https://doi.org/10.11646/zootaxa.1297.1.1
Reis RB, Frota A, Deprá GC, Ota RR, Graça WJ. Freshwater fishes of Paraná State, Brazil: an annotated list, with comments on biogeographic patterns, threats, and future perspectives. Zootaxa. 2020; 4868(4):451–94. https://doi.org/10.11646/zootaxa.4868.4.1
Sabaj MH. Codes for Natural History Collections in Ichthyology and Herpetology (online supplement). American Society of Ichthyologists and Herpetologists, Washington, DC. 2022. Available on: https://asih.org
Taylor WR, Van Dyke GC. Revised procedures for clearing and staining small fishes and other vertebrates for bone and cartilage study. Cybium. 1985; 9(2):107–20.
Teixeira TF, Dutra GM, Penido IS, Santos SA, Pessali TC. A new species of Hyphessobrycon (Characiformes, Characidae) from the upper rio Jequitinhonha basin, Minas Gerais, Brazil, with comments on morphological similarities with Stethaprioninae species from headwater environments. J Fish Biol. 2020; 96(1):140–53. https://doi.org/10.1111/jfb.14198
Thomaz AT, Arcila D, Ortí G, Malabarba LR. Molecular phylogeny of the subfamily Stevardiinae Gill, 1858 (Characiformes: Characidae): classification and the evolution of reproductive traits. BMC Evol Biol. 2015; 15:146. https://doi.org/10.1186/s12862-015-0403-4
Torres-Mejia M, Vari RP. New species of Creagrutus (Teleostei: Characiformes: Characidae) from the Río Magdalena basin, Colombia. Copeia. 2004; 2004(4):812–17. https://doi.org/10.1643/0045-8511(2005)005[0812:NSOCTC]2.0.CO;2
Vari RP, Ferraris CJ. A new species of Nannocharax (Characiformes: Distichodontidae) from Cameroon, with the description of contact organs and breeding tubercles in the genus. Proc Biol Soc Wash. 2004; 117(4):551–63.
Vari RP, Siebert DJ. A new, unusually sexually dimorphic species of Bryconamericus (Pisces: Ostariophysi: Characidae) from the Peruvian Amazon. Proc Biol Soc Wash. 1990; 103(3):516–24.
Wiley ML, Collette BB. Breeding tubercles and contact organs in fishes: their occurence, structure, and significance. Bull Carnegie Mus Nat. 1970; 143:143–216.
Authors

![]() Thiago Henrique Pedroso1
Thiago Henrique Pedroso1 ![]() ,
, ![]() Gabriel de Carvalho Deprá1 and
Gabriel de Carvalho Deprá1 and ![]() Carla Simone Pavanelli1,2
Carla Simone Pavanelli1,2
[1] Programa de Pós-Graduação em Ecologia de Ambientes Aquáticos Continentais, Universidade Estadual de Maringá. Av. Colombo, 5790, 87020-900 Maringá, PR, Brazil. (THP) thiagohpedroso@gmail.com (corresponding author), (GCD) gabrieldepra@gmail.com.
[2] Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (Nupélia), Universidade Estadual de Maringá. Av. Colombo, 5790, 87020-900 Maringá, PR, Brazil. (CSP) carlasp@nupelia.uem.br.
Authors’ Contribution 

Thiago Henrique Pedroso: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Writing-original draft, Writing-review and editing.
Gabriel de Carvalho Deprá: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Writing-original draft, Writing-review and editing.
Carla Simone Pavanelli: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Writing-original draft, Writing-review and editing.
Ethical Statement
Not applicable.
Competing Interests
The author declares no competing interests.
How to cite this article
Pedroso TH, Deprá GC, Pavanelli CS. A new species of Bryconamericus (Characidae: Stevardiinae) with breeding tubercles from the upper rio Paraná basin. Neotrop Ichthyol. 2024; 22(1):e230049. https://doi.org/10.1590/1982-0224-2023-0049
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2024 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted December 19, 2023 by Priscila Camelier
Accepted December 19, 2023 by Priscila Camelier
![]() Submitted May 9, 2023
Submitted May 9, 2023
![]() Epub February 19, 2024
Epub February 19, 2024