![]() Emanuel B. Neuhaus1
Emanuel B. Neuhaus1 ![]() ,
, ![]() Marcelo R. Britto1,
Marcelo R. Britto1, ![]() José Luís O. Birindelli2 and
José Luís O. Birindelli2 and ![]() Leandro M. Sousa3
Leandro M. Sousa3
PDF: EN XML: EN | Cite this article
Abstract
A new Ancistrus species is described from Tapajós and Xingu river basins. It is distinguished from its congeners by the singular body color pattern, consisting of dark vermiculated stripes almost all over the body, and also by combination of features as a narrow head, large internostril distance, and absence of rows of enlarged odontodes on the lateral plates. In addition, the new species is distinguished from congeners that inhabit the rio Tapajós basin by the presence of a fully-developed adipose fin (vs. adipose fin absent in Ancistrus parecis and A. tombador, and vestigial adipose fin or absent in A. krenakarore). It differs from A. ranunculus, also from the rio Xingu, by the color pattern, smaller body size, smaller gill opening, and narrower cleithral width. The new taxon adds a new record to the list of species shared among the Xingu and Tapajós basins.
Keywords: Amazon, Brazilian Shield, Bristlenose pleco, Hypostominae, Taxonomy.
Uma nova espécie de Ancistrus é descrita das bacias dos rios Tapajós e Xingu. Distingue-se de suas congêneres pelo padrão de colorido único, que consiste em listras vermiculadas escuras em quase todo o corpo, e também pela combinação de características como cabeça estreita, maior distância entre as narinas e ausência de séries de grandes odontódeos nas placas laterais. Adicionalmente, a nova espécie pode ser diferenciada de suas congêneres da bacia do rio Tapajós pela presença de uma nadadeira adiposa totalmente desenvolvida (vs. nadadeira adiposa ausente em Ancistrus parecis e A. tombador,nadadeira adiposa vestigial ou ausente em A. krenakarore). Ela difere de A. ranunculus, a única outra espécie do rio Xingu, pelo padrão de colorido, menor tamanho corporal, menor abertura branquial e menor largura entre os cleitros. O novo táxon adiciona um novo registro à lista de espécies compartilhadas entre as bacias do Xingu e Tapajós.
Palavras-chave: Amazônia, Cascudo, Escudo Brasileiro, Hypostominae, Taxonomia.
Introduction
The genus Ancistrus Kner, 1854 includes 75 valid species (Fricke et al., 2021) distributed in South and Central Americas from Panama to Argentina, occurring in cis- and trans-Andean rivers. Among Loricariidae, the diversity of Ancistrus is only smaller than that of Hypostomus Lacepède, 1803. Ancistrus is easily distinguished from other loricariids by having naked snout margins (i.e., without plated snout, Figs. 1, 2) bearing fleshy expansions (tentacles). The genus Chaetostoma Tschudi, 1846 also has unplated snout, but differs from Ancistrus by the absence of fleshy tentacles and by having five plate rows at the narrowest part of the caudal peduncle (vs. three plate rows in Ancistrus). As for other highly diverse genera of Neotropical fish, the diversity of Ancistrus remains underestimated. In addition, more than half of valid species were described before 1950 in poorly-informative descriptions that leave doubt of the species delimitations. Consequently, there are a large number of possibly undescribed species, several of them highly appreciated in the ornamental fish trade. To overcome the taxonomic issues, aquarium fish hobbyists apply an alphanumeric code (the L-number system) to provisionally name the putative new species and morphotypes that appear in the hobby (Stawikowski, 1988).
Recent ichthyological surveys in the Brazilian Shield Amazon rivers captured several specimens of Ancistrus with a distinctive color pattern in the Tapajós and Xingu river basins. The specimens from the Xingu drainage are already known by aquarists by the code L159 (Stawikowski, 1994; Dignall, 2014). The present contribution aims to describe the new species, illustrating and diagnosing it, with comparisons to congeners.
Material and methods
Measurements were obtained with digital calipers to the nearest 0.1 mm. Measurements and counts were made only in the left side of the body, except when not possible, excluding the infraorbitals that were counted in both sides. Most morphometric data were taken following Fisch-Muller et al. (2001), and Armbruster (2003), in addition to nares-eye distance taken from posterior border of left naris to anterior border of left eye. Morphometrics are reported as percentages of standard length (SL), except for subunits of head which are reported as percentages of head length (HL). Plate terminology follows Schaefer (1997) and osteological nomenclature is according to Schaefer (1987), except for prefrontal plate used to name the plate which forms the posterolateral nostril margin and anterodorsal orbit margin (Schaefer, 1997), and compound pterotic used instead of pterotic-supracleithrum (Aquino, Schaefer, 2002). Terminology for snout areas of tentacles follows Sabaj et al. (1999). Some specimens were cleared and stained (cs) according to the protocol of Taylor, Van Dyke (1985). Meristic data follows Armbruster (2003) with addition of: complete series of mid-dorsal and mid-ventral plates; plates along dorsal-fin base: dorsal plates between dorsal-fin spine and the insertion of the last dorsal-fin branched ray; plates between end of the dorsal fin and adipose-fin origin: dorsal plates posterior to the insertion of the last dorsal-fin branched ray and adipose-spine origin; plates between adipose and caudal fins: dorsal plates between end of adipose-fin membrane and caudal fin; preanal plates: ventral plates before unbranched ray of anal fin; ventral plates between anal-fin base and caudal fin; vertebral count including those of the Weberian-complex and the counting the PU1+U2 as a single element; number of ribs; number of left and right infraorbitals; number of pterygiophores of the dorsal fin.
In the description, numbers between brackets represent the total number of specimens with those counts, whereas asterisks indicate counts of the holotype. Comparative data of Ancistrus caucanus Fowler, 1943, A. dolichopterus Kner, 1854, A. dubius Eigenmann & Eigenmann, 1889, A. eustictus (Fowler, 1945), A. hoplogenys (Günther, 1864), A. latifrons (Günther, 1869), and A. malacops (Cope, 1872) were obtained through their original descriptions and photographs of type-specimens available from Morris et al. (2006). Institutional abbreviations follow Sabaj (2020). Due to the wide distribution of the new species and consequent variation of diagnostic features, we restricted the type-series to specimens from the Teles Pires basin. Therefore, the non-type series embraces non-measured specimens, specimens in no good condition for reliable identification, live specimens and specimens from the Xingu basin.
Results
Ancistrus luzia, new species
urn:lsid:zoobank.org:act:31D5520F-B248-4226-8413-896A0535A341
(Figs. 1–3; Tab. 1)
Ancistrus L 159. ―Stawikowski, 1994:145 [DATZ magazine, new imports from Brazil, figs. 6 and 7].
Ancistrus sp. “lineolatus”. ―Ohara et al., 2017:259 [identification guidebook from Teles Pires].
Holotype. MNRJ 51458, male, 75.9 mm SL, Brazil, Mato Grosso, Guarantã do Norte, rio Teles Pires basin, unknown stream next to bridge on road about 8 km from the road BR-163, 09°47’15”S 54°57’33”W, 1 Oct 2008, M. R. Britto, J. Gomes, F. R. Carvalho & L. Fries.

FIGURE 1| Ancistrus luzia, holotype, MNRJ 51458, male, 75.9 mm SL, Brazil, Mato Grosso, Guarantã do Norte, a tributary of rio Teles Pires, Tapajós basin. Black spots in the ventral view are parasites.
Paratypes. Brazil, Mato Grosso, rio Tapajós basin: MNRJ 35484, 11, 26.1–74.7 mm SL (10, 33.4–74.7 mm SL), collected with the holotype. MNRJ 24622, 9, 1 cs, 33.2–62.9 mm SL, Terra Nova do Norte, riacho Doce, tributary of rio Bastião, rio Teles Pires basin, 10°33’00”S 55°4’25”W, 23 Jan 2002, P. Buckup, A. Aranda, F. Silva & C. Figueiredo. MNRJ 24641, 3, 47.8–52.2 mm SL, Peixoto de Azevedo, tributary of rio Peixoto de Azevedo at road BR-080, rio Teles Pires basin, 10°25’57”S 54°10’59”W, 24 Jan 2002, P. Buckup, A. Aranda, F. Silva & C. Figueiredo. MNRJ 24692, 2, 58.6–71.2 mm SL, Carlinda, tributary of rio Teles Pires, left margin, at road MT-208, 09°59’04”S 55°46’03”W, 23 Jan 2002, P. Buckup, A. Aranda, F. Silva & C. Figueiredo. MNRJ 35486, 3, 1 cs, 56.2–60.9 mm SL, Guarantã do Norte, tributary of rio Braço do Norte, Teles Pires basin, 09°56’55”S 55°01’51”W, 2 Oct 2008, P. Buckup, J. Maldonado, J. Birindelli, C. Chamon & W. Felzmann. MNRJ 35487, 11, 25.6–34.9 mm SL (1, 34.9 mm SL), Novo Mundo, stream and wetland at the road between Novo Mundo and Alta Floresta, 10°03’37”S 55°26’54”W, 2 Oct 2008, M. Britto, J. Gomes, F. Carvalho, L. Fries, F. Jerep & C. Zawadzki. MNRJ 35489, 2, 52.0–77.1 mm SL, Alta Floresta, corredeira Boca Quente, rio Teles Pires, 09°38’42”S 55°59’23”W, 3 Oct 2008, M. Britto, J. Gomes, C. Chamon, L. Fries, F. Jerep & C. Zawadzki. MZUEL 20724, 2, 40.2–54.6 mm SL, collected with the holotype. MZUEL 20725, 1, 69.0 mm SL, Alta Floresta, corredeira Boca Quente, rio Teles Pires, 09°38’42”S 55°59’23”W, 3 Oct 2008, M. Britto, J. Gomes, C. Chamon, L. Fries, F. Jerep & C. Zawadzki. MZUEL 20726, 1, 64.3 mm SL, Carlinda, tributary of rio Teles Pires, left margin, at road MT-208, 09°59’04”S 55°46’03”W, 23 Jan 2002, P. Buckup, A. Aranda, F. Silva & C. Figueiredo. MZUSP 62659, 8 (4, 42.6–54.8 mm SL), Claudia, córrego Loanda, at road BR-163, rio Teles Pires, 11°25’00”S 55°21’00”W, 21 Feb 1998, M. I. Landim & C. Moreira. MZUSP 95904, 6 (3, 61.0–82.9 mm SL), Itauba, rio Teles Pires, 11°03’44”S 55°19’08”W, 26 Sep 2007, J. Birindelli & P. H. Carvalho. MZUSP 96150, 4 (2, 54.9–55.1 mm SL), Paranaíta, rio Teles Pires, 09°26’58”S 56°29’19”W, 28 Sep 2007, L. Sousa & A. Netto-Ferreira. MZUSP 96820, 46, 1 cs (5, 45.7–79.5 mm SL), Peixoto de Azevedo, Cachoeira da Neblina, in a tributary of rio Peixoto de Azevedo, rio Teles Pires, 18 Oct 2007, J. Birindelli, L. Sousa, A. Netto-Ferreira, M. Sabaj-Perez & N. Lujan. Pará, rio Tapajós basin: MZUSP 96834, 4 (2, 22.1–29.1 mm SL), Novo Progresso, tributary of rio Braço do Norte, rio Peixoto de Azevedo, rio Teles Pires, at bridge of the road BR-163, 09°25’55”S 54°52’11”W, 19 Oct 2007, J. Birindelli, L. Sousa, A. Netto-Ferreira, M. Sabaj-Perez & N. Lujan. MZUSP 99877, 28 (18, 29.6–59.8 mm SL), Jacareacanga, rio Teles Pires, downstream Sete Quedas, 09°18’42”S 56°46’47”W, 9 Jun 2008, L. Sousa & A. Netto-Ferreira. MZUSP 99931, 4 (2, 58.7–61.1 mm SL), Jacareacanga, tributary of the left margin of rio Teles Pires, downstream of Sete Quedas, 09°18’27”S 56°47’38”W, 9 Jun 2008, L. Sousa & A. Netto-Ferreira.
Non-types. Brazil, Mato Grosso, rio Tapajós basin: MNRJ 35490, 1, 40.4 mm SL,Alta Floresta, cachoeira do Limão, at rio Cristalino, tributary right margin of rio Teles Pires, 09°32’17”S 55°54’14”W, 4 Oct 2008, P. Buckup, F. Carvalho, J. Birindelli, J. Gomes, F. Becker, R. Silvano & W. Felzmann. MNRJ 35491, 3, 53.6–61.8 mm SL, Alta Floresta, igarapé Corgão, and adjacent margin of rio Peixoto de Azevedo, Teles Pires basin, 09°43’37”S 55°50’06”W, 4 Oct 2008, P. Buckup, F. Carvalho, J. Birindelli, J. Gomes, F. Becker, R. Silvano & W. Felzmann. MNRJ 35492, 1, 32.2 mm SL, Alta Floresta, stream in front of Ilha do Refúgio, rio Teles Pires, 09°38’40”S 56°01’58”W, 4 Oct 2008, M. Britto, J. Gomes, C. Chamon, J. Maldonado, F. Jerep & C. Zawadzki. MNRJ 35493, 1, 32.7 mm SL, Alta Floresta, Ilha do Domingão, rio Teles Pires, 09°38’13”S 55°58’55”W, 4 Oct 2008, M. Britto, J. Gomes, C. Chamon, J. Maldonado, F. Jerep & C. Zawadzki. MZUSP 95639, 1, 36.7 mm SL, Paranaíta, rio Teles Pires, near the ferry of the road MT-426, 09°27’07”S 56°30’46”W, 27 Sep 2007, L. Sousa & A. Netto-Ferreira. MZUSP 96564, 1, Itaúba, stream at road BR-163, tributary of rio Teles Pires, 11°06’51”S 55°18’25”W, J. Birindelli & P. H. Carvalho. Pará, rio Xingu basin: LIA 3996, 2, 39.0–42.0 mm SL, igarapé at Morro do Félix, RESEX do Rio Xingu, Altamira, 05°22’18”S 52°55’37”W, 17 Set 2016, A. Gonçalves & M. D. Silva. LIA 7657, 4, 25.1–66.1 mm SL, igarapé without name, 03°28’39”S 52°32’36”W, 15 Set 2012, D. Bastos. MNRJ 35483, 1, 34.1 mm SL, stream at road BR-163, about 14 km north of Castelo dos Sonhos, 30 Sep 2008, M. Britto, J. Gomes, F. Carvalho & L. Fries. MNRJ 35503, 2, 36.1–42.9 mm SL, Altamira, rio Curuá, rio Iriri basin, 08°06’35”S 55°00’58”W, 30 Set 2008, P. Buckup, J. Maldonado, C. Zawadski, C. Chamon & F. Jerep. MNRJ 35504, 6, 22.5–48.5 mm SL (4, 37.5–48.5 mm SL), Altamira, rio Curuá, rio Iriri basin, small stream upstream the first waterfall, 08°44’02”S 54°57’45”W, 1 Oct 2008, M. Britto, J. Gomes, F. Carvalho & L. Fries. MNRJ 35505, 8, 23.4–71.6 mm SL (5, 36.9–71.6 mm SL), Altamira, rio Curuá, rio Iriri basin, waterfall of rio Curuá, 54 km from Castelo dos Sonhos, 08°44’06”S 54°57’43”W, 1 Oct 2008, M. Britto, J. Gomes, F. Carvalho & L. Fries. MNRJ 35506, 2, 61.3–80.7 mm SL, rio Curuá, rio Iriri basin, waterfall of rio Curuá, upstream the first waterfall, 08°44’10”S 54°57’47”W, 1 Oct 2008, M. Britto, J. Gomes, F. Carvalho & L. Fries. MZUEL 20727, 1, 62.5 mm SL, rio Curuá, rio Iriri basin, waterfall of rio Curua, upstream the first waterfall, 08°44’10”S 54°57’47”W, 1 Oct 2008, M. Britto, J. Gomes, F. Carvalho & L. Fries.
TABLE 1 | Morphometric data of Ancistrus luzia. N = number of specimens; SD = standard deviation.
Diagnosis. Ancistrus luzia is distinguished from its congeners, except A. claro Knaack, 1999 and A. centrolepis Regan, 1913, by the singular body color pattern, consisted of dark-vermiculated stripes on head, dorsal and lateral plates of trunk (vs. body uniformly colored or with light or dark spots or blotches over head or dorsal and lateral plates of trunk). Ancistrus luzia is distinguished from A. claro by its narrower head (head width 30.8–35.1% vs. 35.2–38.2% HL) and greater internostril distance (18.7–23.7% vs. 14.9–17.0% HL), and from A. centrolepis by the absence of rows of greatly enlarged odontodes on the lateral plates (vs. presence of one to three rows of enlarged odontodes on the lateral plates). Juveniles of A. leucostictus (Günther, 1864) might show lines similar to A. luzia, but color changes to light, large spots as specimens grow. In addition, the new species is distinguished from its congeners that inhabit the Tapajós basin by the presence of adipose fin fully developed (vs. adipose fin absent in A. parecis Fisch-Muller, Cardoso, da Silva & Bertaco, 2005, A. tombador Fisch-Muller, Cardoso, da Silva & Bertaco, 2005, and vestigial adipose fin or absent in A. krenakarore de Oliveira, Rapp Py-Daniel & Zuanon, 2016). Ancistrus luzia differs from A. ranunculus Muller, Rapp Py-Daniel & Zuanon, 1994, the only described species from the rio Xingu, by the smaller body size in adults, smaller gill opening (HL/interbranchial distance equals to 1.7 to 2.0 vs. 2.3 to 3.9) and narrower cleithral width (SL/cleithral width equals to 2.8–3.2 vs. 2.1–2.9).
Description. Morphometrics data in Tab. 1. Dorsal profile convex, ascending from the tip of snout until posterior limit of supraoccipital; almost straight to slightly convex between supraoccipital and dorsal-fin origin, descending from this point until posterior margin of adipose-fin base; slightly concave between end of adipose and first procurrent caudal-fin ray. Body width greatest at opercular region, progressively narrowing until end of caudal peduncle. Greatest body height at dorsal-fin origin and lowest at the caudal peduncle, near adipose-fin terminus. Body in cross section horizontally elliptical at abdominal region and vertically oval at caudal peduncle. Ventral profile gently convex between tip of snout and pelvic-fin origin, and then straight between pelvic fin and first procurrent caudal-fin ray.
Body covered with bony plates, except along dorsal-fin base, and ventral region between tip of snout and anal-fin origin. Plates arranged in four series from head to pelvic-fin base, with ventral series absent. Plates arranged in five series in trunk from pelvic-fin base to vertical through adipose-fin base with mid-dorsal and mid-ventral series ending at the latter point. Plates arranged in three series at the narrowest part of the caudal peduncle. Small sets of odontodes eventually occurring over the ventral skin close to pectoral-fin insertion. Lateral line visible from posterior margin of compound pterotic to posterior end of caudal peduncle, always following median plate series.
Body plates and fin rays covered with odontodes. Odontodes in regular rows on the plates, larger on posterior margin. Some individuals have odontodes just before insertion of unbranched anal-fin ray, arising from expanded transverse process of first pterygiophore of anal fin. Odontodes larger on distal half of pectoral-fin spine and partially covered with dermic membrane. Odontodes on pelvic-fin unbranched rays larger at ventral portion. Median plates 23(9), 24*(53), 25(2), 26(1); mid-dorsal plates 10(2), 12(1), 13(4), 14(6), 15(16), 16*(19), 17(15), 18(1), 19(1); mid-ventral plates 18(3), 19(38), 20*(17), 21(6), 22(1); plates along dorsal-fin base 5(3), 6*(61), 7(1); plates between end of dorsal and adipose fins 6(1), 7*(46), 8(17), 9(1); plates between adipose and caudal fins, 2(41), 3*(24); preanal plates 2(1), 3(3), 4*(59), 5(2); ventral plates between anal-fin base and caudal fin 12(4), 13*(54), 14(5), 15(1). Urogenital pore covered with tubular-shape urogenital papilla.
Rounded snout, in dorsal view, with naked margin. See description on Sexual Dimorphism for details on snout and tentacles. Presence of subdermal platelets bordering the naked margin and/or in the base of tentacles (Fig. 2). Oral disc rounded, with small papillae on its inner surface decreasing in size toward external margin. Lower lip larger than upper, not reaching pectoral girdle. Barbel short, length nearly half of orbit diameter and free from lower lip in most specimens. Bicuspid teeth with elongate mesial cusp and short lateral cusp, cusps separated from each other by V-shaped notch; 49–76 (mean 62) teeth in premaxilla, 48–75 (mean 62) dentary. Post-opercular region naked, with zero to eight dermal platelets normally grouped near anterior margin of compound pterotic; 6–14 (mean 11) evertible cheek odontodes distally hooked, hyaline at base and yellowish amber at tip.

FIGURE 2| Ancistrus luzia, paratype, MNRJ 24622, cs, female, 48.9 mm SL, showing the subdermal platelets bordering the naked margin of snout. Arrows indicate platelets.
Dorsal-fin origin slightly anterior to pelvic-fin origin; dorsal fin II,6(1), II,7*(63) or II,8(1). First unbranched ray (spinelet) short and V-shaped (one specimen with spinelet atrophied), second unbranched ray (dorsal spine) well developed. Last branched-rays reaching or almost reaching pre-adipose plate, when adpressed. Pectoral fin I,6*(65), when adpressed reaching pelvic-fin insertion in small individual (30 mm) or reaching half-length of pelvic unbranched rays in larger specimens. Pelvic fin i,5*(65), with posterior margin reaching approximately half of anal-fin rays. Anal fin i,3(1) or i,4*(63). Caudal-fin truncate; i,14,i*(65); with upper lobe shorter than lower; dorsal and ventral procurrent caudal-fin rays 5*(28) or 6(16) and 3(8), 4*(31), or 5(1), respectively.
Eyes rounded, located dorsolaterally on head. Orbit delimited dorsally by prefrontal plate, frontal and sphenotic, and ventrally by infraorbitals four to six. Anterior and posterior nares located dorsally on head, separated from each other by a flap of skin. Anterior naris with tubular expansion of skin. Posterior naris separated from orbit by prefrontal plate and frontal. Six infraorbitals in total.
Palatine splint present in one specimen and absent the other two cleared and stained. When present thin, elongate and ossified, located near palatine cartilage and lateral to palatine. Four branchiostegal rays, only first one associated with posterohyal; remaining three supported by branchiostegal membrane. Three basibranchial elements, only first one ossified. Five pairs of hypobranchial elements, only first ossified. Five pairs of ossified ceratobranchials; first with large accessory flange supporting first row of modified gill rakers; fifth expanded, with numerous small teeth on dorsomesial edge, and supporting row of modified gill rakers on dorsal surface. Four ossified epibranchials. Two infrapharyngobranchials: one associated with third epibranchial and the other with fourth. Upper pharyngeal tooth plates with teeth along entire ventral portion.
Vertebrae 27, with sixth vertebra and eighth to 14th(2) or 15th(1) vertebrae bearing ribs. First rib (associated with sixth centrum) conspicuously larger. Neural spine absent in vertebral centrum eight. Centra with bifid neural spines: 10–15(3). Bifid neural arches reaching dorsal dermal plates, except for ninth centrum, reduced. Hemal spines present on vertebrae 9–27(1) or 10–27(2) and reaching ventral dermic plates from 18th or 19th. Dorsal fin with eight pterygiophores, first two sutured to each other and to neural spine of seventh centrum, and in some specimens also to anterior portion of eighth centrum; transverse process present on first five proximal radials; last one with pair of elongate posterior processes. Branched rays of pectoral fin supported by three radials. Lateral and internal pairs of anterior processes of basipterygia reaching each other at central portion of pelvic girdle. First pterygiophore of anal fin contacting hemal spine of centrum 14.

FIGURE 3| Color pattern of juvenile specimens of Ancistrus luzia. A, B. MZUSP 99877, 33.2 mm SL, from rio Teles Pires basin. C, D. MNRJ 35504, 37.5 mm SL, from the rio Curuá, rio Xingu basin.
Color in alcohol. Ground color of head, dorsum and lateral portions of trunk brown, with striated dark lines. In snout, dark lines somewhat parallel to each other, straight or curved, eventually branched, and longitudinal to axis of body. Dark lines outline bones of neurocranium and predorsal plates posterior to orbit. Lips yellowish brown. Ventral region of abdomen brown, without lines or blotches (with exception of two specimens of MNRJ 24622 that showed rounded dark brown blotches). All fins with brown interradial membranes. Dorsal fin sometimes with a dark blotch at base of membrane, between dorsal-fin spine and first branched ray. Dorsal-fin rays with two to five blotches over rays usually expanded over membranes. Caudal fin with dark spots over rays and interradial membranes, forming vertical, straight to sinuous and discontinuous stripes. Vertical dark band close to caudal-fin base, posterior to platelets. Tips of caudal fin and adjacent branched rays yellowish white or hyaline, some individuals with tip of dorsal fin and adjacent branched rays hyaline. Color of pectoral and pelvic fins similar to dorsal fin. Juveniles have caudal fin with wide, vertical dark band; some specimens with tip of dorsal spine and adjacent branched rays, as well as tip of caudal-fin lobes red-brown colored (MZUSP 99877; Figs. 3A, B). In populations from rio Curuá, the juveniles have a similar color pattern described above (Figs. 3C, D). Nevertheless, in adults lateral and dorsal plates do not have vermiculation but an inconspicuous dark longitudinal band passing through median plates instead. Paired and dorsal fins without spots. Caudal fin brown with base and distal portions dark brown. All membrane fins are brown (Fig. 4A).
Color in life. Live specimens show similar color pattern observed in specimens preserved in alcohol, except for ground coloration more greenish, and dark lines more conspicuous (Figs. 4B, 5, 6).

FIGURE 4| Lateral view of Ancistrus luzia, MNRJ 35505, 71.6 mm SL, rio Curuá, Xingu basin. A. Preserved specimen. B. Specimen in life.

FIGURE 5| Ancistrus luzia in life, MZUSP 96564, rio Teles Pires, rio Tapajós basin.

FIGURE 6| Ancistrus luzia in life, specimen not preserved, 80.6 mm SL, rio Bacajaí, 03°35’13”S 51°46’00”W, tributary to middle rio Xingu.
Sexual dimorphism. Naked margin of snoutnarrower in females and juveniles (width smaller than distance between nares) and wide in adult males (width equal or greater than distance between nares). Naked margin in nuptial males reaching the anterior margin of nasal openings. Fleshy tentacles on naked region visible in individuals larger than 30 mm, well developed in nuptial males. Males with numerous and larger tentacles than females but number and size vary among specimens of same sex and similar standard length. Some adult females without tentacles (observed in MZUSP 96820). Tentacles along the snout margin smaller and, with a few exceptions, unbranched. Most tentacles branched in the medial row. Some mature females with yellowish-white dots around urogenital papilla (Fig. 7).
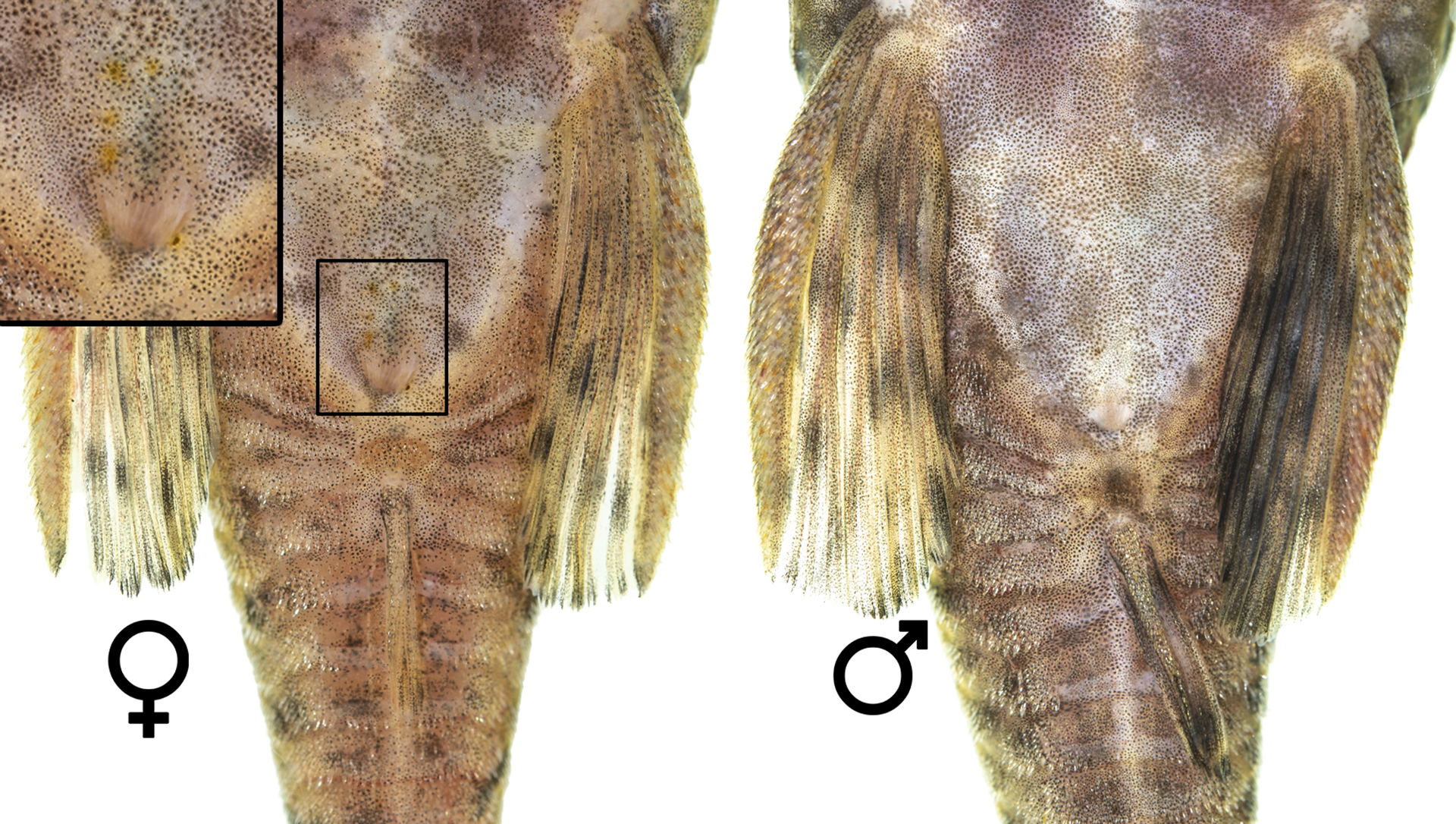
FIGURE 7| Ventral view of pelvic girdle of Ancistrus luzia showing differences in urogenital papillae of female (63.2 mm SL) and male (80.6 mm SL).
Etymology. Named after “Luzia” (Lapa Vermelha IV Hominid 1), a female Homo sapiens skeleton and one of the oldest human remains found in the Americas (11,000–11,500 years before present), in 1974 and 1975 during excavations coordinated by the French archaeologist Annette Lamin-Emperaire (1917–1977) from the site of Lapa Vermelha IV, Lagoa Santa region, municipality of Pedro Leopoldo, state of Minas Gerais, Brazil (Neves et al., 1999). The skeleton is deposited at Museu Nacional Biological Anthropology collection (catalogue number 01959), Universidade Federal do Rio de Janeiro, and was recovered again after the fire that hit the institution on September 2, 2018 by Museu Nacional Rescue team, becoming a symbol of institutional hope and resilience. A noun in apposition.
Geographical distribution. Ancistrus luzia is known from rio Teles Pires and its tributaries (rio Tapajós basin) and small to medium tributaries of middle and upper rio Xingu basin (Fig. 8) always associated with moderate water flow and rocky substrates (Fig. 9).
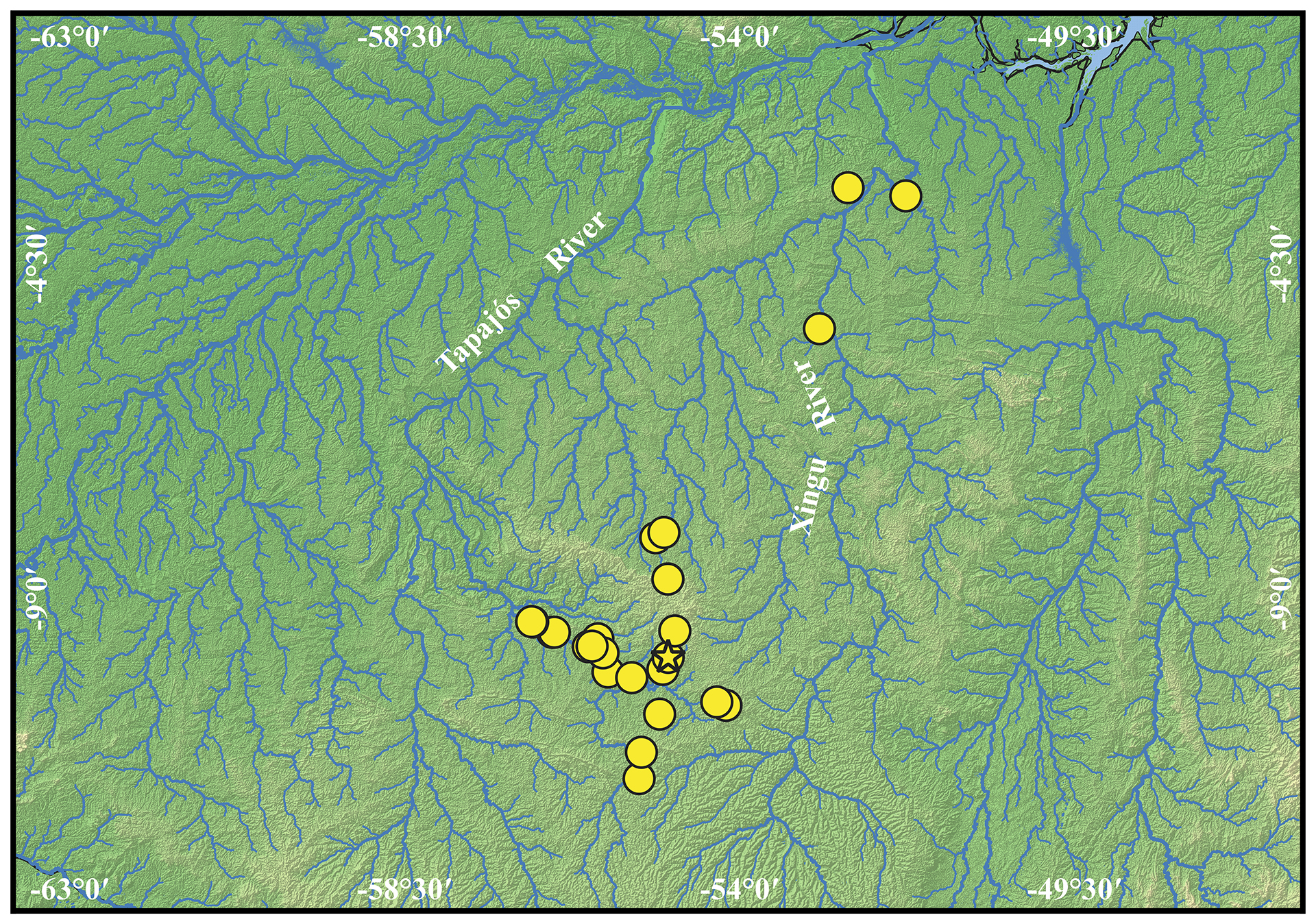
FIGURE 8| Geographic distribution of Ancistrus luzia. Star represents its type locality.

FIGURE 9| Habitats of Ancistrus luzia: (above) rio Teles Pires, Itauba, Mato Grosso (MZUSP 95904), photo by José L. O. Birindelli; (middle) rio Bacajaí (03°35’13”S 51°46’00”W ), tributary to middle rio Xingu, photo by Leandro Sousa; and (below) igarapé without name, Middle Xingu, Altamira, Pará (LIA 7657), photo by Douglas Bastos.
Conservation status. Ancistrus luzia is widely distributed across the Teles Pires basin and in different tributaries of the rio Xingu. A substantial number of cataloged specimens from distinct localities and distinct field expeditions over the past two decades suggest that the species is not threatened. In the rio Xingu, part of its tributaries at Volta Grande region were flooded by the Belo Monte Hydropower dam, but specimens were recently collected (non-cataloged material) in small streams not affected by the dam. Thus, A. luzia may be categorized as Least Concern (LC) according to the categories and criteria of the International Union for Conservation Nature (IUCN Standards and Petitions Subcommittee, 2019).
Discussion
Although there is a relatively large number of valid species allocated in Ancistrus, the diversity of the genus remains underestimated. To overcome the lack of scientific names, aquarium hobbyists developed an alphanumeric code system, the L-number system (Stawikowski, 1988), to address possibly undescribed species. In the present paper, one of these L-numbered species, the L159 (Stawikowski, 1994), is given a scientific name: Ancistrus luzia.
Ancistrus luzia is the second species described from the Xingu basin and the fourth from the Tapajós basin. Ancistrus ranunculus was described from the middle Xingu and Tocantins basins (Muller et al., 1994) and differs from A. luzia by the color pattern (dark body, without vermiculations vs. body brown with dark-line vermiculations), by the wider gill opening (HL/interbranchial distance equals 2.3 to 3.9 vs. 1.7 to 2.0 in A. luzia) and wider body (SL/cleithral width equals to 2.1–2.9 vs. 2.8–3.2). The three described species from the Tapajós are easily distinguished by having adipose fin reduced, or absent. Ancistrus parecis has its type locality in the rio Formiga, a tributary of rio Juruena (Fisch-Muller et al., 2005b); A. tombador was described from the igarapé Ribeirão Preto, a tributary of rio Arinos (Fisch-Muller et al., 2005a); and A. krenakarore has its type locality in the rio Itapacurá, a right bank tributary of rio Tapajós (de Oliveira et al., 2016). According to de Oliveira et al. (2016), the reduction of the adipose fin in these species probably evolved independently due to their distinct development. In A. krenakarore, the spine and a reduced membrane are present in juveniles, and they are gradually absorbed into the skin during ontogenetic development (in a few cases, a vestigial fin can persist in adults). On the other hand, in A. parecis and A. tombador, the adipose fin seems to be absent from early developmental stages. In this case, small-unpaired dorsal plates form a crest that replaces the fin.
There is no phylogenetic hypothesis including the species of Ancistrus treated herein. In addition, no other valid species has a similar color pattern as that seen in A. luzia. Nevertheless, the new species shares with congener’s small ossifications, not supporting odontodes at the bases of each of the large tentacles, a feature first described in Armbruster (2004). Moreover, some specimens of A. luzia bear weak subdermic platelets surrounding the naked margin of snout but not associated to tentacles (Fig. 2). The presence of these features could help to understand the evolution of the new species among congeners.
In addition to Ancistrus luzia, the Tapajós and the Xingu basins share several other species. Buckup et al. (2011) registered 155 species occurring in both basins. According to Dagosta, de Pinna (2017), tributaries of the upper Tapajós, like the Teles Pires, Juruena and Jamanxim rivers, are more related to Xingu and Tocantins rivers than to the lower portion of the Tapajós River. Due to the expansion of agricultural activities, the rio Teles Pires basin has been suffering environmental degradation. Large forest areas have been converted to cattle pasture or soybeans farming (Zaiatz et al., 2018). These activities occur mainly in the middle and high portions of the river basin, which is a transition zone between Amazonian and Cerrado domains (Ackerly et al., 1989). In addition, in the last recent decades, several portions of the river suffered with mining activities and hydroelectric power plants that are changing rapids (habitat of several catfish species) to lentic environments (Fernandes et al., 2011; Ohara et al., 2017; Lucanus et al., 2021).
We considered the conservation status of Ancistrus luzia as Least Concern (LC) based on the substantial number of catalogued specimens collected through the last two decades. Nevertheless, part of localities of A. luzia were converted from lotic to lentic environments due to construction of the hydropower plants São Manoel, Teles Pires, Colíder, and Sinop. They are part of the Amazon’s Tapajós Dam Complex, a plan to construct a total of 43 dams in the Tapajós basin (Fearnside, 2015). Other dams from this complex are predicted for the Teles Pires, which would enable the Tapajós-Teles Pires Water Way, planned to carry soybeans from Mato Grosso State to the Amazon River and the Atlantic Ocean (DNIT, 2018). This waterway would require additional interventions in rapids that impede navigation, eliminating A. luzia habitat in large stretches of the Teles Pires basin. Therefore, further surveys will show how the species will respond to all these environmental changes.
Comparative material examined. Ancistrus abilhoai Bifi, Pavanelli & Zawadzki, 2009. MZUSP 104116, holotype, 99.3 mm SL, Brazil, Paraná, Lapa, middle rio Iguaçu basin, rio Iguaçu, 25°47’35”S 50°11’59”W. Ancistrus agostinhoi Bifi, Pavanelli & Zawadzki, 2009. MZUSP 104119, paratypes, 2, 70.7–85.1 mm SL, Brazil, Paraná, Foz do Jordão, lower rio Iguaçu basin, rio Jordão, 25°39’12”S 51°58’15”W. Ancistrus aguaboensis Fisch-Muller, Mazzoni & Weber, 2001. MNRJ 20850, holotype, 66.3 mm SL, Brazil, Goiás, Minaçú, córrego Água Boa, small tributary of rio Bonito, a left bank tributary of the upper rio Tocantins, 10 km from Minaçú on road Minaçú-Campinaçu, 13º35’S 48º14’W, J. L. Costa Novaes, R. Mazzoni, F. Moraes & C. Weber, 1 Oct 1996. Ancistrus alga (Cope, 1872). LBP 22525, 1, rio Marañon, Amazonas, Peru, 04°19’45.9”S 70°00’30.2”W. C. Oliveira, B. F. Melo, J. M. Marin & G. Aricari, 13 Nov 2016. Ancistrus brevipinnis (Regan, 1904). MNRJ 42578, 27 (10, 41.6–71.47 mm SL), Brazil, Rio Grande do Sul, Caraá, Boco Stream, tributary of rio dos Sinos at Linha Padre Vieira II, Jacuí basin, 29º48’24”S 50º17’18”W, 8 Feb 2014, P. A. Buckup, R. S. Lima & G. V. Buckup. UFRGS 5672, 32 (8, 33.8–82.8 mm SL), Brazil, Rio Grande do Sul, Júlio de Castilhos, Felício stream, about 10 km SSE from Júlio de Castilhos, laguna dos Patos basin, 29°19’6.00”S 53°37’59.01”W, 28 Nov 2000, L. Malabarba, V. Bertaco, M. Azevedo, J. Bastos & C. Ricken. Ancistrus bolivianus (Steindachner, 1915), MHNG 2653.001, synonym of A. marcapatae (Regan, 1904). Bolivia, Tiraque, rivière Huayruru, Espiritu Santo, Chapare, Ichilo, Mamoré, Madeira basin, 17°28’00”S 65°32’60.0”W, F. M. Carvajal, 14 Jul 2004. Ancistrus bufonius (Valenciennes, 1840). ANSP 182999, 9, Peru, Apurimac, rio Apurimac (Ucayali basin), Cconoc, WSW of Limatambo near bridge crossing of main road to Abancay, 13°32’34”’S 72°38’19”’W, 13 Jul 2004, M. Sabaj et al. Ancistrus chagresi Eigenmann & Eigenmann, 1889. ROM 93181, 1, 109.9 mm SL, Panama, Colon Province, rio Caimito, 08°56’31.20”N 80°40’11.21”W, 20 Apr 2012, R. J. Eakins & J. C. Tetreault. Ancistrus centrolepis. BMNH 1910.7.11.122, paralectotype, female, 148.3 mm SL, Colombia, rio San Juan. Ancistrus cirrhosus (Valenciennes, 1836). CAS-SU 31597, 1, 96.7 mm SL, Argentina, Buenos Aires, La Plata basin. Ancistrus claro. MCP 28667, holotype, male, 64.5 mm SL, Brazil, Mato Grosso, rio Claro, above the bridge, in the direction of Chapada dos Guimaraes. MCP 28668, paratypes, 9 (3, 41.29–61.69 mm SL), same data as holotype. Ancistrus cryptophthalmus Reis, 1987. MZUSP 28809, holotype, 53.6 mm SL, Brazil, Goiás, São Domingos, Caverna Passa Três (about 300 m into cave), rio São Vicente System, 1980, Clube Alpino Paulista leg. Ancistrus cuiabae Knaack, 1999. MCP 28671, holotype, male, 108.5 mm SL, Brazil, Mato Grosso, Pantanal, 36 km southeast of Poconé, in a permanent remnant water of rio Bento Gomes, Paraguai basin. MCP 28672, paratypes, 9 (4, 54–82.1 mm SL), same data as holotype. Ancistrus falconensis Taphorn, Armbruster & Rodríguez-Olarte, 2010. ANSP 189316, paratypes, 2, 116.0–92.7 mm SL. Falcón, El Hueque River, 11º09’41.4”N 69º33’24.9”W, at bridge on highway from Churuguara to Coro, 19 Oct 2005. D. Taphorn, D. Rodríguez Olarte, J. Coronel, A. Amaro & H. Rivera. Ancistrus formoso Sabino & Trajano, 1997. MZUSP 51836, holotype, female, 71.3 mm SL, Brazil, Mato Grosso do Sul, Bonito, Buraco do Ducho cave, Serra da Bodoquena, 21º08’S 56º28’W. Ancistrus galani Pérez & Viloria, 1994. MCP 15634, paratype, female, 55.4 mm SL, Venezuela, Zulia, Sierra de Perijá, Río Socuy, Cueva de Los Laureles (Zu. 31), 10º45’04”N 72º27’42”W, 25 Mar 1991, A. Viloria, J. Lagarde. Ancistrus jataiensis Fisch-Muller, Cardoso, da Silva & Bertaco, 2005. MCP 35244, holotype, male, 52.7 mm SL, Brazil, Goiás, Mambaí, córrego Jataí, tributary of the rio Vermelho, 14°29’S 46°6’W, rio Tocantins basin, Centro de Biologia Aquática – UCG, 8 Sep 2002. Ancistrus karajas de Oliveira, Rapp Py-Daniel, Zawadzki & Zuanon, 2016. NUP 14587, paratype, 1, 34.7 mm SL, rio Parauapebas, Igarapé Alemão, Floresta Nacional de Carajás, 06°00’34”S 50°35’08”W, G. M. dos Santos, 17 Sep 2008. Ancistrus kellerae de Souza, Taphorn & Armbruster, 2019. AUM 62849, paratypes, 3 (1, 55.0 mm SL), Guyana, Region 8 (Potaro/Siparuni), Potaro River drainage, Kuribrong River, in rapids at Grass Shoals, 05°24’28”N 59°31’54”W, 12 Mar 2014 J. Armbruster, D. Werneke, E. Liverpool, D. Fernandes & D. Taphorn. Ancistrus krenakarore. NUP 14585, paratypes, 2, 36.1–38.9 mm SL, Brazil, Pará, Rurópolis, rio Itapacurá, right bank tributary of rio Tapajós, 04°36’20”S 55°30’41”W, F. Ribeiro & W. Pedroza, 9 Sep 2009. Ancistrus leoni de Souza, Taphorn & Armbruster, 2019. AUM 43665, paratypes, 5 (4, 44.9–49.3 mm SL), Casiquiare River, left bank upstream from mouth of Siapa River, 02°09’21”S 66°27’50”W, 19–22 Mar 2005, N. Lujan, M. Arce, T. Wesley & G. Santaella. Ancistrus leucostictus. BMNH 1864.1.21.85, 1, 48.6 mm SL, holotype of Chaetostomus leucostictus Günther, 1864. Essequibo River, Guyana. Ancistrus lithurgicus Eigenmann, 1912. BMNH 1911.10.31.107–108, 1, 35.1 mm SL, paratype, Essequibo River, Guyana. Ancistrus macrophthalmus AUM 42105, 5 (4, 74.8–59.9 mm SL), Venezuela, Rio Orinoco, 50 km of San Fernando de Atabapo Co., Amazonas, Venezuela, 03°58’2.32”N 67°15’12.49”W, 3 Mar 2005, N. K. Lujan, D. C. Werneke, M. H. Sabaj & M. Arce. Ancistrus marcapatae. 1902.5.29.211, 1, 76.8 mm SL, holotype of Chaetostomus marcapatae Regan, 1904. Marcapata Valley, eastern Peru. Ancistrus maximus de Oliveira, Zuanon, Zawadzki & Rapp Py-Daniel, 2015, NUP 14588, paratype, 1, 153.3 mm SL, Brazil, Roraima, Rorainópolis, Macoari stream, tributary to right side of rio Branco, 01°09’12.1”S 61°50’40.8”W, 2 Nov 2005, A. Negrão da Silva & M. L. Picanço. Ancistrus minutus Fisch-Muller, Mazzoni & Weber, 2001. MNRJ 20851, holotype, male, 57 mm SL, Brazil, Goiás, Minaçú, córrego Batéias, a left bank tributary of the upper rio Tocantins, 13º49’S 48º20’W, J. L. Costa Novaes, R. Mazzoni, F. Moraes & C. Weber, 5 Oct 1996. Ancistrus melas Eigenmann, 1916. FMNH 58339, holotype, 73.6 mm SL, synonym of Ancistrus centrolepis, Colombia, Condoto, 05º05’40”N 76º39’02”W. Ancistrus montanus (Regan, 1904), CAS-ICH 77039, 4, 54.9–53.5 mm SL, Bolivia, La Paz, Tumupasa, 30 miles northwest of Rurrenabaque, Dec 1921, N. E. Pearson. Ancistrus mullerae Bifi, Pavanelli & Zawadzki, 2009. MCP 22577, paratypes, 8 of 16, 52.4–64.4 mm SL, Catanduvas, stream at PR 477 road between Catanduvas and Três Barras do Paraná, 25°13’59”S 53°09’42”W. Ancistrus multispinis (Regan, 1912). MNRJ 1078, 70 (30, 44.5–106.7 mm SL), Brazil, Santa Catarina, Hanse (today county of Corupa), rio Humboldt, tributary of Itapocu river, Ehrhardt. Ancistrus nudiceps (Müller & Troschel, 1849). AUM 48095 (1, 81.8 mm SL), Takutu River, N of Sand Creek, rio Branco, rio Negro, Amazon basin, Guyana, 02°57’29”N 59°57’33”W, 15 Nov 2007, L. S. de Souza, D. C. Taphorn, J. N. Baskin, T. Geerinckx & J. L. Hwan. Ancistrus occloi Eigenmann, 1928.CAS-ICH 71818, paratype, male, 1, 87.4 mm SL, Peru, rio Urubamba (rio Santa Ana), Ollantayatambo Village, Incaic ruins, 22 Nov 1918, C. H. Eigenmann. Ancistrus parecis, MCP 35570, holotype, male, 55.9 mm SL, Brazil, Mato Grosso, Campos de Júlio, rio Formiga tributary of rio Juruena, upper rio Tapajós basin, on the road BR-364 (MT-235), between Campos de Júlio and Sapezal, 13°41’01”S 59°12’11”W, 13 Jul 2004, P. Lehmann, V. A. Bertaco, J. F. P. Silva & F. Langeani. Ancistrus patronus de Souza, Taphorn & Armbruster, 2019. AUM 43868, paratype, 1, 79.4 mm SL, Caño Soromoni, 11.8 km E of La Esmeralda, 03°11’38”N 65°39’07”W, 26 Mar 2005, N. Lujan, M. Arce-Hernández, L. Richmond, B. Grant & E. Wesley. Ancistrus pirareta Muller, 1989. MCP 13704, paratypes, 2, 71.3–77.9 mm SL, tributary of Río Tebicuary-mi, 25°30’S 56°55’W, Salto Pirareta, South of Piribebuy, Cordillera, Paraguay. Ancistrus piriformis Muller, 1989. MCP 13703, paratypes, 2, 63.3–65.0 mm SL, Río Acaray, below the lake of the dam, 25°23’S 54°42’W, Alto Paraná, Paraguay. Ancistrus ranunculus, MZUSP 47614, paratypes, 2, 50.9–56.3 mm SL, Pará, Cachoeira de Kaituká, rio Xingu, 9 Oct 1990, L. H. Rapp Py-Daniel & J. Zuanon. Ancistrus reisi Fisch-Muller, Cardoso, da Silva & Bertaco, 2005. MCP 34818, holotype, 60.4 mm SL, Brazil, Goiás, Mambaí, córrego das Dores, tributary of the rio Vermelho, 14°29’S 46°06’W, rio Tocantins basin, 15 Dec 2002, Centro de Biologia Aquática – UCG. Ancistrus saudades de Souza, Taphorn & Armbruster, 2019. AUM 48283, paratypes, 32 (2, 65.0–73.45 mm SL), Rupununi, Takutu River drainage, Moco-Moco Creek at Moco-Moco hydropower station, 18.8 km SE of Lethem, 03°17’47.87”N 59°38’42.25”W, 13 Nov 2007, L. de Souza, D. Taphorn, J. Baskin, T. Geerinckx & J. Hawn. AUM 67055, paratypes, (5, not measured), Tumong Creek, left-hand tributary of Ireng River, 04°42’50”N 60°01’20”W, 3 Jan 2016, N. Lujan, J. Armbruster, D. Werneke & M. Ram. Ancistrus tamboensis Fowler, 1945, MUSM 47882, 1, 83.3 mm SL, Peru, Rio Urubamba, 10°41’34.44”S 73°13’21.87”W. Ancistrus taunayi Miranda Ribeiro, 1918. MZUSP 975, lectotype, rio Lageado, Itaqui, Rio Grande do Sul, Brazil. Ancistrus tolima Taphorn, Armbruster, Villa-Navarro & Ray, 2013. AUM 54491, paratype, Colombia, Rio Magdalena drainage, Quebrada El Pescado, 03°35’57.50”N 74°51’16.40”W, vereda San Pablo, Dolores, departamento de Tolima, Colombia. Ancistrus tombador. MCP 33000, holotype, Brazil, Mato Grosso, Porto dos Gaúchos, igarapé Ribeirão Preto, on the road MT-338, about 26 km SE from Porto dos Gaúchos, 11°39’27”S 57°12’7”W, a tributary of rio Arinos, upper rio Tapajós basin, 19 Jan 2002, R. E. Reis, L.R. Malabarba, E. H. Pereira, V. A. Bertaco & A. R. Cardoso. Ancistrus triradiatus Eigenmann, 1918. AUM 36554, 3, 50.8–86.2 mm SL, Venezuela, Estado Bolívar, Rio Caroní – Rio Orinoco Drainage, Río Claro, 50.5 km SE of Ciudad Bolívar, 07°55’24.89”N 63°06’55.01”W, 10 Jun 2003, J. W. Armbruster, T. P. Pera, N. K. Lujan & O. Léon. Ancistrus verecundus Fisch-Muller, Cardoso, da Silva, Bertaco, 2005. MCP 35572, holotype, male, 52.2 mm SL, Brazil, Rondônia, Vilhena, igarapé Piracolina, about 6 km W of Vilhena, on the road BR-364, upper rio Madeira basin, 12°43’33”S 60°11’34”W, 14 Jul 2004, R. E. Reis, P. A. Buckup, A. R. Cardoso & E. H. Pereira.
Acknowledgments
We are grateful to Mário C. C. de Pinna and Michel Gianeti (MZUSP) for the loan of specimens and for Jonathan W. Armbruster for providing QGIS files for the map. EBN is supported by Proex/Capes (Proc. 88882.183270/2018–01). We thank Murilo Bastos (MNRJ) for information about Luzia in the Biological Anthropology collection and help with etymology. MRB is supported by grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, process #309285/2018–6305955/2015–2) and Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ, grant #200.103/2019). LMS received a CNPq Productivity Grant #309815/2017–7 and #311148/2020–4, and JLB received CNPq Productivity Grant #302872/2018–3. We also thank the Catfish Study Group for supporting the acquisition of the digital camera used to take the photos of the preserved specimens.
References
Ackerly DD, Thomas WW, Ferreira CAC, Pirani JR. The forest-cerrado transition zone in southern Amazonia: Results of the 1985 Projeto Flora Amazônica expedition to Mato Grosso. Brittonia. 1989; 41:113–28. https://doi.org/10.2307/2807515
Aquino AE, Schaefer SA. The temporal region of the cranium of loricarioid catfishes (Teleostei: Siluriformes): morphological diversity and phylogenetic significance. Zool Anz. 2002; 241(3):223–44. https://doi.org/10.1078/0044-5231-00071
Armbruster JW. Peckoltia sabaji, a new species from the Guyana Shield (Siluriformes: Loricariidae). Zootaxa. 2003; 344(1):1–12. https://doi.org/10.11646/zootaxa.344.1.1
Armbruster JW. Phylogenetic relationships of the suckermouth armoured catfishes (Loricariidae) with emphasis on the Hypostominae and the Ancistrinae. Zool J Linn Soc. 2004; 141(1):1–80. https://doi.org/10.1111/j.1096-3642.2004.00109.x
Buckup PA, Britto MR, Gomes JR, Birindelli JLO, Lima FCT, Maldonado-Ocampo JA, Zawadski CH, Carvalho FR, Jerep FC, Chamon CC, de Fries LCC, Vila Verde L, Camargo M, Souza-Lima R, Bartolette R, Wingert JM. Inventário da ictiofauna da ecorregião aquática Xingu-Tapajós. Rio de Janeiro: CETEM/MCT; 2011.
Dagosta FC, de Pinna M. Biogeography of Amazonian fishes: deconstructing river basins as biogeographic units. Neotrop Ichthyol. 2017; 15(3):e170034. https://doi.org/10.1590/1982-0224-20170034
Departamento Nacional de Infraestrutura de Transportes (DNIT). Hidrovia do Tapajós -Teles Pires [Internet]. Brazil; 2018. https://www.gov.br/dnit/pt-br/assuntos/aquaviario/hidrovia-do-tapajos-teles-pires
Dignall J. List of Original L-numbers; 2014. http://www.planetcatfish.com/shanesworld/shanesworld.php?article_id=514
Fearnside PM. Amazon dams and waterways: Brazil’s Tapajós Basin plans. Ambio. 2015; 44:426–39. https://doi.org/10.1007/s13280-015-0642-z
Fernandes FRC, Silva GR, Lima MHMR, Teixeira NS, Verde RV. A ecorregião aquática Xingu-Tapajós na Amazônia: Abordagem demográfica, econômica e social. Ecorregião Aquática Xingu-Tapajós. Rio de Janeiro: CETEM; 2011.
Fisch-Muller S, Cardoso AR, Silva JFP, Bertaco VA. Three new species of Ancistrus Kner (Teleostei: Siluriformes: Loricariidae) from the upper Tapajós and Tocantins rivers. Rev Suisse Zool. 2005a; 112(2):559–72. https://doi.org/10.5962/bhl.part.80314
Fisch-Muller S, Cardoso AR, da Silva JFP, Bertaco VA. Two new Amazonian species of armored catfishes (Siluriformes: Loricariidae): Ancistrus verecundus and Ancistrus parecis. Neotrop Ichthyol. 2005b; 3(4):525–32. https://doi.org/10.1590/S1679-62252005000400010
Fisch-Muller S, Mazzoni R, Weber C. Genetic and morphological evidences for two new sibling species of Ancistrus (Siluriformes: Loricariidae) in upper rio Tocantins, Brazil. Ichthyol Explor Freshw. 2001; 12(4):289–304.
Fricke R, Eschmeyer WN, Van der Laan R. Eschmeyer’s catalog of fishes: genera, species, references [Internet]. San Francisco: California Academy of Science; 2021. http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
International Union for Conservation of Nature (IUCN). Standards and Petitions Subcommittee. Guidelines for using the IUCN Red List categories and criteria, Version 14 [Internet], Cambridge, 2019. https://www.iucnredlist.org/resources/redlistguidelines.
Lucanus O, Kalacska M, Arroyo-Mora JP, Sousa L, Carvalho LN. Before and after: A multiscale remote sensing assessment of the Sinop Dam, Mato Grosso, Brazil. Earth. 2021; 2(2):303–30. https://doi.org/10.3390/earth2020018
Morris PJ, Yager HM, Sabaj Pérez MH. ACSImagebase: A digital archive of catfish images compiled by participants in the All Catfish Species Inventory; 2006. http://acsi.acnatsci.org/base
Muller S, Rapp Py-Daniel LH, Zuanon J. Ancistrus ranunculus, a new species of loricariid fish (Siluriformes: Loricariidae) from the Xingú and Tocantins rivers, Brazil. Ichthyol Explor Freshw. 1994; 5(4):289–96.
Neves WA, Powell JF, Prous A, Ozolins EG, Blum M. Lapa Vermelha IV Hominid 1: morphological affinities of the earliest known american. Genet Mol Biol. 1999; 22(4):461–69. https://doi.org/10.1590/S1415-47571999000400001
Ohara WM, Lima FCT, Salvador GN, Andrade MC. Peixes do rio Teles Pires: Diversidade e guia de identificação. MZUSP/UFPA/MZUEC; 2017.
Oliveira RR, Rapp Py-Daniel LH, Zawadzki CH, Zuanon J. Two new Amazonian species of Ancistrus with vestigial adipose fin, with an appraisal on adipose fin loss in neotropical armoured catfishes (Teleostei: Loricariidae). Ichthyol Explor Freshw. 2016; 27(1):67–80.
Sabaj MH. Codes for natural history collections in ichthyology and herpetology. Copeia. 2020; 108(3): 593–69. https://doi.org/10.1643/ASIHCODONS202
Sabaj MH, Armbruster JW, Page LM. Spawning in Ancistrus with comments on the evolution of snout tentacles as a novel reproductive strategy: larval mimicry. Ichthyol Explor Freshw. 1999; 10(3):217–29.
Schaefer SA. Osteology of Hypostomus plecostomus (Linnaeus) with a phylogenetic analysis of the loricariid subfamilies (Pisces: Siluroidei). Contributions in Science, Natural History Museum of Los Angeles County. 1987; 394(3–4):1–31. https://doi.org/10.5962/p.241283
Schaefer SA. The neotropical cascudinhos: systematics and biogeography of the Otocinclus catfishes (Siluriformes: Loricariidae). Proc Acad Nat Sci Phila. 1997; 148:1–120. https://www.jstor.org/stable/4065046
Stawikowski R. Harnischwelse aus südlichen Amazonaszuflüssen. Die Aquarien- und Terrarienzeitschrift (DATZ). 1988; 41:556–58.
Stawikowski R. Neu importiert: Loricariiden aus Brasilien. Die Aquarien- und Terrarienzeitschrift (DATZ). 1994; 47(3):145–46.
Taylor WR, Van Dyke GC. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium. 1985; 9(2):107–19. https://sfi-cybium.fr/en/node/2423
Zaiatz APSR, Zolin CA, Vendrusculo LG, Lopes TR, Paulino J. Agricultural land use and cover change in the Cerrado/Amazon ecotone: A case study of the upper Teles Pires River basin. Acta Amaz. 2018; 48(2):168–77. https://doi.org/10.1590/1809-4392201701930
Authors

![]() Emanuel B. Neuhaus1
Emanuel B. Neuhaus1 ![]() ,
, ![]() Marcelo R. Britto1,
Marcelo R. Britto1, ![]() José Luís O. Birindelli2 and
José Luís O. Birindelli2 and ![]() Leandro M. Sousa3
Leandro M. Sousa3
[1] Programa de Pós-graduação em Ciências Biológicas (Zoologia), Universidade Federal do Rio de Janeiro, Museu Nacional, Departamento de Vertebrados, Setor de Ictiologia, Quinta da Boa Vista, São Cristóvão, 20940-040 Rio de Janeiro, RJ, Brazil. (EBN) ebnsl@yahoo.com.br (corresponding author), (MRB) mrbritto@mn.ufrj.br.
[2] Universidade Estadual de Londrina, Centro de Ciências Biológicas, Departamento de Biologia Animal e Vegetal, Rodovia Celso Garcia Cid, km 380, Câmpus Universitário, 86051990 Londrina, PR, Brazil. josebirindelli@yahoo.com.
[3] Universidade Federal do Pará, Câmpus Altamira, Rua Coronel José Porfírio, 2515, São Sebastião, 68372-040 Altamira, PA, Brazil. leandro.m.sousa@gmail.com.
Authors’ Contribution 

Emanuel B. Neuhaus: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing-original draft, Writing-review and editing.
Marcelo R. Britto: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
José Luís O. Birindelli: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Leandro M. Sousa: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Ethical Statement
Not applicable.
Competing Interests
The authors declare no competing interests.
How to cite this article
Neuhaus EB, Britto MR, Birindelli JLO, Sousa LM. A new species of Ancistrus (Siluriformes: Loricariidae) from Tapajós and Xingu basins, Brazil. Neotrop Ichthyol. 2022; 20(1):e210129. https://doi.org/10.1590/1982-0224-2021-0129
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2022 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted December 15, 2021 by George Mattox
Accepted December 15, 2021 by George Mattox
![]() Submitted August 16, 2021
Submitted August 16, 2021
![]() Epub March 21, 2022
Epub March 21, 2022