![]() Luis Guillermo Constante-Pérez1,2,
Luis Guillermo Constante-Pérez1,2, ![]() Manuel Castillo-Rivera1
Manuel Castillo-Rivera1 ![]() and
and ![]() Alejandra Serrato-Díaz3
Alejandra Serrato-Díaz3
PDF: EN XML: EN | Cite this article
Abstract
Snappers are marine fishes, but juveniles of many species migrate to estuaries, using these systems as nursery areas. The purpose of this study was to know the environmental factors mainly related to the migration patterns of lutjanids in La Mancha lagoon inlet. During 19 months, 24-hour cycles were performed monthly, taking samples every two hours (442 samples). Environmental variables recorded in situ and with regional records such as rainfall, atmospheric temperature and day length were considered. Genetic barcoding (COI) was used to validate species identity. Significant differences were evaluated by PERMANOVA and a Canonical Correspondence Analysis was used to determine the importance of environmental variables. Six species were recorded and the most abundant showed significant differences among months, with migration patterns during the warm-rainy season. Day length, salinity and rainfall (two of them with regional incidence) were the variables significantly associated with the distribution of the species. The abundance of Lutjanus analis, L. jocu, and L. cyanopterus was inversely associated with the length of daylight and directly related to rainfall, while L. synagris and L. griseus showed segregation along a salinity gradient. Thus, migration patterns were mainly correlated with factors of regional coverage and greater seasonal influence.
Keywords: Atmospheric Temperature, Competition, Photoperiod, Water Level, Zooplankton.
Los pargos son peces marinos, pero los juveniles de muchas especies migran a los estuarios, utilizándolos como áreas de crianza. El propósito del estudio fue conocer los factores ambientales principalmente relacionados con los patrones de migración de lutjánidos en la boca de la laguna La Mancha. Durante 19 meses se realizaron ciclos de 24 horas, tomando muestras cada dos horas (442 muestras). Se consideraron variables ambientales registradas in situ y con registros regionales tales como lluvias, temperatura atmosférica y duración del día. Se utilizó código de barras genético (COI) para validar la identidad de las especies. Diferencias significativas fueron evaluadas por PERMANOVA y un Análisis de Correspondencia Canónica permitió determinar la importancia de las variables ambientales. Se registraron seis especies y las más abundantes mostraron diferencias significativas entre meses, con patrones de migración durante la época cálida-lluviosa. La duración del día, la salinidad y la precipitación (dos con incidencia regional) fueron las variables asociadas significativamente a la distribución de las especies. La abundancia de Lutjanus analis, L. jocu y L. cyanopterus estuvo inversamente asociada con la duración del día y directamente con las lluvias, mientras que L. synagris y L. griseus mostraron segregación a lo largo de un gradiente de salinidad. Así, los patrones de migración se correlacionaron principalmente con factores de cobertura regional y de mayor influencia estacional.
Palabras clave: Competencia, Efecto de Marea, Fotoperiodo, Temperatura Atmosférica, Zooplancton.
Introduction
Snappers (Lutjanidae) are marine fishes that occur in tropical and subtropical oceans, mostly bottom-associated, occurring from shallow inshore areas to depths of about 550 m, mainly over reefs or rocky outcrops. However, juvenile stages of some species enter estuaries and the lower reaches of rivers. They are mostly carnivores with nocturnal activity and spawning is also apparently at night (Anderson, 2002; McEachran, Fechhelm, 2005; Castillo-Rivera et al., 2020).
The Lutjanidae family includes about 110 species (Nelson et al., 2016), of which 16 species in six genera occur in the Gulf of Mexico (Anderson, 2002; McEachran, Fechhelm, 2005). Twelve of these species have been recorded on the continental shelf of Veracruz State in Mexico, although only seven of them have been recorded in estuarine systems (Lara-Domínguez et al., 2011).
In this sense, for a better evaluation of the diversity and temporal distribution patterns of fishes in an area, a greater sampling effort is necessary, increasing the number of months sampled, as well as sampling throughout the 24-h cycle. Indeed, in many studies of estuarine communities it has been observed that during the twilight or at night, a greater number of fish species is caught due to their foraging activity (Ley, Halliday, 2007; Castillo-Rivera et al., 2017). For this reason, surveys that also considered night and twilight samples can provide a better understanding of fish richness and abundance (Hagan, Able, 2008; Castillo-Rivera et al., 2017).
Seasonal and diel changes in the relative abundance of fish species may be influenced by constant fluctuations in environmental factors. Traditionally, environmental variables that are recorded in situ, such as water temperature and salinity, and dissolved oxygen levels, have been considered. However, environmental variables with regional rather than local coverage can also play an important role, as has been observed for currents (Sanvicente-Añorve et al., 2011; Castillo-Rivera et al., 2020), wind effects (Bruno et al., 2013, 2015; Souza et al., 2018), photoperiod (Capossela et al., 2013; Garcia, Pessanha, 2018), and rainfall (Bruno et al., 2013; Castillo-Rivera et al., 2017; Souza et al., 2018). Indeed, species abundance patterns at the local scale are likely to be influenced by environmental processes that operate at both the local and regional scales (Alonso Aller et al., 2014; Valesini et al., 2014; Hayduk et al., 2019).
Although snapper adults inhabit the marine environment, larvae and juveniles of many species migrate to estuaries, where they may spend their first life stages, using these systems as nursery areas (Anderson, 2002; Serrano et al., 2011; Lustosa-Costa et al., 2020). In this sense, the contribution of the current study is to know what are the main environmental factors related to the migration patterns of lutjanids in an estuary.
In this way, it was hypothesized that snapper migration patterns to estuarine systems may be more related to seasonal variations in environmental factors than to diel variations in these factors. Our main objective was, through a high-frequency temporal sampling, to determine the diversity of lutjanids, and to evaluate the relationship of environmental factors with the seasonal and diel distribution patterns of these species at the inlet of La Mancha lagoon.
Material and methods
Study area. La Mancha lagoon is located in Veracruz State, Mexico (19°33’55” –19°35’44”N and 96°22’41” – 96°23’39”W), in the southwestern Gulf of Mexico. It is a small system, with an area of ~1.6 km2 and bordered by mangrove forest. At its northeastern end, the system opens to the ocean via an inlet that discharges through a sand barrier, showing a seasonal pattern of open/close, which is regulated by the tides and freshwater discharge (Lara-Domínguez et al., 2006).
The region has a warm sub-humid climate (Köppen climate classification: Aw2), with mean monthly rainfall between 12.8 mm in February and 265.8 mm in August, including a warm-rainy season (mean values > 100 mm from June to October) and a dry season (mean values < 60 mm from November to May). In La Mancha lagoon, the abundance of zooplankton and chlorophyll a show two peaks: one during August–October and another in February–March (Contreras-Espinosa et al., 2005; Mecalco-Hernández et al., 2018).
Data collection. In a sandy habitat within the lagoon, for 19 months, 24-hour cycles of collection activities were carried out, taking samples every two hours, simultaneously, and monthly, at two sites, one at 325 m and the other at 225 m from the inlet system (a total of 442 samples) (Fig. 1). The samples were taken with a beach seine net (37 m long, 1.2 m deep and 1 cm mesh size). The net was pulled 50 m parallel to the shore and covered an approximate sampling area of 1,500 m2 (gear length x haul distance). Immediately after their capture, some fish individuals were processed and frozen for DNA analysis and the others were anesthetized (clove oil solution) and posteriorly fixed with 10% formalin and preserved with 70% ethanol. The individuals were identified according to Anderson (2002), counted, and measured (Total length – TL and Standard length – SL) in the laboratory. One specimen of each species was cataloged and deposited in the fish collection of the Department of Biology, Universidad Autónoma Metropolitana, Unidad Iztapalapa (UAMI-CI).

FIGURE 1| Geographical location of La Mancha lagoon and sampling points (black circles).
In each sampling, the time of day, water salinity (ATAGO S-10E Refractometer), water temperature, dissolved oxygen (YSI 550A DO Instrument), chlorophyll a (spectrophotometric method, SCOR, 1966), and zooplankton biomass (circular trawl with 150 µm mesh net and volumetric method) were recorded in situ. Additionally, the monthly averages of rainfall were considered, as well as the duration of daylight hours for each sampling day and the water level (tidal stage effect) for each sampling time, all recorded for the study period (regional data for the state from Veracruz). The monthly averages (corresponding to 60 years: 1951–2010) of the atmospheric temperature were also considered (SMN, 2020).
DNA analyses. Particularly for snappers, it should be noted that juveniles and young adults could be difficult to identify taxonomically, since most species share meristic and overlap in morphometrics. Therefore, the use of DNA barcodes has been suggested, especially when rare and common species coexist and share a similar appearance (Victor et al., 2009). Therefore, 47 specimens (including at least one specimen per species) were used for DNA analysis to validate species identity using genetic barcoding (mtDNA, partial sequence of cytochrome oxidase subunit I – COI). For each of these individuals, tissue samples collected from caudal fins were preserved in molecular grade 95% ethanol. The extraction, amplification, and sequencing protocols were in accordance with Castillo-Rivera et al. (2020). The sequence electropherogram was edited in MEGA-X and was compared with the database of BOLD Systems. A sequence of each registered snapper species was submitted to GenBank database.
Statistical analyses. For the most abundant species (> 10% of relative abundance), two-way univariate PERMANOVA was used to assess significant differences in the number and size of fish, with months and diel periods (day, twilight, and night) as fixed factors, followed by of posteriori pair-wise comparisons among factor levels. In the same way, the environmental variables recorded in situ were analyzed. PERMANOVA is a routine based on a resemblance measure, generally highly appropriate, because most ecological data (being counts of abundances of species) tend to be overdispersed, with a substantial proportion of zeros. This method is also robust because uses multiple random permutations to obtain P-values, thus normality and homogeneity of variances are directly implied by the permutation procedure (Anderson, 2001; Anderson et al., 2008). These analyses were performed using Euclidean distance (on square root transformed data), and permutated residuals under a reduced model, Type III (maximum permutations = 999). The square-root transformation is used most frequently with count data, which often follows a Poisson distribution (Gotelli, Ellison, 2004), as in the present case. These analyses were performed using PRIMER v7 (Clarke, Gorley, 2015).
For ordination purposes, according to Lepš, Šmilauer (2003), a Detrended Correspondence Analysis (DCA) was first calculated, which showed that the gradient lengths for the first two axes were 3.121 and 3.340 respectively. These values suggested that the use of unimodal ordination methods is appropriate. Thus, a Canonical Correspondence Analysis (CCA) was applied to the abundance matrix of all species (dependent set) and to the environmental data matrix (independent set). Rare species were downweighted according to ter Braak, Šmilauer (2002). CCA is an appropriate ordination technique designed to describe and visualize differential habitat preferences of species via an ordination diagram. Thus, this method explores the correspondence between species distribution and environmental factors. The significance of these factors was determined using 499 unrestricted Monte Carlo permutations. Correlations between environmental variables and species-derived sample scores (inter-set correlations) were used to determine correspondence between these data sets (ter Braak, Šmilauer, 2002). These analyses were performed using the package CANOCO ver. 4.5.
Results
Environmental variables that characterized the system are shown in Tab. 1. All of them showed differences between months and between diel periods (Tab. 2). Pair-wise comparisons showed that the significantly higher monthly values were for temperature during June–October, for salinity in November–December and July–August of the second year, and for dissolved oxygen during June–May and January–March of the second year. A significant pulse was reached for chlorophyll a in June–July and for zooplankton biomass in January (dominated by copepods) and August (dominated by decapod larvae). Diel variation shown that temperature, dissolved oxygen, and chlorophyll a presented significantly higher values during daylight hours, while salinity and zooplankton biomass shown the highest values at night and twilight. The inlet of the lagoon was closed during January–March and the tidal cycle was mainly diurnal with high tides at 04:00 h from March to August and at 16:00 h from September to February.
TABLE 1 | Mean values, standard deviations (SD), and range of environmental parameters of La Mancha lagoon, recorded throughout the 19 months of sampling.
Mean±SD | Range | |
Salinity | 22.53±9.36 | 5–40 |
Water temperature (°C) | 28.01±2.92 | 20.9–34.7 |
Dissolved oxygen (mg/L) | 5.38±1.12 | 2.1–8.8 |
Chlorophyll a (mg/m3) | 12.88±29.30 | 0.05–299.4 |
Zooplankton (mL/m3) | 7.38±10.11 | 0.0-49.0 |
Water level (tidal effect, cm) | 6.62±18.17 | -23.06–61.42 |
A total of 272 individuals of Lutjanus griseus (Linnaeus, 1758), 95 of L. synagris (Linnaeus, 1758), 64 of L. analis (Cuvier, 1828), 41 of L. jocu (Bloch & Schneider, 1801), seven of L. cyanopterus (Cuvier, 1828) and one of Ocyurus chrysurus (Bloch, 1791) were caught. The catalog numbers of each specimen deposited in the fish collection were: L. griseus (UAMI-CI 356), L. synagris (UAMI-CI 359), L. analis (UAMI-CI 352), L. jocu (UAMI-CI 357), L. cyanopterus (UAMI-CI 354) and O. chrysurus (UAMI-CI 361). The COI sequences of the 47 specimens selected for DNA analysis, which included at least one specimen per species (24 L. griseus, four L. synagris, 14 L. analis, three L. jocu, one L. cyanopterus, and one O. chrysurus), confirmed the identity of species. For these specimens, the overall genetic identities ranged from 99% to 100%. In addition, a DNA sequence of each species was submitted to the GenBank database under the following accession numbers: KU314762 for L. griseus, KU314765 for L. synagris, KU314763 for L. analis, KX119130 for L. jocu, KX119131 for L. cyanopterus and KU314764 for O. chrysurus.
TABLE 2 | Univariate two-way PERMANOVA to test the effects of months and diel period (daytime, twilight, and nighttime) on the in situ environmental variables studied. Significant P values are shown in bold.
Source
of variance | df | MS | Pseudo-F | P
(permuted) |
Salinity | ||||
Months | 18 | 1392.00 | 88.03 | 0.001 |
Diel | 2 | 69.18 | 4.375 | 0.023 |
Months x Diel | 36 | 60.27 | 3.811 | 0.001 |
Residual | 385 | 15.81 | ||
Water temperature | ||||
Months | 18 | 126.49 | 56.30 | 0.001 |
Diel | 2 | 18.66 | 8.306 | 0.001 |
Months x Diel | 36 | 1.812 | 0.806 | 0.761 |
Residual | 385 | 2.247 | ||
Dissolved oxygen | ||||
Months | 18 | 14.301 | 29.455 | 0.001 |
Diel | 2 | 2.184 | 4.498 | 0.013 |
Months x Diel | 36 | 0.855 | 1.761 | 0.007 |
Residual | 385 | 0.486 | ||
Chlorophyll a | ||||
Months | 18 | 1765.6 | 3.171 | 0.018 |
Diel | 2 | 4075.4 | 7.320 | 0.006 |
Months x Diel | 36 | 1243.0 | 2.232 | 0.022 |
Residual | 385 | 556.77 | ||
Zooplankton | ||||
Months | 18 | 1055.4 | 30.798 | 0.001 |
Diel | 2 | 151.38 | 4.417 | 0.014 |
Months x Diel | 36 | 101.04 | 2.949 | 0.001 |
Residual | 385 | 34.268 |
The monthly variation of the relative abundances showed that the three most abundant snapper species enter the system during the warm-rainy season, which presents the longest periods of daylight (June–October) (Fig. 2). This variation showed differences among the monthly mean values (Tab. 3). According to a posteriori pair-wise comparisons, the significantly highest monthly mean values of abundance throughout the 19 months were in July of the first year and October of the second year for L. griseus, while for L. synagris these were during July and August of the second year. For L. analis the significantly higher monthly mean values were in September and October of the second year. In this way, a segregation pattern was observed, in which L. griseus occurs mainly in June–August of the first year (when the mean salinity is less than 27) and L. synagris in June–August of the second year (when salinity mean is greater than 32). The main entry of L. analis was in September–October of the second year (mean salinity around 25) (Fig. 2A). For the three species, there was no interaction between the month and diel period factors, indicating independence of each factor. Day length was longer in June–July and rainfall peaked in August–September in both years, while during these periods mean salinity was higher in the second year (Fig. 2B).
TABLE 3 | Univariate two-way PERMANOVA to test the effects of months and diel period (daytime, twilight, and nighttime) on species abundance. Significant P values are shown in bold.
Source
of variance | df | MS | Pseudo-F | P
(permuted) |
Lutjanus griseus | ||||
Months | 18 | 2.287 | 6.039 | 0.001 |
Diel | 2 | 1.621 | 4.280 | 0.017 |
Months x Diel | 36 | 0.512 | 1.352 | 0.132 |
Residual | 385 | 0.379 | ||
Lutjanus synagris | ||||
Months | 18 | 0.686 | 3.898 | 0.002 |
Diel | 2 | 0.036 | 0.205 | 0.820 |
Months x Diel | 36 | 0.221 | 1.255 | 0.210 |
Residual | 385 | 0.176 | ||
Lutjanus analis | ||||
Months | 18 | 0.376 | 3.159 | 0.004 |
Diel | 2 | 0.013 | 0.109 | 0.919 |
Months x Diel | 36 | 0.145 | 1.218 | 0.183 |
Residual | 385 | 0.119 |
At the diel level, only L. griseus showed differences in abundance among diel periods with a pulse from 16:00 to 18:00 h (Fig. 3). Pair-wise comparisons showed that the mean values during the day and twilight hours were higher than those at night. By contrast, L. synagris and L. analis did not show differences among diel periods (Tab. 3). Individuals of L. cyanopterus and O. chrysurus were captured only during twilight and night hours.

FIGURE 2| Monthly variation of: A. Mean and standard error values of the three most abundant lutjanid species, and B. Values of the main environmental conditions studied in the study area.
The range and mean values of TL (mm) were: L. griseus (19–254, = 131.89), L. synagris (21–160, = 56.21), L. analis (21–172, = 64.85), L. jocu (19–254, = 131.89), L. cyanopterus (161–391, = 181.71), O. chrysurus (76.08). The three most abundant species showed significant differences in size among months (P’s = 0.001) and according to multiple comparison tests, the smaller individuals of L. griseus were recorded during October, while the smallest of L. synagris and L. analis were recorded in July and October. Regarding diel variation, only the length of L. synagris showed differences between diel periods (P = 0.001), with the larger individuals collected during the night and twilight hours.
The first two canonical axes were significant (P = 0.002 and P = 0.018) and jointly explained 88.3% in “species-environment relation” of the total cumulative variance (Fig. 4). The species-environment correlation coefficients of these two axes were 0.567 and 0.454, respectively. In the tri-plot of this analysis, the length and direction of arrows indicate the relative importance and direction that each factor has on the species ordination. As inlet state and diel period (daytime, twilight, and nighttime) are ordinal variables, the corresponding vector indicates, in the direction of the arrows, a preference for open inlet and day period, respectively. According to significance tests derived from CCA, only daylight hours of each diel cycle, water salinity and mean monthly rainfall were the environmental variables related to the species distribution (Tab. 4). These environmental factors also showed high values of inter-set correlations. According to ter Braak, Šmilauer (2002), the variance inflation factors (VIF) showed that the measurements are not strongly correlated.
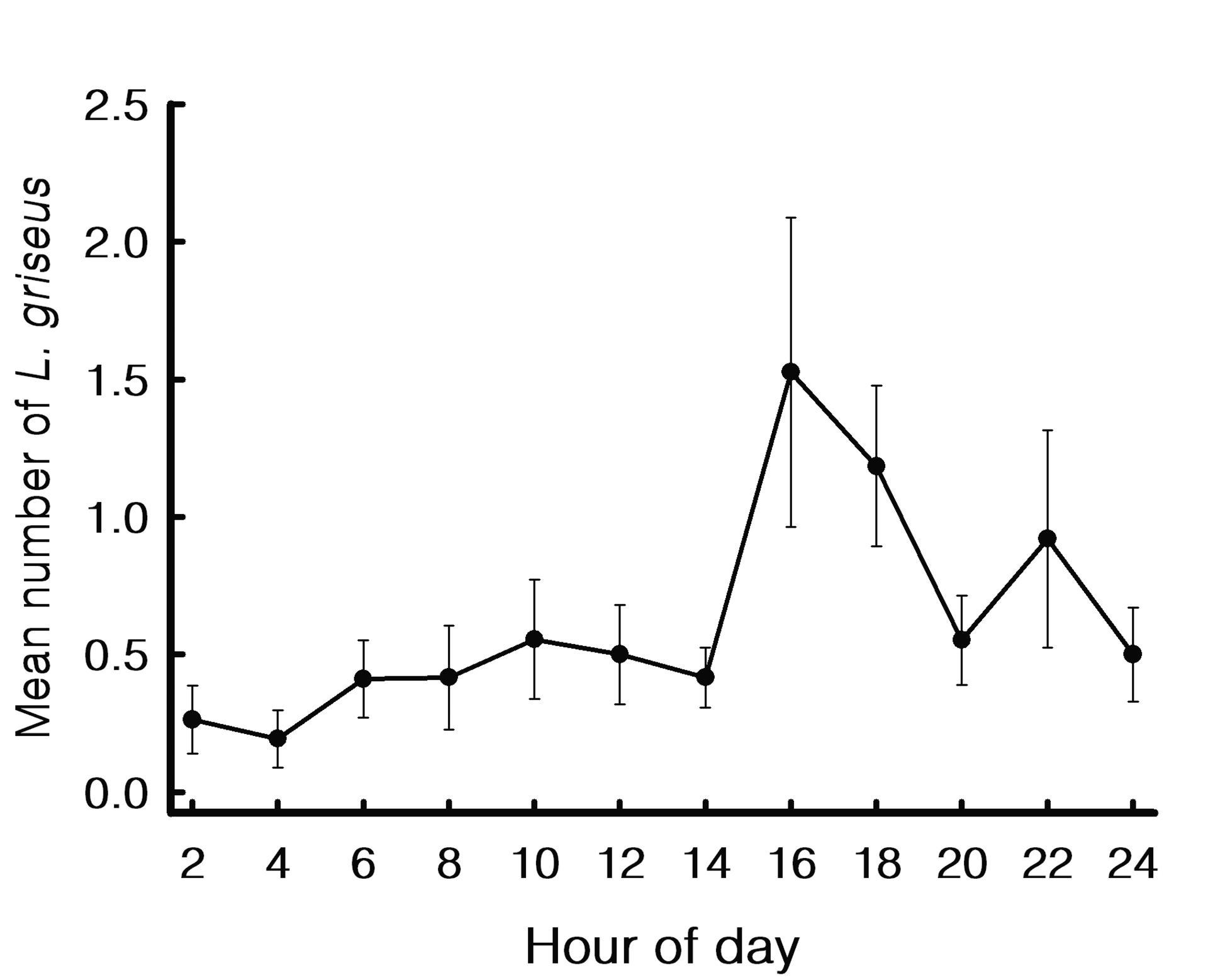
FIGURE 3| Mean number and standard error of Lutjanus griseus abundance by hour of day at the inlet of La Mancha lagoon.
Although zooplankton biomass, mean atmospheric temperature and water level show relatively high correlations with the first or second axis, these variables did not have a significant effect on species distribution. Furthermore, chlorophyll a, diel effect, water temperature and dissolved oxygen had a negligible importance on the species distribution (P’s > 0.50).
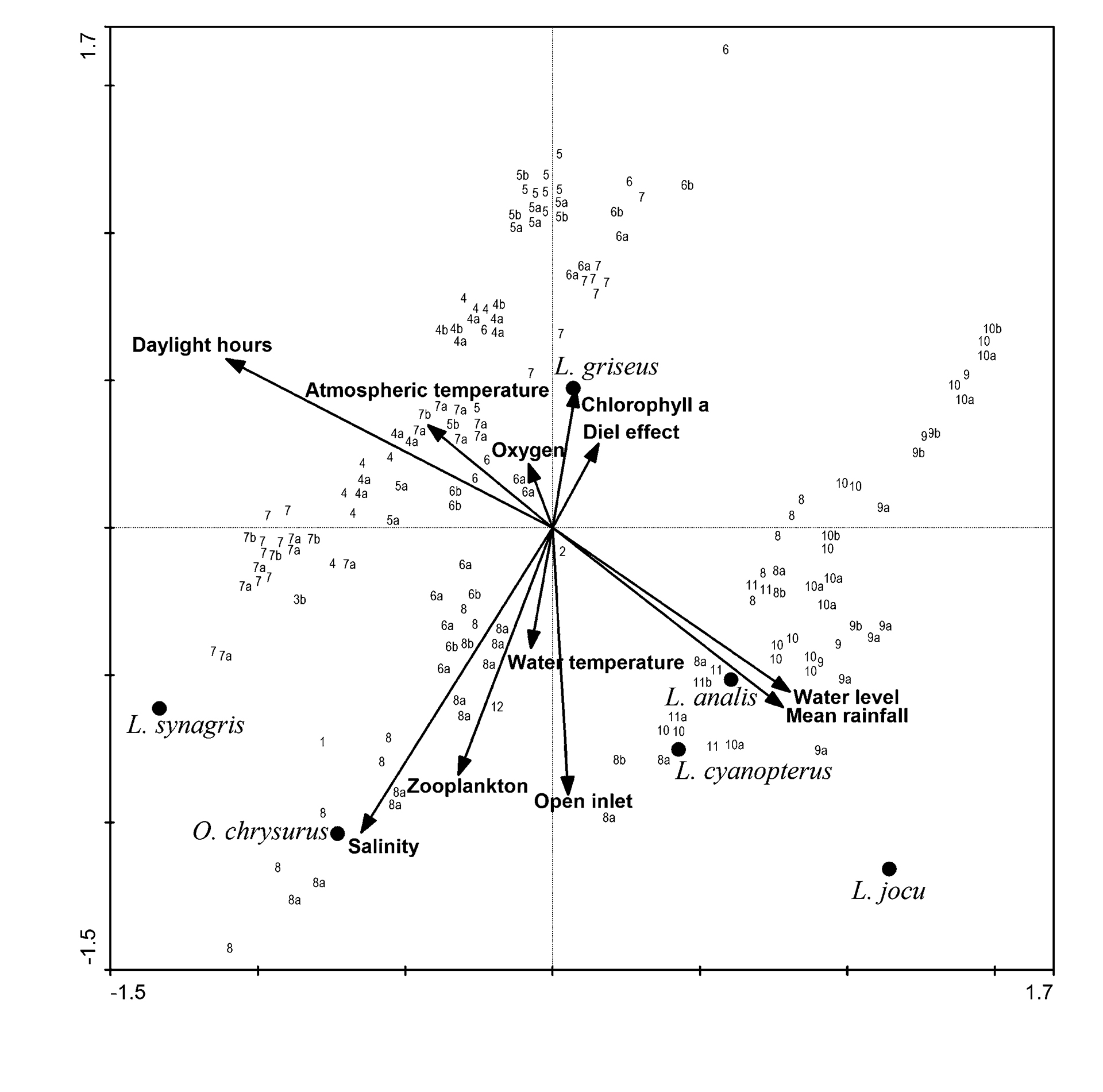
FIGURE 4| Canonical Correspondence Analysis tri-plot of species, samples (months represented by numbers; letters “a” and “b” represent night and twilight samples, respectively, and the absence of letter represents daytime samples) and environmental variables (arrows). Diel periods were considered to be in a light-dark gradient in an ordinal scale with values of 2 (day), 1 (twilight), and 0 (night).
TABLE 4 | Inter-set correlations between environmental factors and species scores, significance of these factors (unrestricted Monte Carlo permutations), and variance inflation factors (VIF), from Canonical Correspondence Analysis. Significant P values are shown in bold.
Inter-set correlations | ||||
Environmental factors | Axis 1 | Axis 2 | P(permuted) | VIF |
Daylight hours | -0.386 | 0.184 | 0.002 | 8.416 |
Salinity | -0.227 | -0.332 | 0.002 | 3.294 |
Mean rainfall | 0.273 | -0.196 | 0.002 | 3.570 |
Zooplankton | -0.111 | -0.270 | 0.078 | 1.327 |
Atmospheric temperature | -0.148 | 0.112 | 0.112 | 7.682 |
Day/twilight/night effect | 0.054 | 0.091 | 0.530 | 1.185 |
Dissolved oxygen | -0.029 | 0.070 | 0.612 | 3.486 |
Open inlet | 0.019 | -0.291 | 0.654 | 2.858 |
Water level | 0.281 | -0.179 | 0.718 | 2.431 |
Chlorophyll a | 0.027 | 0.151 | 0.78 | 1.271 |
Water temperature | -0.026 | -0.132 | 0.828 | 4.734 |
These results reveal a main ordination gradient related to daylight hours, and in the opposite direction to rainfall (Fig. 4). In this sense, the abundance of L. analis, L. jocu and L. cyanopterus was inversely associated with the duration of daylight and directly related to rainfall. High water levels also seem to be positively linked to the abundance of these species. Another major trend, nearly perpendicular to the first, was related to a salinity gradient, in which L. synagris was mainly related to higher salinities, while in the opposite way, L. griseus was mainly associated with relatively lower salinities (Fig. 5).

FIGURE 5 | Number of individuals captured of the two most abundant species, Lutjanus griseus and L. synagris, according to a salinity gradient.
Discussion
Considering that the length at first maturity of these species varies from 225 to 520 mm (García-Cagide et al., 1994), the individuals captured by us were mostly juveniles. Thus, although adults of most snapper species are associated with reefs, juvenile of several species of lutjanids are known to enter brackish waters (Anderson, 2002; Chen, 2017), as observed herein. In this way, six species of snapper were recorded, all confirmed by DNA barcoding (COI). In particular, the capture of O. chrysurus represents one of the few records of this species for estuarine systems in the southwestern Gulf of Mexico (Castillo-Rivera et al., 2020). Thus, La Mancha lagoon is the system with the highest number of snapper species recorded in all estuarine environments of the southern Gulf of Mexico (Lara-Domínguez et al., 2011), mainly due to intensive sampling used by us, both seasonal and diel.
Seasonal migration of snapper species to the lagoon occurred mainly during the summer and early autumn, when the smallest individuals of L. griseus, L. synagris, and L. analis were recorded (July and October). During this period, longer daytime, higher atmospheric temperature, and heavy rainfall characterized the study area. In this sense, CCA results showed that the variables with a higher seasonal incidence (daytime length, salinity, and rainfall) had a significant correlation with the abundance of the lutjanid species.
Seasonal ecological effects caused by photoperiod and temperature are often considered minimal in tropical regions (Castillo-Rivera, 2013; Souza et al., 2018; Whitfield, 2021). However, although the difference in daylight length throughout the year was only 2:18 h (13:18 h in June and 11:00 h in December), this seasonal variation of the photoperiod (i.e., day length) in the study area was the factor with the highest correlation value with the abundance of lutjanid species. The importance of the length of day is that it can act as a stimulus (along with changes in water temperature), for migration, reproduction and feeding. For estuarine fish, photoperiod and temperature have been shown to affect larval growth and survival (Sirois, Dodson, 2000), sexual maturity (Wang et al., 2001; Guèye et al., 2013), migration and residence within the estuary (Capossela et al., 2013; Garcia, Pessanha, 2018). Thus, as in many species, the photoperiod can actas a cue for seasonal migration. In this sense, the migration within the lagoon of L. analis, L. jocu and L. cyanopterus was linked to shorter days, and the migration of. L. griseus and L. synagris was associated with longer periods of daylight.
Regarding salinity, a segregation of the two most abundant species was observed along a gradient, in which L. griseus was more abundant at lower salinities (< 27), while this species was slightly displaced by L. synagris at higher salinities (> 32). Salinity has important direct effects on the distribution of fish in estuarine areas (Barletta et al., 2005; Whitfield, 2021), as well as indirectly through the modification of the result of biotic interactions, such as competition between species (Pessanha, Araújo, 2003; Castillo-Rivera, Kobelkowsky, 2000; Whitfield, 2021). In relation to interspecific competition for resources, it is relatively common for eurytopic species to be weaker competitors, while stenotopic species are stronger competitors. In general, L. griseus has been found to possess similar or higher osmoregulatory capabilities compared to many euryhaline species (especially compared with Lutjanus species), and thus should be considered an euryhaline species (Castro-Aguirre et al., 1999; Serrano et al., 2011). In contrast, in the current study, L. synagris was found more regularly under euhaline conditions. In this context, the observed segregation between these species may also be related to osmoregulatory capabilities and metabolic costs. Thus, in a low salinity range, metabolic costs are significantly lower for L. griseus (Wuenschel et al., 2005), whereas L. synagris will require more energy simply to maintain itself, putting it at a serious competitive disadvantage with L. griseus. In a high salinity range, L. griseus requires more energy for daily maintenance and less is available for other biological functions (Wuenschel et al., 2005), whereas metabolic costs may be lower for L. synagris and its abundance in number may be greater than that of L. griseus.
Rainfall and associated runoff constitute an important environmental drive of fish abundance in tropical estuarine communities (Castillo-Rivera, 2013; Andrade et al., 2015; Castillo-Rivera et al., 2017; Souza et al., 2018). In general, higher rainfall in tropical latitudes increases river discharge and riverine freshwater runoff, and brings an increased amount of allochthonous organic matter and dissolved nutrients to systems, leading to higher primary productivity and food availability. Thus, an increase in food resources would favor juvenile individuals entering the estuarine system for food (Castillo-Rivera et al., 2010; Garcia, Pessanha, 2018).
Although zooplankton biomass and water level did not have a significant effect, these variables tended to have some importance on migration patterns. Zooplankton biomass showed a significant pulse in August and the migration of L. synagris and O. chrysurus may be favored when this resource is abundant. Indeed, planktonic crustaceans are dominant in the diet of juveniles of these species (e.g., Sierra, 1997; Lustosa-Costa et al., 2020). Similarly, flood and high tide, when the inlet is open, can be primarily used by L. analis, L. cyanopterus, and L. jocu to enter the estuarine system.
CCA also revealed negligible importance of environmental variables recorded in situ, such as water temperature, dissolved oxygen, and chlorophyll a, even though these variables showed differences between months and between diel periods. In the same way, the diel variability also seems to be of little importance. Thus, although the less abundant species were recorded exclusively at twilight and night hours, only L. griseus showed differences among diel periods (with grater abundance in the late afternoon and dusk). This may be related to migrations at dusk from their daytime shelter sites to the lagoon at night to feed, as has been observed for lutjanid species in other coastal environments (Nagelkerken et al., 2000; Ley, Halliday, 2007).
Thus, the migration patterns of lutjanid species into La Mancha lagoon were mainly correlated with variables of greater seasonal influence (i.e., day length, salinity and rainfall), present during the warm-rainy period (summer and early autumn). Two of these environmental variables (day length and rainfall) have regional rather than local coverage. In this sense, the variables with regional records had the same or more importance than the variables recorded in situ, so the environmental processes operating at local and regional scales may act synergistically on the distribution patterns at the local scale.
Acknowledgments
This research was funded by the Universidad Autónoma Metropolitana, Unidad Iztapalapa. Luis Guillermo Constante-Pérez had the support of the Consejo Nacional de Ciencia y Tecnología (CONACyT), through the scholarship: CVU–637206. Constructive comments on the manuscript from two anonymous reviewers were very valuable.
References
Alonso Aller E, Gullström M, Eveleens Maarse FKJ, Gren M, Nordlund LM, Jiddawi N, Eklöf JS. Single and joint effects of regional‑ and local‑scale variables on tropical seagrass fish assemblages. Mar Biol. 2014; 161:2395–405. http://doi.org/10.1007/s00227-014-2514-7
Anderson MJ. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001; 26(1):32–46. http://doi.org/10.1111/j.1442-9993.2001.01070.pp.x
Anderson MJ, Gorley RN, Clarke KR. PERMANOVA+ for PRIMER: Guide to software and statistical methods. PRIMER-e: Plymouth, UK; 2008.
Anderson WD. Lutjanidae. In: Carpenter KE, editor. The living marine resources of the Western Central Atlantic. Volume 3: Bony fishes part 2 (Opistognathidae to Molidae), sea turtles and marine mammals. FAO Species Identification Guide for Fishery Purposes and American Society of Ichthyologists and Herpetologists Special Publication; 2002. p.1479–504.
Andrade H, Santos J, Ixquiac MJ. Ecological linkages in a Caribbean estuary bay. Mar Ecol Prog Ser. 2015; 533:29–46. https://doi.org/10.3354/meps11342
Barletta M, Barletta-Bergan A, Saint-Paul U, Hubold G. The role of salinity in structuring in structuring the fish assemblages in tropical estuaries. J Fish Biol. 2005; 66(1):45–72. http://doi.org/10.1111/j.0022-1112.2005.00582.x
ter Braak CJF, Šmilauer P. CANOCO reference manual and CanoDraw for Windows. User’s guide: software for canonical community ordination (version 4.5). Ithaca, New York; 2002.
Bruno DO, Barbini SA, Díaz Astarloa JM, Martos P. Fish abundance and distribution patterns related to environmental factors in a choked temperate coastal lagoon (Argentina). Braz J Oceanogr. 2013; 61(1):43–53. https://doi.org/10.1590/S1679-87592013000100005
Bruno DO, Cousseau MB, Díaz Astarloa JM, Acha EM. Recruitment of juvenile fishes into a small temperate choked lagoon (Argentina) and the influence of environmental factors during the process. Sci Mar. 2015; 79(1):43–55. http://dx.doi.org/10.3989/scimar.04085.02A
Capossela KM, Fabrizio MC, Brill RW. Migratory and within-estuary behaviors of adult Summer Flounder (Paralichthys dentatus) in a lagoon system of the southern mid-Atlantic Bight. Fish B-NOAA. 2013; 111(2):189–201. http://doi.org/10.7755/FB.111.2.6
Castillo-Rivera M. Influence of rainfall pattern in the seasonal variation of fish abundance in a tropical estuary with restricted marine communication. J Water Resource Prot. 2013; 5(3):311–19. http://doi.org/10.4236/jwarp.2013.53A032
Castillo-Rivera M, Kobelkowsky A. Distribution and segregation of two sympatric Brevoortia species (Teleostei: Clupeidae). Estuar Coast Shelf Sci. 2000; 50(1):593–98. https://doi.org/10.1006/ecss.1999.0588
Castillo-Rivera M, Zárate-Hernández R, Ortiz- Burgos S, Zavala-Hurtado J. Diel and seasonal variability in the fish community structure of a mud-bottom estuarine habitat in the Gulf of Mexico. Mar Ecol-Evol Persp. 2010; 31(4):633–42. http://doi.org/10.1111/j.1439-0485.2010.00394.x
Castillo-Rivera M, Ortiz-Burgos S, Zárate-Hernández R. Temporal changes in species richness and fish composition in a submerged vegetation habitat in Veracruz, Mexico. Acta Ichthyol Piscat. 2017; 47(1):23–32. http://doi.org/10.3750/AIEP/01997
Castillo-Rivera M, Constante-Pérez LG, Serrato-Díaz A. First record and DNA barcode for Ocyurus chrysurus (Lutjanidae) from a coastal lagoon in the southwestern Gulf of Mexico. Lat Am J Aquat Res. 2020; 48(3):492–96. http://doi.org/10.3856/vol48-issue3-fulltext-2404
Castro-Aguirre JL, Espinosa-Pérez H, Schmitter-Soto JJ. Ictiofauna estuarino-lagunar y vicaria de México. México: Limusa; 1999.
Chen Y. Fish resources of the Gulf of Mexico. In: Ward CH, editor. Habitats and biota of the Gulf of Mexico: Before the deepwater horizon oil spill, Vol. II. New York: Springer; 2017. p.869–1038. https://doi.org/10.1007/978-1-4939-3456-0_1
Clarke KR, Gorley RN. PRIMER v7: User Manual/Tutorial. Plymouth: PRIMER-E; 2015.
Contreras-Espinosa F, Rivera-Guzmán NE, Segura-Aguilar R. Nutrientes y productividad primaria fitoplanctónica en una laguna costera tropical intermitente (La Mancha, Ver.) del Golfo de México. Hidrobiológica. 2005; 15(3):299–310. Available from: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0188-88972005000300006
Garcia AFS, Pessanha ALM. Temporal dynamics of tidal mudflat fish assemblages in north-eastern Brazil: short and medium-term variations. J Mar Biol Assoc UK. 2018; 98(7):1745–55. https://doi.org/10.1017/S0025315417001199
García-Cagide A, Claro R, Koshelev BV. Reproducción. In: Claro R, editor. Ecología de los peces marinos de Cuba. México: Instituto de Oceanología, Academia de Ciencias de Cuba, Centro de Investigaciones de Quintana Roo (CIQRO); 1994. p.187–262.
Gotelli NJ, Ellison AM. A primer of ecological statistics. Sunderland, Massachusetts: Sinauer Associates; 2004.
Guèye M, Kantoussan J, Tine M. The impact of environmental degradation on reproduction of the black-chinned tilapia Sarotherodon melanotheron from various coastal marine, estuarine and freshwater habitats. CR Biol. 2013; 336(7):342–53. http://dx.doi.org/10.1016/j.crvi.2013.05.004
Hagan SM, Able KW. Diel variation in the pelagic fish assemblage in a temperate estuary. Estuaries Coasts. 2008; 31:33–42. http://dx.doi.org/10.1007/s12237-007-9018-3
Hayduk JL, Hacker SD, Henderson JS, Tomas F. Evidence for regional-scale controls on eelgrass (Zostera marina) and mesograzer community structure in upwelling-influenced estuaries. Limnol Oceanogr. 2019; 64(3):1120–34. http://doi.org/10.1002/lno.11102
Lara-Domínguez AL, Day JW, Yáñez-Arancibia A, Sáinz-Hernández E. A dynamic characterization of water flux through a tropical ephemeral inlet, La Mancha lagoon, Gulf of Mexico. In: Singh VP, Xu YJ, editors. Coastal Hydrology and Processes. Chelsea: Water Resources Publications; 2006. p.413–22.
Lara-Domínguez AL, Franco-López J, Bedia-Sánchez C, Abarca-Arenas LG, Díaz-Ruiz S, Aguirre-León A, González-Gándara C, Castillo-Rivera M. Diversidad de peces en los ambientes costeros y plataforma continental. In: Cruz Aragón C, editor. La Biodiversidad en Veracruz: Estudio de Estado. Xalapa: CONABIO, Gobierno del Estado de Veracruz, Universidad Veracruzana, Instituto de Ecología, AC; 2011. p.505–16.
Lepš J, Šmilauer P. Multivariate analysis of ecological data using CANOCO. New York: Cambridge University Press; 2003.
Ley JA, Halliday IA. Diel variation in mangrove fish abundances and trophic guilds of northeastern Australian estuaries with a proposed trophodynamic model. B Mar Sci. 2007; 80(3):681–720. Available from: https://www.ingentaconnect.com/content/umrsmas/bullmar/2007/00000080/00000003/art00016#
Lustosa-Costa SY, Duarte MRN, Araújo PRV, Pessanha ALM. Resource partitioning among juvenile snappers in a semi-arid estuary in north-eastern Brazil. J Mar Biol Assoc UK. 2020; 100(5):807–16. https://doi.org/10.1017/S0025315420000375
McEachran JD, Fechhelm JD. Fishes of the Gulf of Mexico. Volume 2: Scorpaeniformes to Tetraodontiformes. Austin, Texas: University of Texas Press; 2005.
Mecalco-Hernández Á, Castillo-Rivera MA, Sanvicente-Añorve L, Flores-Coto C, Álvarez-Silva, C. Variación estacional y nictímera en la distribución del zooplancton dominante en una laguna costera tropical. Rev Biol Mar Oceanog. 2018; 53(1):39–49. https://doi.org/10.4067/S0718-19572018000100039
Nagelkerken I, Dorenbosch M, Verberk WCEP, Cocheret de la Moriniere E, van der Velde G. Day-night shifts of fishes between shallow-water biotopes of a Caribbean bay, with emphasis on the nocturnal feeding of Haemulidae and Lutjanidae. Mar Ecol Prog Ser. 2000; 194:55–64. https://doi.org/10.3354/meps194055
Nelson JS, Grande TC, Wilson MVH. Fishes of the world. 5th ed. Hoboken, New Jersey: John Wiley & Sons; 2016.
Pessanha ALM, Araújo FG. Spatial, temporal and diel variations of fish assemblages at two sandy beaches in the Sepetiba Bay, Rio de Janeiro, Brazil. Estuar Coast Shelf Sci. 2003; 57(5–6):817–28. https://doi.org/10.1016/S0272-7714(02)00411-0
Sanvicente-Añorve L, Sánchez-Ramírez M, Ocaña-Luna A, Flores-Coto C, Ordóñez-López U. Metacommunity structure of estuarine fish larvae: the role of regional and local processes. J Plankton Res. 2011; 33(1):179–94. http://doi.org/10.1093/plankt/fbq098
Scientific Committee on Oceanic Research (SCOR). Determination of photosynthetic pigments in sea-water. Monographs on Oceanographic Methodology. France, UNESCO; 1966.
Serrano X, Serafy J, Grosell M. Osmoregulatory capabilities of the gray snapper, Lutjanus griseus: salinity challenges and field observations.Mar Freshw Behav Phy. 2011; 44(3):185–96. http://dx.doi.org/10.1080/10236244.2011.585745
Servicio Meteorológico Nacional (SMN). Normales climatológicas por estado [Internet]. México; 2020. Available from: https://smn.conagua.gob.mx/es/informacion-climatologica-por-estado?estado=ver
Sierra LM. Relaciones tróficas de los juveniles de cinco especies de pargo (Pisces: Lutjanidae) en Cuba. Rev Biol Trop. 1997; 45(1):499–506. https://revistas.ucr.ac.cr/index.php/rbt/article/view/30659
Sirois P, Dodson JJ. Critical periods and growth-dependent survival of larvae of an estuarine fish, the rainbow smelt Osmerus mordax. Mar Ecol Prog Ser. 2000; 203:233–45. https://doi.org/10.3354/meps203233
Souza CD, Batista VS, Fabré NN. What are the main local drivers determining richness and fishery yields in tropical coastal fish assemblages? Zoologia. 2018; 35:e12898. https://doi.org/10.3897/zoologia.35.e12898
Valesini FJ, Tweedley JR, Clarke KR, Potter IC. The importance of regional, system-wide and local spatial scales in structuring temperate estuarine fish communities. Estuaries Coasts. 2014; 37:525–47. http://doi.org/10.1007/s12237-013-9720-2
Victor BC, Hanner R, Shivji M, Hyde J, Caldow C. Identification of the larval and juvenile stages of the Cubera Snapper, Lutjanus cyanopterus, using DNA barcoding. Zootaxa. 2009; 2215:24–36. https://doi.org/10.11646/zootaxa.2215.1.2
Wang HY, Weng CF, Tu MC, Lee SC. Synchronization of plasma sexual steroid concentrations and gonadal cycles in the sleeper, Eleotris acanthopoma. Zool Stud. 2001; 40:14–20. Available from: zoolstud.sinica.edu.tw/Journals/40.1/14.pdf
Whitfield AK. Estuaries – how challenging are these constantly changing aquatic environments for associated fish species? Environ Biol Fish. 2021; 104:517–28. https://doi.org/10.1007/s10641-021-01085-9
Wuenschel MJ, Jugovich AR, Hare JA. Metabolic response of juvenile gray snapper (Lutjanus griseus) to temperature and salinity: Physiological cost of different environments. J Exp Mar Biol Ecol. 2005; 321(2):145–54. https://doi.org/10.1016/j.jembe.2005.01.009
Authors

![]() Luis Guillermo Constante-Pérez1,2,
Luis Guillermo Constante-Pérez1,2, ![]() Manuel Castillo-Rivera1
Manuel Castillo-Rivera1 ![]() and
and ![]() Alejandra Serrato-Díaz3
Alejandra Serrato-Díaz3
[1] Laboratorio de Peces, Departamento de Biología, Universidad Autónoma Metropolitana, Unidad Iztapalapa, Avenida San Rafael Atlixco, 186, Col. Vicentina, Iztapalapa, 09340 Ciudad de México, Mexico. (LGCP) consper_luisg@xanum.uam.mx, (MCR) crma@xanum.uam.mx (corresponding author).
[2] PhD Program, Doctorado en Ciencias Biológicas y de la Salud. Universidad Autónoma Metropolitana, Avenida San Rafael Atlixco, 186, Col. Vicentina, Iztapalapa, 09340 Ciudad de México, Mexico.
[3] Laboratorio Divisional de Biología Molecular, Universidad Autónoma Metropolitana, Unidad Iztapalapa, Avenida San RafaelAtlixco, 186, Col. Vicentina, Iztapalapa, 09340 Ciudad de México, Mexico. alej@xanum.uam.mx.
Authors’ Contribution 

Luis Guillermo Constante-Pérez: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Writing-original draft, Writing-review and editing.
Manuel Castillo-Rivera: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Writing-original draft, Writing-review and editing.
Alejandra Serrato-Díaz: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Supervision, Validation, Writing-original draft, Writing-review and editing.
Ethical Statement
All biological collections were authorized and approved by the Cooperativa Pesquera La Mancha (fishermen’s cooperative).
Competing Interests
The authors declare no competing interests.
How to cite this article
Constante-Pérez LG, Castillo-Rivera M, Serrato-Díaz A. Diversity, seasonal and diel distribution of snappers (Lutjanidae: Perciformes) in a tropical coastal inlet in the southwestern Gulf of Mexico. Neotrop Ichthyol. 2022; 20(2):e220009. https://doi.org/10.1590/1982-0224-2022-0009
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2022 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted May 19, 2022 by Fernando Gibran
Accepted May 19, 2022 by Fernando Gibran
![]() Submitted July 24, 2021
Submitted July 24, 2021
![]() Epub June 24, 2022
Epub June 24, 2022