![]() Priscila Polaquini de Macedo Leite1
Priscila Polaquini de Macedo Leite1 ![]() ,
, ![]() Francisco de Menezes Cavalcante Sassi1,
Francisco de Menezes Cavalcante Sassi1, ![]() Manoela Maria Ferreira Marinho2,
Manoela Maria Ferreira Marinho2, ![]() Mauro Nirchio3,
Mauro Nirchio3, ![]() Renata Luiza Rosa de Moraes,
Renata Luiza Rosa de Moraes, ![]() Gustavo Akira Toma1,
Gustavo Akira Toma1, ![]() Luiz Antonio Carlos Bertollo1 and
Luiz Antonio Carlos Bertollo1 and ![]() Marcelo de Bello Cioffi1
Marcelo de Bello Cioffi1
PDF: EN XML: EN | Cite this article
Abstract
Despite several difficulties in chromosomal analyses of small-sized fishes, the cytogenetics of the Lebiasinidae was largely improved in the last years, showing differential patterns in the chromosomal evolution inside the family. In this context, it has been shown that genus Lebiasina preserves its karyotypic macrostructure, composed of 2n = 36 chromosomes, whereas the other genera generally present higher 2n. This study focused on the comparative cytogenetics of three Lebiasina species, one of them analyzed here for the first time, using conventional and molecular procedures. The results reinforced the differentiated evolutionary path of the genus Lebiasina while, at the same time, highlighted the genomic particularities that have accompanied the evolution of each species. In this sense, the repetitive components of the genome played a significant role in the differentiation of each species. It is also notable that L. minuta and L. melanoguttata, the two species that occur exclusively in the Brazilian territory, show greater chromosomal similarities to each other than to the trans-Andean sister species, L. bimaculata.
Keywords: Chromosomal Evolution, Cytogenetics, FISH, Comparative Genomic Hybridization, Lebiasininae.
Apesar das dificuldades encontradas em se realizar análises cromossômicas em peixes de pequeno porte, os estudos citogenéticos em Lebiasinidae vêm crescendo nos últimos anos e demonstrando padrões diferenciados na evolução cromossômica entre os membros da família. Nesse contexto, o gênero Lebiasina tem mostrado preservar sua macroestrutura cariotípica, composta por 2n = 36 cromossomos, enquanto os demais gêneros geralmente apresentam 2n maiores. Este estudo tem como foco a citogenética comparativa de três espécies de Lebiasina, sendo uma delas analisada pela primeira vez aqui, através do emprego de técnicas convencionais e moleculares. Os resultados obtidos reforçam a trajetória evolutiva diferenciada do gênero Lebiasina, ao mesmo tempo em que evidenciam as particularidades genômicas que acompanham a evolução de cada uma das espécies. Neste contexto, os componentes repetitivos do genoma tiveram um papel importante na caracterização particular de cada uma das espécies. Também, é notável que L. minuta e L. melanoguttata,duas espécies que ocorrem exclusivamente no território brasileiro, apresentam maior proximidade citogenética entre elas do que com a espécie irmã transandina, L. bimaculata.
Palavras-chave: Citogenética, Evolução Cromossômica, Hibridização Genômica Comparativa, FISH, Lebiasininae.
Introduction
Lebiasinidae is a well-supported monophyletic taxon containing seven genera and 75 valid species distributed in two subfamilies, Lebiasininae and Pyrrhulininae (Weitzman, Weitzman, 2003; Netto-Ferreira, Marinho, 2013; Fricke et al., 2021). Lebiasininae encompasses three genera, Lebiasina Valenciennes, 1847 (26 sp.), Piabucina Valenciennes, 1850 (1 sp.) and Derhamia Géry & Zarske, 2002 (1 sp.), and Pyrrhulininae other four ones, Nannostomus Günther, 1872 (21 sp.), Pyrrhulina Valenciennes, 1846 (19 sp.), Copella Myers, 1956 (6 sp.), and Copeina Fowler, 1906 (2 sp.) (Netto-Ferreira, Marinho, 2013; Fricke et al., 2021). All lebiasinids are found in freshwater and are endemic to Central and South Americas, except in Chile’s hydrographic basins. They are characterized by the absence of adipose fin, small anal fins, and an elongated body, which varies greatly in size, from 1.6 cm in Nannostomus to 20 cm in Lebiasina (Weitzman, Weitzman, 2003; Netto-Ferreira, 2010).
The first morphological investigations reported that Lebiasinidae would be related to Erythrinidae, Ctenoluciidae, Serrasalmidae, and Hepsetidae (Ortí, Meyer, 1997; Buckup, 1998; Oyakawa, 1998). However, molecular data suggested that many morphological synapomorphies among the abovementioned groups could be convergent evolutionary traits associated with their predatory lifestyle but positioning Lebiasinidae as a sister group to Ctenoluciidae (Oliveira et al., 2011). Lately, additional data from ultraconserved elements (UCEs) also corroborated their phylogenetic relatedness (Arcila et al., 2017; Betancur‐R. et al., 2019; Melo et al., 2022).
Thus far, only three Lebiasina species were found to occur in Brazilian waters at Serra do Cachimbo (PA): Lebiasina marilynae Netto-Ferreira, 2012, L. melanoguttata Netto-Ferreira, 2012, and L. minuta Netto-Ferreira, 2012, with a fourth additional species, Lebiasina yepezi Netto-Ferreira, Oyakawa, Zuanon & Nolasco, 2011, found in the Brazil-Venezuela border (Netto-Ferreira et al., 2011; Netto-Ferreira, 2012). Cytogenetic analyses pointed that the diploid number (2n = 36) is conserved for L. bimaculata Valenciennes, 1847,and L. melanoguttata, the only two species for which chromosomal data are known (Sassi et al., 2019). Significantly, this number is also conserved in all Ctenoluciidae representatives (de Souza e Sousa et al., 2017; Souza et al., 2021). In fact, two general trends appear to occur within the Lebiasinidae family: i) species with 2n = 36 bi-armed chromosomes, as in Lebiasina (Sassi et al., 2019) and some Nannostomus species (Sember et al., 2020), and ii) with higher diploid numbers, with mostly mono-armed chromosomes, as in Pyrrhulina (de Moraes et al., 2017, 2019, 2021), Copeina (Toma et al., 2019) and Nannostomus species (Sember et al., 2020). Up to now, Pyrrhulina semifasciata Steindachner, 1876 is the only species in the family that displays a morphologically differentiated sex chromosome system, of the X1X1X2X2/X1X2Y type (de Moraes et al., 2019).
The present study aimed to extend the knowledge on the trends and underlying mechanisms of karyotype differentiation in Lebiasinidae. Our main goal was to characterize the chromosomal patterns of the species Lebiasina minuta and highlight the contrasting evolutionary pathways inside the genus Lebiasina. Besides, we will test the hypothesis that a karyotype composed of 36 exclusively bi-armed chromosomes is also shared by other Lebiasina species, representing thus, a probable synapomorphy for this genus. For this, we applied conventional (Giemsa staining, C-banding) and molecular (mapping of repetitive DNA markers, comparative genomic hybridization (CGH), and whole chromosome painting (WCP)) in three Lebiasina species, one of them (L. minuta) now analyzed for the first time. This study is included in a series focusing on the cytogenetics and cytogenomics of Lebiasinidae fishes.
Material and methods
Sampling, chromosomes obtainment, and C-banding. Samples of Lebiasina minuta were collected at Serra do Cachimbo, Xingu River basin (Fig. 1; Tab. 1).The samples of L. bimaculata and L. melanoguttata were the same used by Sassi et al. (2019). The specimens were properly identified by morphological and meristic criteria by Dr. Manoela M. F. Marinho, an expert on Lebiasinidae taxonomy and were deposited in the fish collection of the Museu de Zoologia da Universidade de São Paulo (voucher number MZUSP 126519). The map was designed using the software QGIS Desktop 3.18 and Adobe CC Photoshop 2020.

FIGURE 1| Distribution of Lebiasina species with available cytogenetic data, highlighting the Brazilian state of Pará (orange) and Ecuadorian (purple) territories A. 1. L. bimaculata,2. L. melanoguttata (Sassi et al., 2019), and 3. L. minuta (this study). B. Highlights the position of A in South America, and C. indicates that, although close, species 2 and 3 does not share an overlapped distribution.
TABLE 1 | Sample sites, geographic coordinates, sampling number (N), diploid number (2n) and distribution of ribosomal DNA sequences on chromosomes of Lebiasina species.
Location | Geographic
coordinates | N | 2n | 5S rDNA | 18S rDNA | References | |
Lebiasina bimaculata | Arenillas river lakes, El Oro – Ecuador | 03°30’57.204’’S
80°03’44.2656’’W | 03♀, 04♂ | 36 (36m/sm) | Pair 1 | Pair 3 | Sassi et al.
(2019) |
Lebiasina melanoguttata | Serra do Cachimbo, Cachoeira da Serra – PA, Brazil | 08°58’18.7’’S
54°58’18.77’’W | 22♀, 14♂ | 36 (36m/sm) | Pairs 1 and 13 | Pairs 1, 2, 3, 7 and
9 | Sassi et al.
(2019) |
Lebiasina minuta | PCH Três de Maio, Cachoeira da Serra – PA, Brazil | 08°44’39’’S
55°02’03’’W | 10♀, 08♂ | 36 (36m/sm) | Pairs 1 and 13 | Pair 1 | Present study |
Mitotic chromosomes were obtained from anterior kidney cells employing the classical air-drying method (Bertollo et al., 2015). Chromosomes were stained with 10% Giemsa diluted in Sorensen phosphate buffer (pH 6.8) and the constitutive heterochromatin regions were evidenced through the C-banding protocol (Sumner, 1972).
Probe obtainment and FISH-based experiments. Both 18S and 5S ribosomal DNA sequences (rDNAs) were cytogenetically mapped by FISH, using Hoplias malabaricus Bloch, 1794 (Characiformes, Erythrinidae) genome-isolated sequences. The 18S rDNA probe corresponds to a 1,400 base pairs (bp) segment of the respective gene (Cioffi et al., 2009), and the 5S rDNA probe includes 120 bp of the respective gene plus 200pb of non-transcribed spacers – NTS (Pendás et al., 1994). Both probes were labeled using a Nick-translation kit (Jena Bioscience, Jena, Germany), the 5S rDNA being labeled with Atto550-dUTP (red color) and 18S rDNA with Atto448-dUTP (green color). Three microsatellite sequences – (GA)15, (CA)15, (CGG)10 – that showed accumulation in other Lebiasinidae species previously analyzed (de Moraes et al., 2017, 2019; Sassi et al., 2019; Toma et al., 2019), were directly labeled during their synthesis (Kubat et al., 2008) with Cy-3 (Sigma-Aldrich, Darmstadt, Germany),and mapped on chromosomes, in addition to the telomeric sequence (TTAGGG)n using the Telomere PNA FISH Kit/FITC (DAKO, Glostrup, Denmark).
Microdissected first chromosome pair of Lebiasina bimaculata was used for whole chromosome painting (WCP) (Sassi et al., 2019) and labeled with Spectrum-Orange dUTP fluorophore (Vysis Inc, EUA). Two sets of comparative genome hybridizations (CGH) were also designed. The first one aimed to compare the genomic content of all analyzed Lebiasina species. For that, the male genomic DNAs (gDNAs) of L. bimaculata, L. melanoguttata, and L. minuta, were extracted from liver tissues (Sambrook, Russell, 2001), labeled by Nick-translation with Atto425-dUTP (light blue), Atto488-dUTP (green), and Atto550-dUTP (red) (Jena Biosciences, Jena, Germany), respectively, and co-hybridized against the male chromosomal background of L. minuta, using C0t-1 DNA (i.e., part of genomic DNA enriched for highly repetitive sequences) as a blocker of excessed shared repetitive sequences (Zwick et al., 1997). The final hybridization mixture was composed of 500ng of L. minuta gDNA, 500ng of the each compared gDNAs, and 25μl of unlabeled C0t-1 DNA, in a hybridization buffer containing 50% of formamide, 2× SSC, 10% SDS, 10% dextran sulfate and Denhardt´s buffer (pH = 7.0). The second experiment focused on intraspecific variations between males and females of L. minuta. Male and female-derived gDNA were also obtained by the standard phenol:chloroform:isoamyl alcohol protocol (Sambrook, Russell, 2001), and labeled with Atto550-dUTP (red), and Atto488-dUTP (green), respectively. A male metaphase preparation was used to co-hybridize both genomes. The final hybridization mix was composed of 500ng of male-derived gDNA, plus 500ng of female-derived gDNA and 15µg of unlabelled male-derived C0t-1 DNA. The chosen ratio of probes versus the C0t-1 DNA amount was based on previous data of our research group (Sassi et al., 2020). In all FISH-base experiments, chromosomes were counterstained with 4’,6-diamidino-2-phenylindole (DAPI) and slides mounted with an antifade solution (VECTASHIELD, Vector Laboratories, Burlingame, CA, USA). All the hybridizations procedures followed the high-stringency protocol described in Yano et al. (2017).
Optical analyses and image processing. Metaphase plates were captured at a photomicroscope Olympus BX50 with CoolSNAP and images processed by the software ImagePro Plus 4.1. Chromosomes were classified according to their arms ratio (q/p), following Levan et al. (1964). Karyotypes were assembled with Adobe Photoshop CC 2020 software after the analysis of at least 30 metaphases for each sex to confirm the 2n number, karyotype structure and FISH results.
Results
Conventional data and repetitive DNA mapping. Both males and females of L. minuta have 2n = 36 meta- and submetacentric chromosomes without heteromorphic sex chromosomes (Fig. 2A). C-positive heterochromatin occurs at the pericentromeric regions in all chromosomes (Fig. 2B). The “double-FISH” procedure showed a syntenic condition for both 5S and 18S rDNA sequences in the long (q) arms of the first chromosomal pair of males and females (Fig. 2C) with the 18S rDNA located on the terminal region, whereas 5S rDNA site is found in the pericentromeric region. Additional 5S rDNA sites are also found in the short (p) arms of the 13th chromosomal pair (Fig. 2C).

FIGURE 2| Male and female karyotypes of Lebiasina minuta after A. Giemsa staining, B. C-banding, and C. “double-FISH” with 5S (red) and 18S (green) rDNA probes. Scale bar = 5 µm.
Microsatellites (CA)15 and (GA)15 marks are mainly found in almost all chromosomes of L.minuta (Figs. 3A–B), while (CGG)10 marks occur only in the q terminal region of the first chromosome pair (Fig. 3C). Telomeric sequences (TTAGGG)n were only identified in their standard terminal regions in all chromosomes (Fig. 3D), without interstitial telomeric sites (ITS).

FIGURE 3| Metaphase chromosomes of Lebiasina minuta hybridized with microsatellite probes (A, B and C) and telomeric probes (D), using red signals. Scale bar = 5 µm.
Whole chromosome painting (WCP). The whole chromosome painting (WCP) with a derived probe from the first chromosome pair of L. bimaculata completely hybridized the first chromosome pair of L. minuta, with small-scattered signals in other chromosomes (Fig. 4).

FIGURE 4| Whole chromosome painting (WCP) highlighting the first chromosome pair of Lebiasina minuta completely hybridized with the probe from the first chromosome pair of L. bimaculata.
Intra- and interspecific comparative genomic hybridization (CGH). The genomic probes of L. bimaculata and L. melanoguttata successfully hybridized with the chromosomes of L. minuta. Notably, the Brazilian species L. melanoguttata and L. minuta share more repetitive sequences in their genomes than with the Ecuadorian species L. bimaculata, especially at the pericentromeric regions (Figs. 5A–D). The intraspecific comparison did not identify any sex-specific region, thus discarding the occurrence of distinguishable sex chromosomes (Figs. 5E–H).

FIGURE 5| First Row: Mitotic chromosome spreads of Lebiasina minuta males after CGH— interspecific comparisons (A–D). Male-derived genomic probe of L. minuta (A); L. melanoguttata (B); L. bimaculata (C) hybridized against male metaphase plates of L. minuta (D). Second Row: Mitotic chromosome spreads of Lebiasina minuta males after CGH— intraspecific comparisons (E–H). DAPI image (E); Male-derived genomic probe of L. minuta (F); Female-derived genomic probe of L. minuta (G) hybridized against male metaphase plates of L. minuta (H). The common genomic regions of both compared karyomorphs are depicted in yellow. Scale bar = 5 µm.
Discussion
General chromosomal conservation. The current results show that the presence of 2n = 36 biarmed chromosomes is a general conserved characteristic in all the Lebiasina species analyzed. Such feature is also found in some other lebiasinids, such as Nannostomus eques Steindachner, 1876(Sember et al., 2020), but notably in all representatives of the Ctenoluciidae family, which is represented by the genera Boulengerella Eigenmann, 1903(Souza et al., 2021)and Ctenolucius Gill, 1861 (Souza et al., 2021). Such chromosomal relationship between the Lebiasinidae and Ctenoluciidae families, taken as sister groups (Arcila et al., 2017; Betancur‐R. et al., 2019), indicates that 2n = 36 biarmed chromosomes is a plesiomorphic condition within lebiasinids (Sassi et al., 2020). Thus, while Lebiasininae has retained the 2n = 36 biarmed chromosomes throughout its evolutionary history, the subfamily Pyrrhulininae followed a very different pathway, its species differing by presenting larger diploid numbers and mainly acrocentric chromosomes. Therefore, our results further support the evolutionary differentiation within the Lebiasinidae family by introducing new data on L. minuta.
Another shared characteristic refers to the first pair of chromosomes among the Lebiasina species, which can be easily differentiated, as it is the largest metacentric of the karyotype. Besides its morphological conservation, all three species also share a general genomic composition for this chromosome pair, as evidenced by the WCP experiments (Fig. 4). Indeed, this occurrence does not only refer to L. bimaculata and L. minuta as shown in the current study, but also to L. bimaculata and L. melanoguttata (Sassi et al., 2019). Therefore, this chromosome stands out as a possible useful marker for further investigations regarding the evolutionary process in the genus Lebiasina, as well as in the Lebiasinidae as a whole.
Chromosomal and genomic evolutionary differentiation. Despite the general conservation of the diploid number and chromosomal morphology, some particular features highlight the evolutionary differentiation that was fixed by each of the Lebiasina species, both at the chromosomal and genomic levels. This can be firstly easily verified concerning the distribution pattern of the constitutive heterochromatin (Fig. 2). It was previously shown that L. bimaculata and L. melanoguttata present centromeric and telomeric sites in several chromosome pairs, in addition to an exclusive set of noticeable interstitial bands in L. melanoguttata (Sassi et al., 2019). In turn, L. minuta differs from its sister species by having only pericentromeric sites, both in male and female karyotypes. It is known that heterochromatin has an important role in maintaining the chromosome structure and, consequently, it is usually associated with some specific regions, such as centromeres and telomeres. However, different distribution patterns are often found, even among closely related species as in Lebiasina, indicating probable additional roles played by the heterochromatin throughout the evolutionary pathways of the different species. In fact, the heterochromatic regions are composed by various types of repetitive sequences (Charlesworth et al., 1994; Kidwell, 2002) and, therefore, act as hotspots for chromosomal repatterning processes, driving an intragenomic dynamism during the evolutionary process of many fish species (Cioffi, Bertollo, 2012).
In fact, the distribution of repetitive DNAs represents a powerful tool in exploring the genome dynamics in fishes (Cioffi, Bertollo, 2012), which can be observed regarding the distribution of the rDNA sites among the Lebiasina species (Fig. 6). Concerning the 5S rDNA, all species share a site on the first chromosome pair, although in a different position and condition in L. bimaculata, including an additional site on chromosome 13 in L. melanoguttata and L. minuta. In turn, the 18S rDNA has a very different distribution among species. A single site is found in L. bimaculata and L. minuta, on chromosome pairs three and one, respectively, while L. melanoguttata has several sites distributed in the karyotype, including a bi-telomeric one in pair two (Fig. 6). Thus, the 18S rDNA presents a greater evolutionary dynamism among species compared to that noticed for the 5S. However, regardless of their numerical dispersion, it is notable that all 18S sites occupy a terminal position on chromosomes, which can be considered as a symplesiomorphic trait for lebiasinids, since Nannostomus, Pyrrhulina, Lebiasina, Copeina, and the representatives of the sister family Ctenoluciidae, also share such feature (Sember et al., 2020). In the same way, the syntenic condition for both 18S and 5S rDNAs in the first chromosomal pair of L. minuta and L. melanoguttata also occurs in other lebiasinids karyotypes, such as Pyrrhulina australis Eigenmann & Kennedy, 1903, Pyrrhulina aff. australis, P. brevis Steindachner, 1876 and Pyrrhulina cf. laeta (Cope, 1872) (de Moraes et al., 2017, 2019, 2021). This fact represents an exception among fishes, since a non-syntenic organization for both rDNA classes has been originally assumed to be the plesiomorphic condition for this group (Amemiya, Gold, 1988; Gornung, 2013). Such a syntenic condition can create recombination hotspots in association with heterochromatin (Gornung, 2013; Sochorová et al., 2018), facilitating intrachromosomal rearrangements, as observed in mice and humans (Cazaux et al., 2011; Tchurikov et al., 2021).
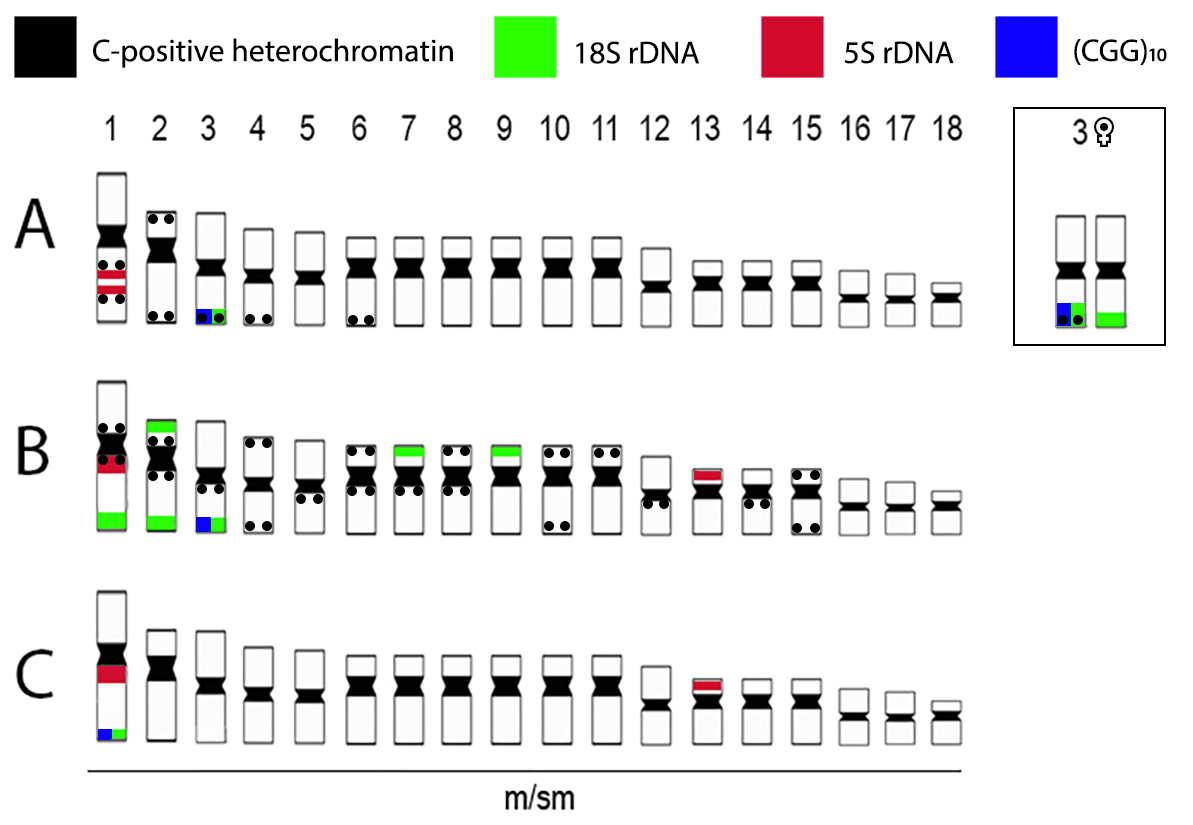
FIGURE 6 | Representative idiograms of L. bimaculata (A); L. melanoguttata (B) and L. minuta (C) highlighting the distribution of the 18S (green) and 5S (red) rDNA sequences; (CGG)n microsatellite (blue) and C-positive heterochromatin (black): Data for L. bimaculata and L. melanoguttata are from Sassi et al. (2019).
Although sharing the same 2n, CGH experiments (Fig. 5) and the microsatellite distribution (Fig. 3) suggest an advanced stage of sequence divergence among the Lebiasina species under study. Such internal reorganization in chromosomes is likely to be related to less identified rearrangements in fish karyotypes and has a marked role in the karyotype evolution of several animal groups (Matsuoka et al., 2004; Barby et al., 2019). Our results indicate that repetitive sequences have divergent patterns of distribution and accumulation, probably fostering the chromosomal differentiation and biodiversity, thus highlighting the differential paths taken by the evolutionary process when comparing the genome organization of the trans-Andean species, L. bimaculata, with the two exclusive Brazilian species, L. minuta and L. melanoguttata.
In spite of the difficulties in obtaining good chromosomal preparations for small-sized fish, the cytogenetics of the Lebiasinidae family has experienced considerable progress in recent years, both on conventional and molecular procedures. Our current data on L. minuta support that 2n = 36 two-armed chromosomes is a plesiomorphic condition for the genus Lebiasina, reinforcing its proximity to Ctenoluciidae species as a sister group. However, despite the maintenance of a general karyotypic macrostructure, these species highlight differential evolutionary features regarding the distribution of repetitive elements of the genome, indicating their dynamic in their genomic differentiation pathways. It is also notable that the distribution and amplification of repetitive DNA classes across the chromosomes followed independent evolutionary trajectories among the Lebiasina species. The two exclusively Brazilian species, L. minuta and L. melanoguttata, are more related to each other, sharing a more genomic closeness than with the trans-Andean species, L. bimaculata. In fact, allopatry is often considered the most common source of speciation among Neotropical fishes (Seehausen, Wagner, 2014) and here may have contributed to the biodiversity of such Lebiasinidae fishes.
Acknowledgments
This work was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq – 302449/2018–3 to MBC); Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP – 2020/11772–8 to MBC, 2020/03046–5 to PPML, 2020/02681–9 to FMCS). This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brasil (CAPES – Finance code 001).
References
Amemiya CT, Gold JR. Chromosomal NORs as taxonomic and systematic characters in North American cyprinid fishes. Genetica. 1988; 76(2):81–90. https://doi.org/10.1007/BF00058806
Arcila D, Ortí G, Vari R, Armbruster JW, Stiassny MLJ, Ko KD et al. Genome-wide interrogation advances resolution of recalcitrant groups in the tree of life. Nat Ecol Evol. 2017; 1(2):0020. https://doi.org/10.1038/s41559-016-0020
Barby FF, Bertollo LAC, Oliveira EA, Yano CF, Hatanaka T, Ráb P et al. Emerging patterns of genome organization in Notopteridae species (Teleostei, Osteoglossiformes) as revealed by Zoo-FISH and Comparative Genomic Hybridization (CGH). Sci Rep. 2019; 9(1):1112. https://doi.org/10.1038/s41598-019-38617-4
Bertollo LAC, Cioffi MB, Moreira-Filho O. Direct chromosome preparation from freshwater teleost fishes. In: Ozouf-Costaz C, Pisano E, Foresti F, Toledo LFA, editors. Fish Cytogenetic Techniques: Ray-fin fishes and Chondrichthyans.Boca Raton: CRC Press; 2015. p.21–26. https://doi.org/10.1201/b18534-6
Betancur-R. R, Arcila D, Vari RP, Hughes LC, Oliveira C, Sabaj MH et al. Phylogenomic incongruence, hypothesis testing, and taxonomic sampling: The monophyly of characiform fishes. Evolution (NY). 2019; 73(2):329–45. https://doi.org/10.1111/evo.13649
Bloch ME. Naturgeschichte der ausländischen Fische. Acther theil. Berlin; 1794.
Buckup PA. Relationships of the Characidiinae and phylogeny of characiform fishes (Teleostei: Ostaryorphysi). In: Malabarba LR, Reis RE, Vari RP, Lucena ZMS, Lucena CAS, editors. Phylogeny and classification of Neotropical fishes. Porto Alegre: EDIPUCRS; 1998. p.123–44.
Cazaux B, Catalan J, Veyrunes F, Douzery EJP, Britton-Davidian J. Are ribosomal DNA clusters rearrangement hotspots? A case study in the genus Mus (Rodentia, Muridae). BMC Evol Biol. 2011; 11(1):124. https://doi.org/10.1186/1471-2148-11-124
Charlesworth B, Sniegowski P, Stephan W. The evolutionary dynamics of repetitive DNA in eukaryotes. Nature. 1994; 371(6494):215–20. https://doi.org/10.1038/371215a0
Cioffi MB, Martins C, Centofante L, Jacobina U, Bertollo LAC. Chromosomal variability among allopatric populations of Erythrinidae fish Hoplias malabaricus: Mapping of three classes of repetitive DNAs. Cytogenet Genome Res. 2009; 125(2):132–41. https://doi.org/10.1159/000227838
Cioffi MB, Bertollo LAC. Chromosomal distribution and evolution of repetitive DNAs in fish. In: Garrido-Ramos MA, editor. Repetitive DNA. Genome Dynamics; Vol. 7. Basel: Karger; 2012. p.197–221. https://doi.org/10.1159/000337950
Fricke R, Eschmeyer WN, Van der Laan R. Eschmeyer’s catalog of fishes: Genera, species, references [Internet]. San Francisco: California Academy of Sciences; 2021. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Gornung E. Twenty years of physical mapping of major ribosomal RNA genes across the teleosts: A review of research. Cytogenet Genome Res. 2013; 141(2–3):90–102. https://doi.org/10.1159/000354832
Kidwell MG. Transposable elements and the evolution of genome size in eukaryotes. Genetica. 2002; 115(1):49–63. https://doi.org/10.1023/A:1016072014259
Kubat Z, Hobza R, Vyskot B, Kejnovsky E. Microsatellite accumulation in the Y chromosome of Silene latifolia. Genome. 2008; 51(5):350–56. https://doi.org/10.1139/G08-024
Levan A, Fredga K, Sandberg AA. Nomenclature for centromeric position on chromosomes. Hereditas. 1964; 52(2):201–20. https://doi.org/10.1111/j.1601-5223.1964.tb01953.x
Matsuoka MP, Gharrett AJ, Wilmot RL, Smoker WW. Genetic linkage mapping of allozyme loci in even- and odd-year pink salmon (Oncorhynchus gorbuscha). J Hered. 2004; 95(5):421–29. https://doi.org/10.1093/jhered/esh069
Melo BF, Sidlauskas BL, Near TJ, Roxo FF, Ghezelayagh A, Ochoa LE et al. Accelerated diversification explains the exceptional species richness of tropical characoid fishes. Syst Biol. 2022; 71(1):78–92. https://doi.org/10.1093/sysbio/syab040
de Moraes RLR, Bertollo LAC, Marinho MMF, Yano CF, Hatanaka T, Barby FF et al. Evolutionary relationships and cytotaxonomy considerations in the genus Pyrrhulina (Characiformes, Lebiasinidae). Zebrafish. 2017; 14(6):536–46. https://doi.org/10.1089/zeb.2017.1465
de Moraes RLR, Sassi FMC, Bertollo LAC, Marinho MMF, Viana PF, Feldberg E et al. Tracking the evolutionary trends among small-size fishes of the genus Pyrrhulina (Characiforme, Lebiasinidae): New insights from a molecular cytogenetic perspective. Front Genet. 2021; 12:769984. https://doi.org/10.3389/fgene.2021.769984
de Moraes RLR, Sember A, Bertollo LAC, de Oliveira EA, Ráb P, Hatanaka T et al. Comparative cytogenetics and neo-Y formation in small-sized fish species of the genus Pyrrhulina (Characiformes, Lebiasinidae). Front Genet. 2019; 10:678. https://doi.org/10.3389/fgene.2019.00678
Netto-Ferreira AL. Revisão taxonômica e relações interespecíficas de Lebiasininae (Ostariophysi: Characiformes: Lebiasinidae). [PhD Thesis]. São Paulo: Universidade de São Paulo; 2010. https://doi.org/10.11606/T.41.2010.tde-02022011-165808
Netto-Ferreira AL. Three new species of Lebiasina (Characiformes: Lebiasinidae) from the Brazilian shield border at Serra do Cachimbo, Pará, Brazil. Neotrop Ichthyol. 2012; 10(3):487–98. https://doi.org/10.1590/S1679-62252012000300002
Netto-Ferreira AL, Marinho MMF. New species of Pyrrhulina (Ostariophysi: Characiformes: Lebiasinidae) from the Brazilian Shield, with comments on a putative monophyletic group of species in the genus. Zootaxa. 2013; 3664(3):369–76. https://doi.org/10.11646/zootaxa.3664.3.7
Netto-Ferreira AL, Oyakawa OT, Zuanon J, Nolasco JC. Lebiasina yepezi, a new Lebiasininae (Characiformes: Lebiasinidae) from the Serra Parima-Tapirapecó mountains. Neotrop Ichthyol. 2011; 9(4):767–75. https://doi.org/10.1590/S1679-62252011000400008
Oliveira C, Avelino GS, Abe KT, Mariguela TC, Benine RC, Ortí G et al. Phylogenetic relationships within the speciose family Characidae (Teleostei: Ostariophysi: Characiformes) based on multilocus analysis and extensive ingroup sampling. BMC Evol Biol. 2011; 11(1):275. https://doi.org/10.1186/1471-2148-11-275
Ortí G, Meyer A. The radiation of characiform fishes and the limits of resolution of mitochondrial ribosomal DNA sequences. Syst Biol. 1997; 46(1):75–100. https://doi.org/10.1093/sysbio/46.1.75
Oyakawa OT. Relações filogenéticas das famílias Pyrrhulinidae, Lebiasinidae e Erythrinidae (Osteichthyes: Characiformes). [PhD Thesis]. São Paulo: Universidade de São Paulo; 1998. Available from: https://repositorio.usp.br/item/000952251
Pendás AM, Moran P, Freije JP, Garcia-Vazquez E. Chromosomal mapping and nucleotide sequence of two tandem repeats of Atlantic salmon 5S rDNA. Cytogenet Genome Res. 1994; 67(1):31–36. https://doi.org/10.1159/000133792
Sambrook J, Russell DW. Molecular Cloning: A laboratory manual. Vol. 1, 2, 3. 3rd ed. New York: Cold Springs Harbor Laboratory Press; 2001.
Sassi FMC, Hatanaka T, de Moraes RLR, Toma GA, Oliveira EA, Liehr T et al. An insight into the chromosomal evolution of Lebiasinidae (Teleostei, Characiformes). Genes (Basel). 2020; 11(4):365. https://doi.org/10.3390/genes11040365
Sassi FMC, Oliveira EA, Bertollo LAC, Nirchio M, Hatanaka T, Marinho MMF et al. Chromosomal evolution and evolutionary relationships of Lebiasina species (Characiformes, Lebiasinidae). Int J Mol Sci. 2019; 20(12):2944. https://doi.org/10.3390/ijms20122944
Seehausen O, Wagner CE. Speciation in freshwater fishes. Annu Rev Ecol Evol Syst. 2014; 45:621–51. https://doi.org/10.1146/annurev-ecolsys-120213-091818
Sember A, Oliveira EA, Ráb P, Bertollo LAC, de Freitas NL, Viana PF et al. Centric fusions behind the karyotype evolution of neotropical Nannostomus pencilfishes (Characiformes, Lebiasinidae): First insights from a molecular cytogenetic perspective. Genes (Basel). 2020; 11(1):91. https://doi.org/10.3390/genes11010091
Sochorová J, Garcia S, Gálvez F, Symonová R, Kovaík A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma. 2018; 127(1):141–50. https://doi.org/10.1007/s00412-017-0651-8
Souza J, Guimarães E, Pinheiro-Figliuolo V, Cioffi MB, Bertollo LAC, Feldberg E. Chromosomal analysis of Ctenolucius hujeta Valenciennes, 1850 (Characiformes): A new piece in the chromosomal evolution of the Ctenoluciidae. Cytogenet Genome Res. 2021; 161(3–4):195–202. https://doi.org/10.1159/000515456
de Souza e Sousa JF, Viana PF, Bertollo LAC, Cioffi MB, Feldberg E. Evolutionary relationships among Boulengerella species (Ctenoluciidae, Characiformes): genomic organization of repetitive DNAs and highly conserved karyotypes. Cytogenet Genome Res. 2017; 152(4):194–203. https://doi.org/10.1159/000480141
Steindachner F. Beiträge zuer Kenntniss der Characinen des Amazonenstromes. Wien: Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Classe; 1876.
Sumner AT. A simple technique for demonstrating centromeric heterochromatin. Exp Cell Res. 1972; 75(1):304–06. https://doi.org/10.1016/0014-4827(72)90558-7
Tchurikov NA, Uroshlev LA, Klushevskaya ES, Alembekov IR, Lagarkova MA, Kravatskaya GI et al. Chromosomal translocations in NK-cell lymphomas originate from inter-chromosomal contacts of active rDNA clusters possessing hot spots of DSBs. Cancers (Basel). 2021; 13(15):3889. https://doi.org/10.3390/cancers13153889
Toma GA, de Moraes RLR, Sassi FMC, Bertollo LAC, de Oliveira EA, Rab P et al. Cytogenetics of the small-sized fish, Copeina guttata (Characiformes, Lebiasinidae): Novel insights into the karyotype differentiation of the family. PLoS ONE. 2019; 14(12):e0226746. https://doi.org/10.1371/journal.pone.0226746
Weitzman M, Weitzman SH. Family Lebiasinidae. In: Reis RE, Kullander SO, Ferraris CJ, Jr., organizers. Check list of the freshwater fishes of South and Central America. Porto Alegre: EDIPUCRS; 2003.
Yano CF, Bertollo LAC, Cioffi MB. Fish-FISH: Molecular cytogenetics in fish species. In: Liehr T, editor. Fluorescence in situ hybridization (FISH): Application Guide. 2nd ed. Berlin, Heildelberg: Springer Protocols Handbooks; 2017. p.429–43. https://doi.org/10.1007/978-3-662-52959-1_44
Zwick MS, Hanson RE, Islam-Faridi MN, Stelly DM, Wing RA, Price HJ et al. A rapid procedure for the isolation of C0t-1 DNA from plants. Genome. 1997; 40(1):138–42. https://doi.org/10.1139/g97-020
Authors

![]() Priscila Polaquini de Macedo Leite1
Priscila Polaquini de Macedo Leite1 ![]() ,
, ![]() Francisco de Menezes Cavalcante Sassi1,
Francisco de Menezes Cavalcante Sassi1, ![]() Manoela Maria Ferreira Marinho2,
Manoela Maria Ferreira Marinho2, ![]() Mauro Nirchio3,
Mauro Nirchio3, ![]() Renata Luiza Rosa de Moraes,
Renata Luiza Rosa de Moraes, ![]() Gustavo Akira Toma1,
Gustavo Akira Toma1, ![]() Luiz Antonio Carlos Bertollo1 and
Luiz Antonio Carlos Bertollo1 and ![]() Marcelo de Bello Cioffi1
Marcelo de Bello Cioffi1
[1] Laboratório de Citogenética de Peixes, Departamento de Genética e Evolução, Universidade Federal de São Carlos (UFSCar), Rod. Washington Luiz, s/n, 13565-905 São Carlos, SP, Brazil. (PPML) priscila_pm@live.com (corresponding author), (FMCS) fmcsassi@estudante.ufscar.br, (RLRM) rlrdm@hotmail.com, (GAT) gustavo_toma@hotmail.com, (LACB) bertollo@ufscar.br, (MBC) mbcioffi@ufscar.br.
[2] Departamento de Sistemática e Ecologia, Universidade Federal da Paraíba, 58051900 João Pessoa, PB, Brazil. manumfm@yahoo.com.br.
[3] Facultad de Ciencias Agropecuarias, Universidad Técnica de Machala, Av. Panamericana, km 5.5, Vía a Pasaje, 070151 Machala, Ecuador. mauro.nirchio@gmail.com.
Authors’ Contribution 

Priscila Polaquini de Macedo Leite: Formal analysis, Methodology, Validation, Visualization, Writing-original draft.
Francisco de Menezes Cavalcante Sassi: Data curation, Formal analysis, Investigation, Methodology, Software, Writing-review and editing.
Manoela Maria Ferreira Marinho: Formal analysis, Investigation, Validation, Writing-review and editing.
Mauro Nirchio: Formal analysis, Validation, Writing-review and editing.
Renata Luiza Rosa de Moraes: Investigation, Methodology, Writing-review and editing.
Gustavo Akira Toma: Formal analysis, Methodology, Visualization, Writing-review and editing.
Luiz Antonio Carlos Bertollo: Investigation, Methodology, Validation, Visualization, Writing-review and editing.
Ethical Statement
Samples were collected under licenses 48628-2 and A96FF09 issued by the Brazilian bureaus of environmental control ICMBio/SISBio and SISGEN, respectively. All procedures followed ethical, and anesthesia conducts according to the Committee of Ethics in Animal Use and Experimentation of the Universidade Federal de São Carlos (Process number CEUA 1853260315).
Competing Interests
The authors declare no competing interests.
How to cite this article
Leite PPM, Sassi FMC, Marinho MMF, Nirchio M, Moraes RLR, Toma GA, Bertollo LAC, Cioffi MB. Tracking the evolutionary pathways among Brazilian Lebiasina species (Teleostei: Lebiasinidae): a chromosomal and genomic comparative investigation. Neotrop Ichthyol. 2022; 20(1):e210153. https://doi.org/10.1590/1982-0224-2021-0153
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2022 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted January 11, 2022 by Claudio Oliveira
Accepted January 11, 2022 by Claudio Oliveira
![]() Submitted October 22, 2021
Submitted October 22, 2021
![]() Epub March 21, 2022
Epub March 21, 2022