![]() Jonas de Andrade Santos1
Jonas de Andrade Santos1 ![]() ,
, ![]() Rafael Menezes1,
Rafael Menezes1, ![]() Ricardo de Souza Rosa2 and
Ricardo de Souza Rosa2 and ![]() Telton Pedro Anselmo Ramos2
Telton Pedro Anselmo Ramos2
PDF: EN XML: EN | Cite this article
Abstract
Sciaenidae species (croakers and drums) are known for specializations in their sensory apparatus, such as otoliths and lateral line. Within this family, some Stelliferinae members remain taxonomically unresolved due to their cryptic aspects and low phenotypic disparities. Stellifer punctatissimus comprises a species complex putatively formed by three species on morphological grounds, but molecular data have indicated only two evolutionary lineages. Herein, we tested the hypothesis that this complex is composed of Stellifer gomezi (GM), S. menezesi (MN), and S. punctatissimus (PC), using the otolith shape and morphology to differentiate them. Seventy-seven sagittal otoliths (GM = 30, MN = 30, PC = 17) were photographed to outline the otolith contour by Elliptical Fourier descriptors. Ten otoliths for each group were used to detect shape variation in the sulcus acusticus through semilandmarks. Differences in otolith shape contour were recorded among the species by PERMANOVA (Pseudo-F = 4.02, df = 2, p < 0.001) and corroborated by the Linear Discriminant Analysis, which demonstrated three partially segregated groups with satisfactory re-classification rates. A larger sulcus acusticus (GM and MN), rounded projection (PC), and distinct rectangularity pattern were also recorded. Our results support the morphological hypothesis and thus contribute to narrowing the taxonomic gaps in Sciaenidae.
Keywords: Croakers, Cryptic species, Otolith morphology, Speciation, Taxonomy.
As espécies de Sciaenidae (corvinas e pescadas) são conhecidas por especializações nos aparatos sensoriais, como otólitos e linha lateral. Nesta família, alguns membros de Stelliferinae permanecem com sua taxonomia não resolvida devido às características crípticas e baixas disparidades fenotípicas. Stellifer punctatissimus supostamente compreende um complexo formado por três espécies, mas os dados moleculares têm indicado apenas duas linhagens evolutivas. Aqui, testamos a hipótese de que esse complexo é composto por Stellifer gomezi (GM), S. menezesi (MN), e S. punctatissimus (PC), usando a forma e morfologia do otólito para diferenciá-las. Setenta e sete otólitos sagita (GM = 30, MN = 30, PC = 17) foram fotografados para delimitar o seu contorno pelos descritores Elípticos de Fourier. Dez otólitos, por grupo, foram usados para detectar a variação de forma no sulcus acusticus através dos semilandmarks. Foram registradas diferenças na forma do otólito entre as espécies através da PERMANOVA (Pseudo-F = 4,02, df = 2, p < 0,001) e pela análise discriminante linear, demonstrando três grupos parcialmente segregados com taxas de reclassificação satisfatórias. Também foram registrados um sulcus acusticus mais largo (GM e MN), projeção arredondada (PC) e padrões alométricos distintos no índice de retangularidade. Nossos resultados apoiam a hipótese morfológica; assim, contribuem na redução das lacunas taxonômicas em Sciaenidae.
Palavras-chave: Especiação, Espécies crípticas, Morfologia do otólito, Pescadinhas, Taxonomia.
Introduction
Sciaenidae is one of the most speciose families within Acanthuriformes (nearly 300 species), and members of this family are widely known as ‘croakers’ or ‘drums’ (i.e., they produce sound) (Chao et al., 2015; Fricke et al., 2022). The diversity of morphological traits in the family has enabled the adaptation of the species to a variety of habitats, such as estuaries, sand bottoms, and surf zones, with few species being strictly freshwater or reef-associated (Chao et al., 2015). Although recent evidence based on morphological and molecular approaches has supported the monophyly of this family, evolutionary relationships in less inclusive taxonomic ranks remain uncertain (Chao, 1978; Lo et al., 2015). As an example, the status of some genera of the monophyletic subfamily Stelliferinae (also known as ‘Stellifer-group’ sensu Chao, 1978) is still unsolved due to their great morphological similarity or even the lack of diagnostic characters to distinguish them (Chao et al., 2015; Silva et al., 2018).
The paucity of informative characters has led to shifts in the generic positioning of morphologically similar species, such some originally described in Stellifer Oken, 1817 and later changed to Ophioscion Gill, 1863, or vice versa (Chao et al., 2021). Species of both genera are common or abundant, particularly in coastal waters of North-Northeastern Brazil, frequently caught as bycatch in shrimp trawls. Despite their abundance, the taxonomic status of several species remains undefined (Chao et al., 2015; Silva et al., 2018). A particular case of uncertainty is Stellifer punctatissimus (Meek & Hildebrand, 1925), initially described in Ophioscion by Meek, Hildebrand (1925). Later, a species complex with two additional species was recognized by Chao (2002) using a morphological approach. Recent molecular data, however, have pointed out only two evolutionary lineages within this complex (Barbosa et al., 2014; Silva et al., 2018). Finally, one of these species was described and another redescribed, both now included in Stellifer, which is considered a senior synonym of Ophioscion (Chao et al., 2021).
Otoliths are calcified structures located in the inner ear of bony fishes (Teleostei) disposed in the three semicircular canals (utricle, saccule and lagena), which correspond to the lapillus, sagitta and asteriscus pairs (Schulz-Mirbach et al., 2019). They are surrounded by endolymph and connected to the macula (which holds the sensory hair cells and nerves) by the otolithic membrane, which lies on the sulcus acusticus (Popper et al., 2005; Schulz-Mirbach et al., 2019). Due to their higher density when compared to the fish body, otoliths exhibit a different response (i.e., distinct amplitudes and phases) to sound- or motion-induced movements (e.g., angular acceleration). Thus, variations in otolith shape, as well as differences in the features of their sensory cells (e.g., stereocilia number, cell orientation), may be correlated with directional sound sensitivity (Ramcharitar et al., 2001; Popper et al., 2005). Furthermore, otolith shape is influenced by extrinsic factors, such as salinity and temperature, and intrinsic, such as physiology and ontogeny (Campana, Thorrold, 2001; Clark et al., 2021).
By growing throughout the fish life cycle, otoliths also seem to respond to other factors, such as ontogenetic shifts and those linked to environmental gradients (Schulz-Mirbach et al., 2019). Likewise, as they carry a signal of those traits, surveys that use otolith shape provide an indirect and effective way to investigate the living habits of species and to seek diagnostic details in cryptic species. Such analyses contrast with traditional taxonomic approaches, which generally show more overlap than otolith shape analyses (Lombarte et al., 2006; Capoccioni et al., 2011). Many techniques have been used to delineate the otolith shape contour (e.g., shape indices, wavelet, Fourier descriptors). Shape indices appear to be powerless to differentiate cryptic species due to their low resolution. However, they provide valuable biological information by shedding light on the association of shape patterns with habits such as feeding and water depth (Wong et al., 2016; Assis et al., 2020). Fourier descriptors are definitely more efficient in distinguishing slight differences in cryptic- and similar-species than other methods, particularly because they decompose the otolith’s contour into several harmonics, which encompass almost the entire variation across that border (Wong et al., 2016; Avigliano et al., 2018). In contrast, the application of geometric morphometric methods (GMM) is impaired by methodological constraints. In body shape, this method likely achieves a fine resolution of cryptic species or species-rich groups (Anjos et al., 2020; Argolo et al., 2020). However, due to the lack of homologous points, GMM’s use in otolith analysis is mostly restricted to contour descriptions with semi-landmarks (Tuset et al., 2016). However, it provides additional data when applied to structures such as the sulcus acusticus (i.e., ostium and cauda); as a result, such information could be applied to discuss hearing capabilities, taxonomy, and prey identification (Byrd et al., 2020; Granados-Amores et al., 2020).
This study aimed to test the following hypothesis: Stellifer punctatissimus species complex is formed by three distinct cryptic species that can be distinguished on morphological grounds by complementary tools such as otolith shape analysis.
Material and methods
Sampling sites and otolith preparation. The fishes were sampled along the coast of Bahia state, and 77 right sagittal otoliths of three Stellifer species were sampled as follows: Stellifer gomezi (Cervigón, 2011) (GM) – 30; Stellifer menezesi Chao, Carvalho-Filho & Andrade Santos, 2021 (MN) – 30; and Stellifer punctatissimus (Meek & Hildebrand, 1925) (PC) – 17. The sampling was carried out at four sites: Itaparica Island, Porto Seguro (#2), and Caravelas (Tab. 1; Fig. 1). The surveys were conducted in April 2018 and February 2019 on Itaparica Island, May and March 2016 in Porto Seguro and Caravelas, respectively. The vouchers were deposited at the fish collection of Museu de Zoologia da Universidade Estadual de Feira de Santana, Feira de Santana, Bahia, Brazil (MZFS) under the numbers 17694, 18115, 18116, 18117, 18119, 18124, 18125, 18126, 18127, 18128, 18129, 18131, 18133, 18135. Five trawls were deployed parallel to the coast (totaling about 250 m) using a manual beach-seine net (9 m in length, 1.7 m in height, 13 mm mesh on the lateral, and 5 mm mesh in the center). At the Caravelas site, fish sampling was performed in shrimp trawling, and specimens were caught as bycatch.
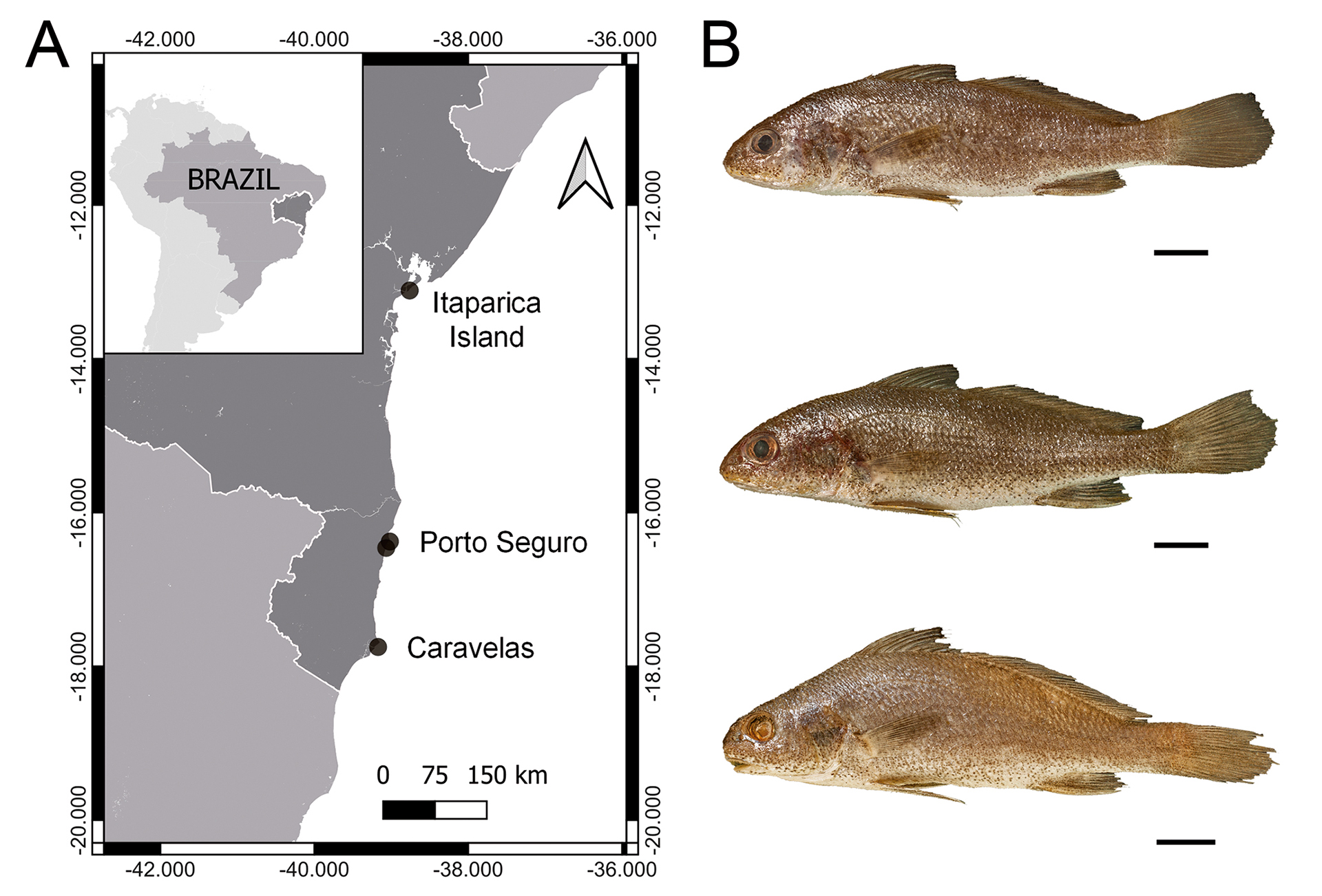
FIGURE 1| Map of the study area highlighting the sampling sites in the Bahia State, Brazil. B. Representative specimens of the species studied. Stellifer gomezi (top); S. menezesi (middle); S. punctatissimus (bottom). Scale bars = 10 mm.
TABLE 1 | The number of specimens analyzed per site, their mean body size (in mm) with standard deviation (SD) and length range by species.
Species | Sites | Body length | |||
Itaparica | Porto
Seguro | Caravelas | Mean ± SD | Range | |
Stellifer gomezi | 21 | 9 | – | 67.24±14.31 | 46.43-111 mm |
Stellifer menezesi | 15 | 15 | – | 65.50±15.05 | 38.57-99.98 mm |
Stellifer punctatissimus | 4 | 12 | 1 | 60.45±20.32 | 35.33-109 mm |
Sagittal otoliths were extracted preferentially by cutting through the upper end of the right gill cover. Otoliths were then manually cleaned, washed in distilled water, dried, and stored in individually labeled Eppendorf®️ tubes. Right otoliths were photographed using a stereomicroscope Leica EZ4 HD (for contour analysis) and Leica M205 A (for description and geometric morphometrics). All pictures had their contrast improved in Adobe Photoshop CC 2019 and were digitally cleaned to avoid noise in contour analysis. Otolith’s descriptive nomenclature (Fig. 2) was based on Chao (1978), Schwarzhans (1993), and Aguilera et al. (2016).

FIGURE 2| Illustration of representative sagitta otolith with key anatomical features, based on an average shape specimen. Filled dots = landmarks, empty circles = semilandmarks; A = anterior, M = medial, D = dorsal (asterisk: sulcus acusticus = ostium + cauda).
Statistical analyses. Each image was processed by ImageJ software (Schneider et al., 2012) to obtain its pixel-cm ratio. Initially, the following shape indices were calculated: aspect ratio, circularity, ellipticity, form factor, rectangularity, and roundness. An ANOVA test was used to determine differences among species using species as a fixed factor (3 levels: GM, MN, and PC).
We created an analytical framework for assessing the degree of differentiation of the three putative species of the Stellifer punctatissimus complex, inferred from the otolith shape (adapted from Tuset et al., 2020). Firstly, Elliptical Fourier descriptors (EFDs) were used to quantify the otolith shape contour with ‘shapeR’ package (Libungan, Pálsson, 2015) in the R platform (R Development Core Team, 2020). Allometry was examined and controlled by the standard length (SL, in mm) of each fish using stdCoefs function (‘ShapeR’ package). Six EFDs exhibited significant allometric effects and were then omitted (45 – 6 = 39 EFDs). Next, a covariance-based Principal Component Analysis was applied with 39 EFDs for extracting the most informative principal components (as retained for eigenvalues > 1) and thus creating an orthogonal matrix (uncorrelated variables) that avoids multicollinearity (Jolliffe, 1986). This procedure shrunk 39 EFDs into 16 components that explained 94.6% of the total variation of data. Lastly, the normality for each component was examined by Shapiro-Wilk test, with three of them excluded (p < 0.001) (16 – 3 = 13 components), and a Bartlett’s test demonstrated homogeneity of covariance matrix across species (Bartlett’s K-squared = 0.44, df = 2, p = 0.80).
A Linear Discriminant Analysis (LDA) was performed to identify the degree of separation of the species (response variable) based on 13 components of otolith shape contour (predictors). LDA is an adequate technique as it reduces the dimensionality of the data and works also as a classification method. The predictive accuracy of the model was estimated by the percentage of correctly individuals ascribed to their species, using the leave-one-out re-classification method (Venables, Ripley, 2002). Complementarily, a PERMANOVA was performed for testing multivariate differences in the otolith shape contour (13 components) among species, with permutational pairwise comparisons (nperm = 999) used as a post-hoc test (‘RVAideMemoire’ package; Hervé, 2022).
For the geometric morphometric methods (GMM), ten photographs of each species were analyzed using the Tps series (Rohlf, 2017a, 2021). To account for the variation within the sulcus acusticus, a curve was drawn on its entire contour, with landmarks on the initial and final points of that curve. The original curve was resampled to 35 points (by length), which were later assigned as landmarks (LM), by using the function “append tps curves to landmarks”. Semilandmarks (sLM) at the initial and final points of the curve were removed from the dataset, remaining only the 33 points (sLM) from the curve plus the two landmarks (LM) (Fig. 2). In the sequence, the sliding step was performed on those curves based on the minimum bending energy method. All data were initially subjected to a Generalized Procrustes Analysis (GPA) using tpsRelw (Rohlf, 2017b). To describe sulcus acusticus morphology, a Principal Component Analysis (PCA) was performed using the Procrustes residuals. As the dataset had much more variables (33 sLM + 2 LM) than specimens (10 by species), the statistical power was impaired. Thus, in all GMM analyses, we rather consider it as a descriptive approach for such analysis than its statistical significance in regard to distinction among groups. Although the otoliths present an angled inner face in the anteroposterior axis, we disregard such variation in our results once it was viewed by the same tangent space approximation. Thus, by comparing species with similar angled sagittal otoliths, the rules of Kendall’s shape space were not broken (Klingenberg, 2020). It would be a case in a comparison with “flat-otolith” species (e.g., Macrodon spp.) because in this scenario, for instance, the proportions between ostium and cauda would present a bigger disparity.
Results
General otoliths description. Sagittal otoliths’ contour somewhat rectangular. Inner face strongly convex. A well-marked dorsal furrow. Postdorsal spine broad, relatively short. Dorsal depression not well-defined, smooth, and shallow; a curve follows between predorsal spine and predorsal angle. The ontogenetic variation shows a more rounded otolith in juveniles (vs. more rectangular in adults), a smoother ostium in juveniles, and postdorsal spine shorter in juveniles (Fig. 3).

FIGURE 3| Otoliths of Stellifer punctatissimus complex. A. S. gomezi. B. S. menezesi. C. S. punctatissimus. Left: inner face; Middle: inset of the ostium; Right: dorsal profile. Arrow indicates spout-like groove; asterisk indicates projection on the outer face. Scale bars = 1 mm.
Stellifer gomezi: Predorsal spine relatively long, straight to slightly curved outwards in the dorsal profile. Dorsal margin smooth, slightly concave; forming an angle of approximately 130° with predorsal angle. Predorsal angle rounded, occasionally projecting posteriorly. Postventral notch smooth, with an angle of approximately 130°. Ostium short, shallow, and wide; narrowed close to the ostial-caudal joint. A spout-like groove between the ostium and dorsal depression, bent at approximately 100°. Cauda deep, widened, strongly bent; rounded on its tip. Outer face rather straight, except for the posterior margin, slightly convex. A well-defined mid-dorsal projection, rounded to slightly sharp and oriented backward. The ontogenetic variation shows the cauda tip slightly pointed in juveniles, and the projection on the outer face less pronounced in juveniles (Fig. 3A).
Stellifer menezesi: Predorsal spine relatively long, straight, also nearly straight in dorsal profile. Dorsal margin smooth, slightly concave; forming an angle of approximately 140° with predorsal angle. Predorsal angle rounded, occasionally projecting outwards. Postventral notch smooth, with an angle of approximately 130°. Ostium short, generally shallow (except for a depression on its mid-posterior section), slightly ventrally widened, narrowed close to the ostial-caudal joint. A spout-like groove between the ostium and dorsal depression, bent at approximately 130°. Deep, widened, strongly bent cauda, slightly forward-pointed on its tip. Outer face rather straight, except for the posterior margin slightly convex. A well-defined projection on the mid-dorsal section; its margin squarish to rhomboidal and backward oriented. The ontogenetic variation shows the cauda slightly less bent in adults and its tip somewhat pointed in juveniles, predorsal angle less pronounced in juveniles, and the projection on the outer face less pronounced in juveniles (Fig. 3B).
Stellifer punctatissimus: Predorsal spine relatively long, straight, sharp to slightly rounded, nearly outwards directed in dorsal profile. Dorsal margin rather smooth or slightly sinuate forming an angle of approximately 135° with predorsal angle. Predorsal angle rounded, occasionally projecting outwards. Postventral notch smooth, with an angle of approximately 135°. Ostium short, surface irregular (a depression on its mid-posterior section), slightly widened ventrally, narrowed close to the ostial-caudal joint. A shallow, rather inconspicuous spout-like groove between the ostium and dorsal depression, bent at approximately 130°. Cauda deep, widened, strongly bent, slightly forward-pointed on its tip. Outer face flat to somewhat irregular, posterior margin slightly convex. A well-defined projection on the mid-dorsal section; its margin C-shaped to somewhat backward oriented. The ontogenetic variation shows the cauda tip somewhat pointed in juveniles, predorsal spine more rounded in adults, predorsal angle less pronounced in juveniles, projection on the outer face with its margin squarish to rhomboidal and backward oriented in adults (Fig. 3C).
Shape indices and shape analyses. Only the circularity and form factor exhibited statistically significant differences among species (ANOVA: F = 3.3, p < 0.05 and F = 3.1, p < 0.05, respectively), which displayed differences in allometry (Tab. 2; Fig. 4). Differences in otolith allometry were found among the species: negative in Stellifer gomezi and positive in S. menezesi and S. punctatissimus, the latter with the highest rectangularity value among all species (Fig. 4).
TABLE 2 | Values of shape indices by species in the Stellifer punctatissimus complex. Different letters show significant differences (p < 0.05), using ANOVA and Tukey’s post hoc tests.
Species | Shape indices | |||||
Aspect ratio | Circularity | Ellipticity | Form factor | Rectangularity | Roundness | |
Stellifer gomezi | 0.64±0.02 | 16.17±0.46a | 6.44±0.35 | 0.78±0.02a | 4.93±0.44 | 5.19±0.21 |
Stellifer menezesi | 0.64±0.02 | 16.00±0.28 | 6.43±0.33 | 0.78±0.01 | 4.92±0.45 | 5.18±0.17 |
Stellifer punctatissimus | 0.64±0.01 | 15.90±0.28b | 6.37±0.32 | 0.79±0.01b | 4.90±0.47 | 5.25±0.19 |

FIGURE 4| A. Scatterplot of otolith weight using a LOESS curve fitting. B. Scatterplot of rectangularity index. GM = Stellifer gomezi, MN = S. menezesi, PC = S. punctatissimus.
The shape contour (Fourier) of all otolith sections showed a higher variation at the anterior margin: S. gomezi exhibited the most concave ostium contour, while S. menezesi had a more elongated predorsal spine and a rounded predorsal angle (Fig. 5). Results from LDA showed a clustering pattern across species, with a certain degree of overlap near zero on both axes. There was a slight separation between S. punctatissimus (PC) and other species S. gomezi (GM) and S. menezesi (MN): the former had most of its variation enclosed in the positive values of the linear dimension 2, whereas its relatives were almost evenly distributed in positive and negative values. This pattern was repeated in the linear dimension 1, which gave a minor separation between GM and MN, respectively in the negative and positive values. This is reinforced by reclassification accuracy rates attained by the leave-one-out method, with relatively high percentages for GM (73.3%) and MN (80.0%) and a moderate one for PC (52.9%). The observed pattern was again corroborated by PERMANOVA, which recorded multivariate differences in the otolith shape contour among species (Pseudo-F = 4.02, df = 2, p < 0.001), with significant differences for all pairwise combinations: GM and MN (p = 0.001), MN and PC (p = 0.002), and GM and PC (p = 0.046) (Fig. 6).

FIGURE 5| Otolith’s contour reconstruction (Fourier). GM = Stellifer gomezi, MN = S. menezesi, PC = S. punctatissimus.

FIGURE 6| Linear Discriminant Analysis (LDA) of contour shape according to elliptical Fourier descriptors (EFDs). GM = Stellifer gomezi, MN = S. menezesi, PC = S. punctatissimus.
Regarding geometric morphometrics, the PCA explained the major variation of shape in the otolith’s cauda. The first two components of the PCA accounted for 65.1% of the shape variation in the sulcus acusticus; PC1 corresponded to a narrower/larger cauda, angle at the cauda almost straight, slightly bent cauda tip, and a narrower ostium (on the dorso-ventral axis). The second component (PC2), in turn, showed a narrower ostial-caudal joint, a wider ostium (on the antero-posterior axis), and an expansion on the cauda. On the PCA plot, most of the morphospace overlapped among the species. However, slight differences were found among them, especially for Stellifer gomezi (GM) and S. menezesi (MN). Stellifer menezesi (MN) had an almost centered distribution. The GM-MN group was allocated mostly in the positive values of the PC2, while PC exhibited the highest negative values and a more spread distribution. On the pair comparisons, GM-PC displayed the higher variation – linked to the expansion of the cauda, sulcus, and ostium and a less straight caudal curve; followed by MN-PC, with slight expansion on the ostium and caudal tip; finally, the pair GM-MN presented a lower distinction, which was restricted to small variations in caudal curve and ostium expansion (Fig. 7).

FIGURE 7| A. Principal Component Analysis (PCA) of sulcus acusticus’s shape variation using Procrustes residual of geometric morphometric method. Shape variation in each principal component (PC1 in the horizontal and PC2 in the vertical) in the upper left corner. B. Pairwise shape comparisons; GM = Stellifer gomezi, MN = S. menezesi, PC = S. punctatissimus.
Discussion
Integrative approaches have increasingly been used to distinguish cryptic species of coastal fishes (e.g., Carvalho et al., 2020b; Figueiredo-Filho et al., 2021). An initial morphological study indicated the existence of a species complex in Stellifer punctatissimus (Chao, 2002), but this hypothesis has not been confirmed by molecular evidence, which suggested instead a recent speciation with only two evolutionary lineages in this complex (Silva et al., 2018). More recently, all these species were formally recognized through a morphological basis (Chao et al., 2021), but complementary studies were still needed to elucidate their status. Herein, elliptic Fourier descriptors (EFDs) provided further evidence to support our initial hypothesis of a complex formed by three species, especially by satisfactory reclassification rates for all species.
Surveys using single species at spatiotemporal or stock levels have shown higher reclassification rates than those found here (Hoff et al., 2020; Kikuchi et al., 2021). Despite seeming to be rare, rates higher than those found here (52–80%) were already recorded in three (> 90%; Bani et al., 2013) and four (> 95%; Zischke et al., 2016) sympatric species comparisons. However, closely related species comparisons provide a reclassification range similar to ours (i.e., 52–80%; La Mesa et al., 2020). Therefore, these results demonstrate the usefulness of this method to distinguish closely related species. A recent and likely incipient speciation may explain why, despite being a high-resolution method, our LDA showed a partially overlapping among those species. In fact, a previous analysis showed highly segregated groups when the allometry effect was not removed from our data. It might suggest a likely trend in this species complex’ differentiation. Stellifer punctatissimus species complex’ evolutionary history has led to high morphological similarity without clear genetic divergence, where allopatric and sympatric events probably drove the species’ divergence (Silva et al., 2018). It has been shown that some species might differentiate even though they present non-significant genetic variation (Carvalho-Filho et al., 2009; Marceniuk et al., 2019).
However, as elliptic Fourier descriptors (EFDs) is a method for outlining a contour, its automated use is unlikely in “inner” structures such as the sulcus acusticus (i.e., by being embedded inside another contour), which can be used as a proxy for prey identification (e.g., in dolphins’ diet studies – Rodrigues et al., 2020; Byrd et al., 2020). For instance, the geometric morphometric method (GMM) provided a contrasting result, a partially less overlapped pattern among the species. Interestingly, the observed pattern exhibits a “gradient” from Stellifer punctatissimus (PC) to S. menezesi and S. gomezi (MN, GM), which is in agreement with their external similarity. A convergence between body morphology and otolith shape may explain this pattern; the higher similarity between GM-MN than with PC (most visually distinctive species – Fig. 1B) resulted in its final “gradient” morphospace (Chao et al., 2021; unpublished data). In a similar case, La Mesa et al. (2020) found that a relatively morphologically distinct icefish species was also the most differentiated from the other two species by otolith shape. Indeed, previous studies have shown that similar species (i.e., external morphology) may occupy a similar place in the morphospace, either when otolith shape or body shape is analyzed, as seen in the genus Centropomus (Granados-Amores et al., 2020; Figueiredo-Filho et al., 2021). Although the actual shape may be affected by the phylogenetic inertia – similar shape among closely related species coming from a lagged rate of evolution of a trait – it could present distinct growth rates that might allow distinguishing such species (Lombarte et al., 2010).
Studies on species delimitation must consider a broad source of data to yield a reliable taxonomic recommendation, because the evolutionary origin itself might lead to constraints in phenotypic disparity, such as stasis and niche conservatism, it turns more difficult to identify cryptic species than “regular” ones, thus, finer-scale methods are needed (Riddle et al., 2011; Struck, Cerca, 2019). That is, to reject the null hypothesis in species delimitation (i.e., two species in the complex – molecular hypothesis), a much higher amount of data should be used in a well-studied species than to describe/record a new species – in doing so, a better understanding of the evolutionary process would be achieved and taxonomic issues would be solved (Carstens et al., 2013; Hillis et al., 2021). For instance, a recent study has shown that the Bigtooth corvina (Isopisthus parvipinnis)in SW Atlantic, has well-marked groups based on otolith shape (e.g., populations, years), which probably reflects fishery pressure and changes in population structure toward a metapopulation structure (Hoff et al., 2020). Although there might have additional influences like unknown diversity (Guimarães-Costa et al., 2020). In such situations, only the combined use of approaches and data review might lead to a resolution; as discussed herein, different growth rates and otolith shapes aid in a well-based result that places certainty on taxonomic status.
Despite our data came from a single source (otoliths), they were not appraised through a single perspective. For instance, its description reveals some agreement with the current phylogeny, namely, a closer relationship between the Stellifer punctatissimus species complex and S. scierus/S. strabo (both from the Pacific Ocean). That is, they shared a well-marked spout-like groove, with almost a right angle, and shared with fossil species a backward oriented projection on its otolith’s outer face (Aguilera et al., 2016; Silva et al., 2018). In Sciaenidae, a family with an evolutionary perspective of specialization on the sensory apparatus, those characters could help solve incongruencies throughout their phylogeny by being explored in an ancestral state reconstruction (Trewavas, 1977; Schwarzhans, 1993). Additionally, a few differences, such as sulcus acusticus shape and contour angles, add evidence to refute our null hypothesis. Therefore, it agrees with the other methods’ results that indicate the occurrence of three species on its complex.
Conversely, variations in otolith shape of the Stellifer punctatissimus complex can also result from factors not measured herein, such as the metabolic and growth rates (Alewijnse et al., 2021; Jónsson et al., 2021). As Geller (1999) argues, these aspects could help to evaluate cryptic species, as they reveal possible confounding effects in species complexes. This seems to be achieved in our results, which show each species with a specific rate of otolith weight (Fig. 4B), which could suggest a distinct size at first maturity for each species, and in agreement with our initial hypothesis. Additionally, by showing a distinct allometric pattern in the rectangularity index, at least to one of the species (GM), our results further support the hypothesis of three distinct species in the Stellifer punctatissimus species complex. It has been proven that either accretion rates or growth rates in otolith shape can be used as a proxy for differentiating species or even stocks, due to the assumed distinct intrinsic factors by species (Pavlov, 2016; Kikuchi et al., 2021). In the same way, shape indices can be linked to traits such as depth preference, habitat use, and feeding habits (Assis et al., 2020). In doing so, we could apply the differences in ecological niches into a taxonomic approach in order to achieve a better resolution of taxonomic issues, as discussed here. Our data show values above 64% of the aspect-ratio, which may classify all these species as associated with demersal habit over unconsolidated substrata, but do not suggest strictly benthic habit, such as that occurring in Menticirrhus (Jaramillo et al., 2014; Carvalho et al., 2020a). These findings are consistent with data on this species complex, which have changed an incorrect assignment from zoobenthivorous to zooplanktivorous guild (Santos et al., 2021).
Furthermore, of all shape indices, only circularity and form factor were statistically significant. This low contribution to differentiation among the species is not unexpected, because these indices present an unsuitable resolution to detect small differences at the species level (Pavlov, 2016; Wong et al., 2016; Avigliano et al., 2018). Although shape indices rarely support species-delimitation studies, otolith-based approach in Macrodon (Cardoso et al., 2012) had validated previous molecular and morphological hypotheses (Santos et al., 2006; Carvalho-Filho et al., 2010). As discussed by Tuset et al. (2021), the shape indices only provide good results if clearly distinct species are compared – which is not the case for cryptic species. We suggest therefore that Ellipitical Fourier descriptors perform better than shape indices to distinguish cryptic and congeneric species. On the other hand, once the application of homologous landmarks in otoliths is unlikely, we needed to apply several semilandmarks on the sulcus acusticus, as a result, we lost statistical power. Thus, our GMM results would be improved by adding specimens, which could give a balance between variables (semilandmarks) and specimens (otoliths) (Gunz, Mitteroecker, 2013).
As the otolith shape is known to be species-specific, it accurately responds to environmental gradients; thus, we could indicate habitat use through otoliths. We cannot disregard the likelihood of those otolith distinctions holding a signal of a putatively different coastal zone occupation; this could retrieve a recent speciation process likely by ecological divergence (Carvalho-Filho et al., 2009; Silva et al., 2018; Caires et al., 2019). As these shapes vary (e.g., contour, sulcus acusticus) among species, they can be applied for taxonomic purposes to fulfill the gaps within Sciaenidae and in ecological surveys, once otoliths are one of the most informative remains to identify bony fishes in diet studies (Rodrigues et al., 2020; Byrd et al., 2020).
In regard to conservation, both of them – prey and predators – are caught as bycatch in shrimp fisheries, with the former (S. punctatissimus) being classified as Least concern (LC) and Data deficient (DD) according to IUCN and MMA/IBAMA, respectively (ICMBio, 2018; Aguilera Socorro, 2020). To date, S. menezesi has not been evaluated either by IUCN or ICMBIO, and S. gomezi was classified as LC by the last available IUCN’s assessment (Chao, 2020). Despite their lack of commercial value, we could not dismiss the ecological importance of these species because they need better monitoring and some effects of fisheries pressure on marine food webs remain unclear (Márquez-Velásquez et al., 2021; Lira et al., 2022). Due to the possibility of differences in habitat use among those species, such as discussed for the Tonkin weakfish (Caires et al., 2019), the existence of distinct conservation statuses by species in a future assessment is presumable due to the likely specific threats upon each species. For this reason, it is essential to continue pursuing additional data that could elucidate if those species truly have a degree of distinct habitat use throughout their life cycle, which would also help to solve uncertainties in their recent speciation process. Therefore, our results provide significant data that might aid to species’ conservation status assessment, as taxonomic uncertainties are one of the main issues that usually hamper these evaluations.
Acknowledgments
Special thanks to Natália Souza, Hemille Mariane, Marcelo Carvalho-Júnior, Rafael Oliveira, Verônica Costa, and Leonardo Moraes for their help in fieldwork. We are grateful to Pedro Romano and Ilver Alabat for their suggestions on early version of this manuscript and Jaciene Andrade for reviewing Portuguese grammar. We are also grateful to LABICT/UEFS, LAPAQ/UFSB, and CEPENE-Caravelas teams for allowing the use of their facilities and logistic assistance. JAS thanks Jéssica Prata for the otolith’s photos in the Leica M205 A. This work was partially supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil (CAPES – finance code 001) and Fundação de Amparo à Pesquisa do Estado da Bahia (FAPESB) through the project “Migrações ontogenéticas entre ecossistemas costeiros e o papel de estuários como berçários para espécies de peixes marinhos comerciais através de análise microquímica de otólitos” (PAM 0019/2014). This study is part of JAS Master’s dissertation in Biological Sciences at Universidade Federal da Paraíba.
References
Aguilera OA, Schwarzhans W, Béarez P. Otoliths of the Sciaenidae from the Neogene of tropical America. Palaeo Ichthyo. 2016; 14:7–90.
Aguilera Socorro O. Ophioscion punctatissimus. The IUCN Red list of threatened species 2020: e.T47148202A82680558 [internet]. IUCN; 2020. Available from: https://www.iucnredlist.org/species/47148202/82680558
Alewijnse SR, Stowasser G, Saunders RA, Belcher A, Crimmen OA, Cooper N et al. Otolith-derived field metabolic rates of myctophids (Family Myctophidae) from the Scotia Sea (Southern Ocean). Mar Ecol Prog Ser. 2021; 675:113–31. https://doi.org/10.3354/meps13827
Anjos MS, Bitencourt JA, Nunes LA, Sarmento-Soares LM, Carvalho DC, Armbruster JW et al. Species delimitation based on integrative approach suggests reallocation of genus in Hypostomini catfish (Siluriformes, Loricariidae). Hydrobiologia. 2020; 847:563–78. https://doi.org/10.1007/s10750-019-04121-z
Argolo LA, López-Fernández HL, Batalha-Filho H, Affonso PRAM. Unraveling the systematics and evolution of the ‘Geophagus’ brasiliensis (Cichliformes: Cichlidae) species complex. Mol Phylogenet Evol. 2020; 150:e106855. https://doi.org/10.1016/j.ympev.2020.106855
Assis IO, Silva VEL, Souto-Vieira D, Lozano AP, Volpedo AV, Fabré NN. Ecomorphological patterns in otoliths of tropical fishes: Assessing trophic groups and depth strata preference by shape. Environ Biol Fish. 2020; 103(4):349–61. https://doi.org/10.1007/s10641-020-00961-0
Avigliano E, Rolón ME, Rosso JJ, Mabragaña E, Volpedo AV. Using otolith morphometry for the identification of three sympatric and morphologically similar species of Astyanax from the Atlantic Rain Forest (Argentina). Environ Biol Fish. 2018; 101:1319–28. https://doi.org/10.1007/s10641-018-0779-2
Bani A, Poursaeid S, Tuset VM. Comparative morphology of the sagittal otolith in three species of south Caspian gobies. J Fish Biol. 2013; 82(4):1321–32. https://doi.org/10.1111/jfb.12073
Barbosa AJB, Sampaio I, Schneider H, Santos S. Molecular phylogeny of weakfish species of the Stellifer group (Sciaenidae, Perciformes) of the Western South Atlantic based on mitochondrial and nuclear data. PLoS ONE. 2014; 9(7):e102250. https://doi.org/10.1371/journal.pone.0102250
Byrd BL, Hohn AA, Krause JR. Using the otolith sulcus to aid in prey identification and improve estimates of prey size in diet studies of a piscivorous predator. Ecol Evol. 2020; 10(8):3584–604. https://doi.org/10.1002/ece3.6085
Caires RA, Santos WCR, Machado L, Oliveira C, Cerqueira NNCD, Rotundo MM et al. The Tonkin weakfish, Cynoscion similis (Sciaenidae, Perciformes), an endemic species of the Amazonas-Orinoco Plume. Acta Amaz. 2019; 49(3):197–207. https://doi.org/10.1590/1809-4392201803481
Campana SE, Thorrold SR. Otoliths, increments, and elements: keys to a comprehensive understanding of fish populations? Can J Fish Aquat Sci. 2001; 58(1):30–38. https://doi.org/10.1139/f00-177
Capoccioni F, Costa C, Aguzzi J, Menesatti P, Lombarte A, Ciccotti E. Ontogenetic and environmental effects on otolith shape variability in three Mediterranean European eel (Anguilla anguilla, L.) local stocks. J Exp Mar Biol Ecol. 2011; 397(1):1–07. https://doi.org/10.1016/j.jembe.2010.11.011
Cardoso LG, Santos S, Haimovici M. Differences in the otoliths support the distinction of the genus Macrodon into two species in the south-western Atlantic Ocean. Mar Biodivers Rec. 2012; 5:e93. https://doi.org/10.1017/S1755267212000802
Carstens BC, Pelletier TA, Reid NM, Satler JD. How to fail at species delimitation. Mol Ecol. 2013; 22(17):4369–83. https://doi.org/10.1111/mec.12413
Carvalho BM, Volpedo AV, Fávaro LF. Ontogenetic and sexual variation in the sagitta otolith of Menticirrhus americanus (Teleostei; Sciaenidae) (Linnaeus, 1758) in a subtropical environment. Pap Avulsos Zool. 2020a; 60:e20206009. https://doi.org/10.11606/1807-0205/2020.60.09
Carvalho CO, Marceniuk AP, Oliveira C, Wosiacki WB. Integrative taxonomy of the species complex Haemulon steindachneri (Eupercaria; Haemulidae) with a description of a new species from the western Atlantic. Zoology. 2020b; 141:e125782. https://doi.org/10.1016/j.zool.2020.125782
Carvalho-Filho A, Ferreira CEL, Craig M. A shallow water population of Pronotogrammus martinicensis (Guichenot, 1868) (Teleostei: Serranidae: Anthiinae) from South-western Atlantic, Brazil. Zootaxa. 2009; 2228(1):29–42. https://doi.org/10.11646/zootaxa.2228.1.2
Carvalho-Filho A, Santos S, Sampaio I. Macrodon atricauda (Günther, 1880) (Perciformes: Sciaenidae), a valid species from the southwestern Atlantic, with comments on its conservation. Zootaxa. 2010; 2519(1):48–58. https://doi.org/10.11646/zootaxa.2519.1.3
Chao L. Ophioscion gomezi. The IUCN red list of threatened species 2020: e.T143618198A143619520 [internet]. IUCN; 2020. Available from: https://www.iucnredlist.org/species/143618198/143619520
Chao LN. A basis for classifying western Atlantic Sciaenidae (Pisces: Perciformes). Vol. 415. Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service; 1978.
Chao NL, Carvalho-Filho A, Santos JA. Five new species of Western Atlantic stardrums, Stellifer (Perciformes: Sciaenidae) with a key to Atlantic Stellifer species. Zootaxa. 2021; 4991(3):434–66. https://doi.org/10.11646/zootaxa.4991.3.2
Chao NL, Frédou FL, Haimovici M, Peres MB, Polidoro B, Raseira M et al. A popular and potentially sustainable fishery resource under pressure – extinction risk and conservation of Brazilian Sciaenidae (Teleostei: Perciformes). Glob Ecol Conserv. 2015; 4:117–26. https://doi.org/10.1016/j.gecco.2015.06.002
Chao NL. Sciaenidae. In: Carpenter KE, editor. The living marine resources of the Western Central Atlantic. FAO Species Identification Guide for Fishery Purposes and American Society of Ichthyologists and Herpetologists. Special Publication No 5. Rome: FAO. 2002. p.1583–653.
Clark FJK, Lima CSS, Pessanha ALM. Otolith shape analysis of the Brazilian silverside in two northeastern Brazilian estuaries with distinct salinity ranges. Fish Res. 2021; 243:e106094. https://doi.org/10.1016/j.fishres.2021.106094
Figueiredo-Filho JM, Marceniuk AP, Feijó A, Siccha-Ramirez R, Ribeiro GS, Oliveira C et al. Taxonomy of Centropomus Lacépède, 1802 (Perciformes: Centropomidae), with focus on the Atlantic species of the genus. Zootaxa. 2021; 4942(3):301–38. https://doi.org/10.11646/zootaxa.4942.3.1
Fricke R, Eschmeyer WN, Fong JD. Eschmeyer’s catalog of fishes: genera/species by family/subfamily [Internet]. San Francisco: California Academy of Science; 2022. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Geller JB. Decline of a native mussel masked by sibling species invasion. Conserv Biol. 1999; 13(3):661–64. https://doi.org/10.1046/j.1523-1739.1999.97470.x
Granados-Amores E, Granados-Amores J, Zavala-Leal OI, Flores-Ortega JR. Geometric morphometrics in the sulcus acusticus of the sagittae otolith as tool to discriminate species of the genus Centropomus (Centropomidae: Perciformes) from the southeastern Gulf of California. Mar Biodivers. 2020; 50:e10. https://doi.org/10.1007/s12526-019-01030-1
Guimarães-Costa A, Machado FS, Reis-Filho JA, Andrade MC, Araújo RG, Miranda E et al. DNA Barcoding for the assessment of the taxonomy and conservation status of the fish bycatch of the northern Brazilian shrimp trawl fishery. Front Mar Sci. 2020; 7:566021. https://doi.org/10.3389/fmars.2020.566021
Gunz P, Mitteroecker P. Semilandmarks: a method for quantifying curves and surfaces. Hystrix It J Mamm. 2013; 24(1):103–09. https://doi.org/10.4404/hystrix-24.1-6292
Hervé M. RVAideMemoire: Testing and plotting procedures for biostatistics. 2022. Available from: https://CRAN.R-project.org/package=RVAideMemoire
Hillis DM, Chambers EA, Devitt TJ. Contemporary methods and evidence for species delimitation. Ichthyol Herpetol. 2021; 109(3):895–903. https://doi.org/10.1643/h2021082
Hoff NT, Dias JF, Zani-Teixeira ML, Correia AT. Spatio-temporal evaluation of the population structure of the bigtooth corvina Isopisthus parvipinnis from Southwest Atlantic Ocean using otolith shape signatures. J Appl Ichthyol. 2020; 36:439–50. https://doi.org/10.1111/jai.14044
Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio). Livro vermelho da fauna brasileira ameaçada de extinção: Volume I/1. ed. Brasília: ICMBio; 2018.
Jaramillo AM, Tombari AD, Dura VB, Rodrigo ME, Volpedo AV. Otolith eco-morphological patterns of benthic fishes from the coast of Valencia (Spain). Thalassas. 2014; 30(1):57–66. Available from: http://hdl.handle.net/10251/46138
Jolliffe IT. Principal component analysis and factor analysis. In: Jolliffe IT, editor. Principal Component Analysis. New York: Springer; 1986. p.115–28. https://doi.org/10.1007/978-1-4757-1904-8_7
Jónsson EP, Campana SE, Sólmundsson J, Jakobsdóttir KB, Bárðarson H. The effect of growth rate on otolith-based discrimination of cod (Gadus morhua) ecotypes. PLoS ONE. 2021; 16(9):e0247630. https://doi.org/10.1371/journal.pone.0247630
Kikuchi E, Cardoso LG, Canel D, Timi JT, Haimovici M. Using growth rates and otolith shape to identify the population structure of Umbrina canosai (Sciaenidae) from the Southwestern Atlantic. Mar Biol Res. 2021; 17(3):272–85. https://doi.org/10.1080/17451000.2021.1938131
Klingenberg CP. Walking on Kendall’s shape space: understanding shape spaces and their coordinate systems. Evol Biol. 2020; 47:334–52. https://doi.org/10.1007/s11692-020-09513-x
La Mesa M, Guicciardi S, Donato F, Riginella E, Schiavon L, Papetti C. Comparative analysis of otolith morphology in icefishes (Channichthyidae) applying different statistical classification methods. Fish Res. 2020; 230:105668. https://doi.org/10.1016/j.fishres.2020.105668
Libungan LA, Pálsson S. ShapeR: An R package to study otolith shape variation among fish populations. PLoS ONE. 2015; 10(3):e0121102. https://doi.org/10.1371/journal.pone.0121102
Lira AS, Le Loc’h F, Andrade HA, Lucena-Frédou F. Vulnerability of marine resources affected by a small-scale tropical shrimp fishery in Northeast Brazil. ICES J Mar Sci. 2022; 79(3):633–47. https://doi.org/10.1093/icesjms/fsac004
Lo PC, Liu SH, Chao NL, Nunoo FKE, Mok HK, Chen WJ. A multi-gene dataset reveals a New World origin and Oligocene diversification of croakers (Perciformes: Sciaenidae). Mol Phylogenet Evol. 2015; 88:132–43. https://doi.org/10.1016/j.ympev.2015.03.025
Lombarte A, Chic Ò, Parisi-Baradad V, Olivella R, Piera J, García-Ladona E. A web-based environment for shape analysis of fish otoliths. The AFORO database. Sci Mar. 2006; 70:147–52. https://doi.org/10.3989/scimar.2006.70n1147
Lombarte A, Palmer M, Matallanas J, Gómez-Zurita J, Morales-Nin B. Ecomorphological trends and phylogenetic inertia of otolith sagittae in Nototheniidae. Environ Biol Fish. 2010; 89:607–18. https://doi.org/10.1007/s10641-010-9673-2
Marceniuk AP, Burlamaqui TCT, Oliveira C, Carneiro J, Eleres B, Sales JBL. Incipient speciation, driven by distinct environmental conditions, in the marine catfishes of the genus Aspistor (Siluriformes, Ariidae), from the Atlantic coast of South America. J Zool Syst Evol Res. 2019; 57:1–18. https://doi.org/10.1111/jzs.12261
Márquez-Velásquez V, Raimundo RLG, Rosa RS, Navia AF. The use of ecological networks as tools for understanding and conserving marine biodiversity. In: Ortiz M, Jordán F, editors. Marine Coastal Ecosystems Modelling and Conservation. Cham: Springer; 2021. p.179–202. https://doi.org/10.1007/978-3-030-58211-1_9
Meek SE, Hildebrand SF. The marine fishes of Panama. Part II. Field Museum of Natural History Publications 226. Zoological Series 15. Chicago: Field Museum of Natural History; 1925.
Pavlov DA. Differentiation of three species of the genus Upeneus (Mullidae) based on otolith shape analysis. J Ichthyol. 2016; 56(1):37–51. https://doi.org/10.1134/S0032945216010094
Popper AN, Ramcharitar J, Campana SE. Why otoliths? Insights from inner ear physiology and fisheries biology. Mar Freshw Res. 2005; 56:497–504. https://doi.org/10.1071/MF04267
R Development Core Team. R: A language and environment for statistical computing. [internet] R Foundation for Statistical Computing. 2020. Available from: https://www.R-project.org/
Ramcharitar JU, Higgs DM, Popper AN. Sciaenid inner ears: a study in diversity. Brain Behav Evol. 2001; 58:152–62. https://doi.org/10.1159/000047269
Riddle BR, Ladle RJ, Lourie SA, Whittaker RJ. Basic biogeography: estimating biodiversity and mapping nature. In: Ladle RJ, Whittaker RJ, editors. Conservation Biogeography. Oxford: Wiley; 2011. p.45–92. https://doi.org/10.1002/9781444390001.ch4
Rodrigues VLA, Wedekin LL, Marcondes MCC, Barbosa L, Farro APC. Diet and foraging opportunism of the Guiana dolphin (Sotalia guianensis) in the Abrolhos Bank, Brazil. Mar Mamm Sci. 2020; 36(2):436–50. https://doi.org/10.1111/mms.12656
Rohlf FJ. tpsDig2 v.2.27. State University of New York at Stony Brook. 2017a.
Rohlf FJ. tpsRelw, v. 1.69. State University of New York at Stony Brook. 2017b.
Rohlf FJ. tpsUtil v.1.69. State University of New York at Stony Brook. 2021.
Santos JA, Oliveira RL, Guedes APP, Santos ACA, Moraes LE. Do macrophytes act as restaurants for fishes in a tropical beach? An approach using stomach content and prey availability analyses. Reg Stud Mar Sci. 2021; 47:e101920. https://doi.org/10.1016/j.rsma.2021.101920
Santos S, Hrbek T, Farias IP, Schneider H, Sampaio I. Population genetic structuring of the king weakfsh, Macrodon ancylodon (Sciaenidae), in Atlantic coastal waters of South America: deep genetic divergence without morphological change. Mol Ecol. 2006; 15:4361–73. https://doi.org/10.1111/j.1365-294X.2006.03108.x
Schneider C, Rasband W, Eliceiri K. NIH Image to ImageJ: 25 years of image analysis. Nat Methods. 2012; 9:671–75. https://doi.org/10.1038/nmeth.2089
Schulz-Mirbach T, Ladich F, Plath M, Heâ M. Enigmatic ear stones: what we know about the functional role and evolution of fish otoliths. Biol Rev. 2019; 94:457–82. https://doi.org/10.1111/brv.12463
Schwarzhans W. A comparative morphological treatise of recent and fossil otoliths of the family Sciaenidae (Perciformes). Piscium Catalogus, Otolithi Piscium, 1993; 1:1–245.
Silva TF, Schneider H, Sampaio I, Angulo A, Brito MFG, Santos ACA et al. Phylogeny of the subfamily Stelliferinae suggests speciation in Ophioscion Gill, 1863 (Sciaenidae: Perciformes) in the western South Atlantic. Mol Phylogenet Evol. 2018; 125:51–61. https://doi.org/10.1016/j.ympev.2018.03.025
Struck TH, Cerca J. Cryptic species and their evolutionary significance. eLS. Chichester: John Wiley & Sons, Ltd; 2019. https://doi.org/10.1002/9780470015902.a0028292
Trewavas E. The sciaenid fishes (croakers or drums) of the Indo-West-Pacific. Trans Zool Soc London. 1977; 33:253–541. https://doi.org/10.1111/j.1096-3642.1977.tb00052.x
Tuset VM, Farré M, Otero-Ferrer JL, Vilar A, Morales-Nin B, Lombarte A. Testing otolith morphology for measuring marine fish biodiversity. Mar Freshw Res. 2016; 67:1037–48. https://doi.org/10.1071/MF15052
Tuset VM, Galimay E, Farrés A, Marco-Herreno E, Otero-Ferrer JL, Lombarte A et al. Recognising mollusc shell contours with enlarged spines: Wavelet vs Elliptic Fourier analyses. Zoology. 2020; 140:125778. https://doi.org/10.1016/j.zool.2020.125778
Tuset VM, Otero-Ferrer JL, Siliprandi C, Manjabacas A, Marti-Puig P, Lombarte A. Paradox of otolith shape indices: routine but overestimated use. Can J Fish Aquat Sci. 2021; 78(6):681–92. https://doi.org/10.1139/cjfas-2020-0369
Venables WN, Ripley BD. Modern Applied Statistics with S. 4th ed. New York: Springer; 2002. https://doi.org/10.1007/978-0-387-21706-2
Wong JY, Chu C, Chong VC, Dhillon SK, Loh KH. Automated otolith image classification with multiple views: an evaluation on Sciaenidae. J Fish Biol. 2016; 89(2):1324–44. https://doi.org/10.1111/jfb.13039
Zischke MT, Litherland L, Tilyard BR, Stratford NJ, Jones EL, Wang YG. Otolith morphology of four mackerel species (Scomberomorus spp.) in Australia: species differentiation and prediction for fisheries monitoring and assessment. Fish Res. 2016; 176:39–47. https://doi.org/10.1016/j.fishres.2015.12.003
Authors

![]() Jonas de Andrade Santos1
Jonas de Andrade Santos1 ![]() ,
, ![]() Rafael Menezes1,
Rafael Menezes1, ![]() Ricardo de Souza Rosa2 and
Ricardo de Souza Rosa2 and ![]() Telton Pedro Anselmo Ramos2
Telton Pedro Anselmo Ramos2
[1] Programa de Pós-Graduação em Ciências Biológicas (Zoologia), Centro de Ciências Exatas e da Natureza, UniversidadeFederal da Paraíba, 58051-900 João Pessoa, PB, Brazil. (JAS) jonasandradebio@gmail.com (corresponding author), (RM) rafaelmenez@gmail.com.
[2] Departamento de Sistemática e Ecologia, Centro de Ciências Exatas e da Natureza, Universidade Federal da Paraíba, 58051-900 João Pessoa, PB, Brazil. (RSR) rsrosa@dse.ufpb.br, (TPAR) telton@gmail.com.
Authors’ Contribution 

Jonas de Andrade Santos: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing-original draft, Writing-review and editing.
Rafael Menezes: Conceptualization, Formal analysis, Methodology, Software, Visualization, Writing-original draft, Writing-review and editing.
Ricardo de Souza Rosa: Conceptualization, Methodology, Project administration, Supervision, Validation, Writing-review and editing.
Telton Pedro Anselmo Ramos: Conceptualization, Methodology, Project administration, Supervision, Validation, Writing-review and editing.
Ethical Statement
The samplings were authorized by Sistema de Autorização e Informação em Biodiversidade (Collection Permit SISBIO 47993–1).
Competing Interests
The authors declare no competing interests.
How to cite this article
Santos JA, Menezes R, Rosa RS, Ramos TPA. Otolith shape analysis supports three cryptic species in the Stellifer punctatissimus complex (Acanthuriformes: Sciaenidae). Neotrop Ichthyol. 2022; 20(3):e220005. https://doi.org/10.1590/1982-0224-2022-0005
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2022 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted August 19, 2022 by Osmar Luiz
Accepted August 19, 2022 by Osmar Luiz
![]() Submitted January 17, 2022
Submitted January 17, 2022
![]() Epub October 14, 2022
Epub October 14, 2022