![]() Liliana Paola Cárdenas-Mahecha1,
Liliana Paola Cárdenas-Mahecha1, ![]() Jean Ricardo Simões Vitule1,2 and
Jean Ricardo Simões Vitule1,2 and ![]() Andre Andrian Padial1,3,4
Andre Andrian Padial1,3,4 ![]()
PDF: EN XML: EN | Supplementary: S1 | Cite this article
Abstract
Non-native predators are known to cause ecological impacts through heightened consumption of resources and decimation of native species. One such species is Micropterus salmoides. Often introduced for sport fishing worldwide; it has been listed by International Union for Conservation of Nature and Natural Resources as one of the 100 of the world’s worst invasive species. Whilst impacts conferred towards native prey are well known, its relationship with other non-native species has received much less attention. Therefore, the aim of this study was to experimentally investigate the feeding preference of M. salmoides towards native prey (Geophagus iporangensis) compared to non-native prey(Oreochromis niloticus and Coptodon rendalli) using comparative consumption and the Manly-Chesson selectivity index. We found a higher consumption by M. salmoides towards the non-native prey and a tendency for this non-native preference to increase when prey availability increased. Our results are in line with the hypothesis that the invasion of the non-native predator can be facilitated by the high abundance and reproductive rate of the non-native prey. This is relevant considering the interaction of multiple invaders in an ecosystem, in particular environments modified by humans. Interspecific relationships are complex and their understanding is necessary for environmental management decision-making.
Keywords: Feeding preference, Invasional Meltdown, Micropterus salmoides, Multiple invasions, Tilapia.
Predadores não-nativos são conhecidos por causar impactos ecológicos com o aumento do consumo de recursos e do declínio das espécies nativas. Uma dessas espécies é Micropterus salmoides. Frequentemente introduzida ao redor do mundo para pesca esportiva; essa espécie foi listada pela União Internacional para a Conservação da Natureza como uma das 100 piores espécies invasoras do mundo. Embora os impactos nas presas nativas sejam bem conhecidos, sua relação com outras espécies não-nativas tem recebido muito menos atenção. Portanto, o objetivo deste estudo foi pesquisar experimentalmente a preferência alimentar de M. salmoides por presas nativas (Geophagus iporangensis) em comparação com presas não-nativas (Oreochromis niloticus e Coptodon rendalli) comparando o consumo e o índice de seletividade de Manly-Chesson. Encontramos que M. salmoides tem um maior consumo de presas não-nativas e uma tendência de aumento de preferência pela presa não-nativa com o aumento da disponibilidade de presas. Nossos resultados são consistentes com a hipótese de que a invasão do predador não-nativo pode ser facilitada pela grande abundância e capacidade reprodutiva de presas não-nativas. Isso é relevante considerando a interação de múltiplos invasores em um ecossistema, particularmente em ambientes modificados pelo homem. Relações interespecíficas são complexas e seu entendimento é necessário para a tomada de decisões de gestão ambiental.
Palavras-chave: Fusão invasora, Invasões múltiplas, Micropterus salmoides, Preferência alimentar, Tilápia.
Introduction
Introductions of non-native species are occurring at a growing rate as a result of human-mediated interests that include transport and trade (Frehse et al., 2016; Bezerra et al., 2019a,b; Vitule et al.,2019; Brito et al., 2020; Magalhães et al., 2020). Impacts caused by the introduction of non-native species are considered as one of the major causes of biotic change and occur across different levels of biological organization (Vitule et al., 2009; Cucherousset, Olden, 2011). Despite the fact that species introductions can often be observed over short time scales (Price et al., 2018), the long-term negative impacts can be irreversible, leading to the extinction of native species and the biotic homogenization of once diverse communities (Vitule et al., 2012; Bezerra et al., 2019b; Magalhães et al., 2020). In freshwater ecosystems, the negative impacts of non-native species can be particularly severe (Vörösmarty et al.,2000). Freshwater ecosystems are often heavily invaded by multiple species and, due to a high degree of endemism, can experience heightened extinction rates (Ricciardi, Rasmussen, 1999; Hudina et al., 2011; Burkhead, 2012; Daga et al., 2015). Non-native species are also often introduced for the purposes of aquaculture or sport fishing with many subsequently becoming invasive, especially in environments modified by humans, such as large reservoirs given the construction of dams (Bezerra et al., 2019b; Vitule et al., 2019; Brito et al., 2020).
Brazil is an example of country that has experienced the introduction of a number of highly invasive non-native species for a variety of reasons (Neuhaus et al.,2016; Franco et al.,2018). A globally invasive fish species is the largemouth bass Micropterus salmoides (Lacepède, 1802) of the family Centrarchidae. The native range of this species is from eastern North America to the Rio Grande basin in north-western Mexico (Beltrán Alvarez et al., 2013), however it has now been introduced for sport fishing in several continents around the world, including Asia (Ko et al.,2017), Europe (García-Berthou, Moreno-Amich, 2000; Costantini et al.,2018), Africa (Shelton et al., 2008; Ellender et al.,2014; Khosa et al., 2019), and South America (Garcia et al.,2014; Daga et al., 2015; Ribeiro et al., 2015; Pereira, Vitule, 2019). The largemouth bass usually has considerable impact as a voracious predator, demonstrated by its consumption on native fish and invertebrate prey (Abekura et al., 2004; Alexander et al., 2014), causing large shifts in the species composition and size structure of communities (Pereira, Vitule, 2019). This has led to the inclusion of the species in the list of 100 of the world’s worst invasive alien species by International Union for Conservation of Nature and Natural Resources (IUCN) (ISSG, 2013).
In 1922, M. salmoides was introduced in Brazil for sport fishing,becoming widely distributed in artificial systems, such as reservoirs in South and South-east regions (Schulz, Leal, 2005). Having reproduced and grown rapidly in semi-natural systems, it is now considered a threat to the conservation of Brazilian ecosystems (Schulz, Leal, 2005; Garcia et al., 2014; Daga et al.,2016). A number of species have also been introduced as human food resources, including tilapia species Oreochromis niloticus (Linnaeus, 1758) and Coptodon rendalli (Boulenger, 1897) (Canonico et al.,2005).
Phylogenetically-related and ecologically similar to tilapiine species, an important native cichlid species in Brazilian freshwaters (including reservoirs) is Geophagus iporangensis Haseman, 1911, which is popularly known as cará. Geophagus iporangensis was previously known as G. brasiliensis (Quoy & Gaimard, 1824), which represents is a complex of species across Brazilian basins (sensu Argolo et al., 2020), from the coastal basins of the Northeast Brazil to coastal rivers in Eastern Uruguay. The evolutionary complexity (Argolo et al., 2020) and the fact that G. iporangensis is one of the most abundant groups of cichlids in Brazil makes it a typical species in southern Brazilian freshwaters. G. iporangensis along with the tilapiines constitute a diet resource for invasive predatory fish like M. salmoides in southern Brazilian reservoirs (Bezerra et al., 2019b), and are indeed the most abundant fish species in such environments where they co-occur (Frehse et al., 2021). However, the predation pressure of the largemouth bass toward others invasive species such as the tilapiines is still not fully described, as well as how it compares to the phylogenetically-related and ecologically similar native prey.
The scenario of non-native species interacting with natives at multiple trophic levels is common in man-made ecosystems. In these cases, the knowledge of interspecific interactions is crucial to understand the cumulative impact of multiple invaders (Simberloff, Von Holle, 1999; Hudina et al.,2011). Different outputs are possible when multiple non-native species coexist in an ecosystem (Frehse et al., 2021). These outputs can be co-existence, biotic resistance (Twardochleb et al.,2012; Skein et al.,2020) or invasional meltdown as a consequence of the facilitation amongst invasive species (Simberloff, Von Holle, 1999). Even so, studies focusing impacts of non-native preys are still limited (Cattau et al., 2016); and studies that investigate the interaction of introduced predators foraging on non-native preys are even more limited. Johnson et al. (2009) focused in the advantage of non-native prey in avoiding predation compared to its native counterparts and the resulting synergistic effects of the positive association of the non-natives.
For a predator like M. salmoides, an important factor in prey selectivity is the availability of the resources (Pinnegar et al.,2003). When a preferred prey is poorly available and considering the optimal foraging theory (Pyke, 1984), M. salmoides likely behaves in an opportunistic manner, assuming opportunism as the behavior of taking advantage of the circumstances. Thus, in newly colonized ecosystems, its diet may reflect local prey abundance and availability (Young, Cockcroft, 1994).
We aim to experimentally investigate the feeding selectivity of M. salmoides towards non-native tilapiine complex prey (O. niloticus and C. rendalli) compared to a phylogenetically-related native prey species (G. iporangensis) considering resources availability. We cannot anticipate the predator preference, but if there is preference towards one species, implications are important for the understanding of the impacts of multiple invaders towards native species. Even so, we do expect that food preference of M. salmoides may depend of availability of resources in the scenario of high availability, the predator may chose the better prey, thus showing clearer patterns of preference. To analyse this hypothesis, we use Manly-Chesson’s index to determine the selective feeding between food items. This index may be the most meaningful indicator of prey type preference when preys are present in equal proportions (Confer, Moore, 1987) and is a useful measure for quantifying predator preference in selective predation, considering relative consumption and resource availability (Chesson, 1983).
Material and methods
We carried out a series of food preference experiments with fish that naturally coexist in reservoirs in southern Brazil. The introduced tilapiine species were chosen due to their high abundance and reproductive rate in these reservoirs, and their categorization as invasive species; while the native G. iporangensis prey was chosen because it is a typical and abundant species in the region.
The fishes used in the experiments were captured in March 2018 in the Passaúna reservoir located within Curitiba’s metropolitan area (between parallels 25° 15’– 25° 35’ S and meridians 49° 25’ – 49° 20’ W, see Sotiri et al. (2021) for an environmental description of the reservoir), Paraná State, Brazil. In this reservoir, the preys and the predator are the most abundant fish species (Frehse et al., 2021). Vouchers of species are all available at Museu de História Natural do Capão da Imbúia (MHNCI), Curitiba, Brazil. We collected small-sized predatory M. salmoides (15–20 cm total length, MHNCI 12484) and small sized prey (2–5 cm total length) consisting of non-native tilapias O. niloticus (MHNCI 12689) and C. rendalli (MHNCI 12130) and native prey G. iporangensis (MHNCI 12602). Although fish were not weighted, we argue that the standardization of size and the number of replicates minimized weight differences between experimental treatments (see below).
Fish collections were made under a permanent license for collection of zoological material SISBIO N° 24779. Prey were collected by casting a net trawl (mesh diameter 10 mm) near to the reservoir bank, and the predator M. salmoides was caught on rod and line. Upon collection, fishes were immediately transported to the laboratory where they were placed in 310 L water boxes for acclimation for at least two months where temperatures were maintained at 23 °C. Prey fishes were fed with commercial fish food, while predator was fed with beetle larvae, Tenebrio molitor Linnaeus, 1758 (Subhadra et al., 2006). During acclimation, both prey and predator were treated with Aqualife and Ictio (Labcon®; www.alconpet.com.br) to prevent disease.
The two non-native prey species were used together because they both belong to the same tribe, tilapiine within the family Cichlidae, with similar ecology and behavior (Canonico et al.,2005). Besides that, they are also representative of invasive species complexes found in the region (Cassemiro et al.,2018; Frehse et al., 2021). Experimental trials were conducted from May to December 2018 in 35 L black plastic boxes (experimental arenas) containing chlorine-free drinking and aerated water. The temperature was equally maintained at 23°C and experiments were conducted with natural light conditions. Individual predators were placed in an arena and then left without food for 72 h. After this time, native and non-native preys with similar body sizes were added simultaneously in equal amounts across a range of densities: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 13, 15, 16 fish per species origin (N = 5–7 arenas per density; a total of 76 experimental arenas). That is, for a density ‘1’, one native and one non-native prey were added. Predatory M. salmoides were left to interact with the prey fish for 48 h before they were removed and the amount of remaining prey counted. The experimental time and densities were determined after pilot experiments showing that, in this time interval the predator had enough time to consume at least half of the prey offered. It was also possible to reach the asymptote of available vs. consumed prey graph with the used densities – i.e., from this point on, consumption may remain constant. To eliminate possible noise from competition between preys, experimental arenas did not have shelters (Champneys et al., 2021). For logistical and ethical reasons, predatory M. salmoides (n = 20)were reused between treatments; however, they were only used once per fish density. After use in one trial, predators were placed in a tank for three days and fed a diet of T. molitor before 72 h without food and reused in a new trial. At the end of experiments, predators and remaining preys were euthanized using Benzocaine 80 mg/L or Eugenol 70–90%.
The choice of our experimental design was to compare feeding preference in a scenario of prey coexistence. We analyzed if there is a preference of M. salmoides by a type of prey (native or non-native) using the Manly-Chesson’s index with the following equation.

Where m is the total number of prey types consumed and compares the relative availability of a prey Pi with their relative consumption ri. Manly-Chesson’s index ranges from zero to one. If α = 1 / m, the predator feeds randomly and preys are consumed proportionally to their abundance in the environment. If α > 1 / m, the index indicates preference; and α < 1 / m indicates avoidance (Cochran-Biederman, Vondracek, 2017). Manly-Chesson’s index is a function of forage ratio (forage proportion) (ri / Pi), therefore the sum of all α for a predator is normalized to 1. Therefore, a higher median Manly-Chesson’s value would indicate a more specialist feeding strategy, with consumers feeding heavily on a few species, rather than feeding weakly on many types (Confer, Moore, 1987). In ours experimental, given that the types of prey were offered in equal proportions, Pi is always 0.5, and the Manly-Chesson index, for a non-native or native prey, is equal to the relative consumption of that prey (ri):

We performed the Manly-Chesson index with the dietr package (Borstein, 2020), function Electivity in R (R Development Core Team, 2018). Once the index values were obtained, they were related with the prey availability using a simple linear regression. In this case, the index was calculated for non-native prey type and represents the relative consumption of non-native prey, for that the higher the index, the higher was the consumption towards the non-native prey. Therefore, Manly-Chesson’s index was considered a response variable that indicate the effect size of prey selectivity, and the pattern of the regression against prey density would demonstrate the relationship between food preference and availability, accounting for the fact that preys were simultaneously offered in the same experimental unit. Additionally, we carried out a functional response analysis to reach the maximum consumption rate of the predator (S1).
Results
We found that the relative consumption of non-native prey type is higher when prey availability increases compared to native prey (Fig. 1). Indeed, the simple linear regression show that Manly-Chesson index was related to prey availability, increasing towards the non-native prey as availability increases (Manly-Chesson for non-native prey = 0.5 + 0.007 Prey Availability, R2 = 0.031, F1, 74, p = 0.012) (Fig. 2). Even so, it was clear that M. salmoides showed higher consumption of non-native prey compared to native particularly at high prey densities.

FIGURE 1| Relative consumption of non-native (Oreochromis niloticus and Coptodon rendalli) and native (Geophagus iporangensis) prey, considering different prey availability for Micropterus salmoides.
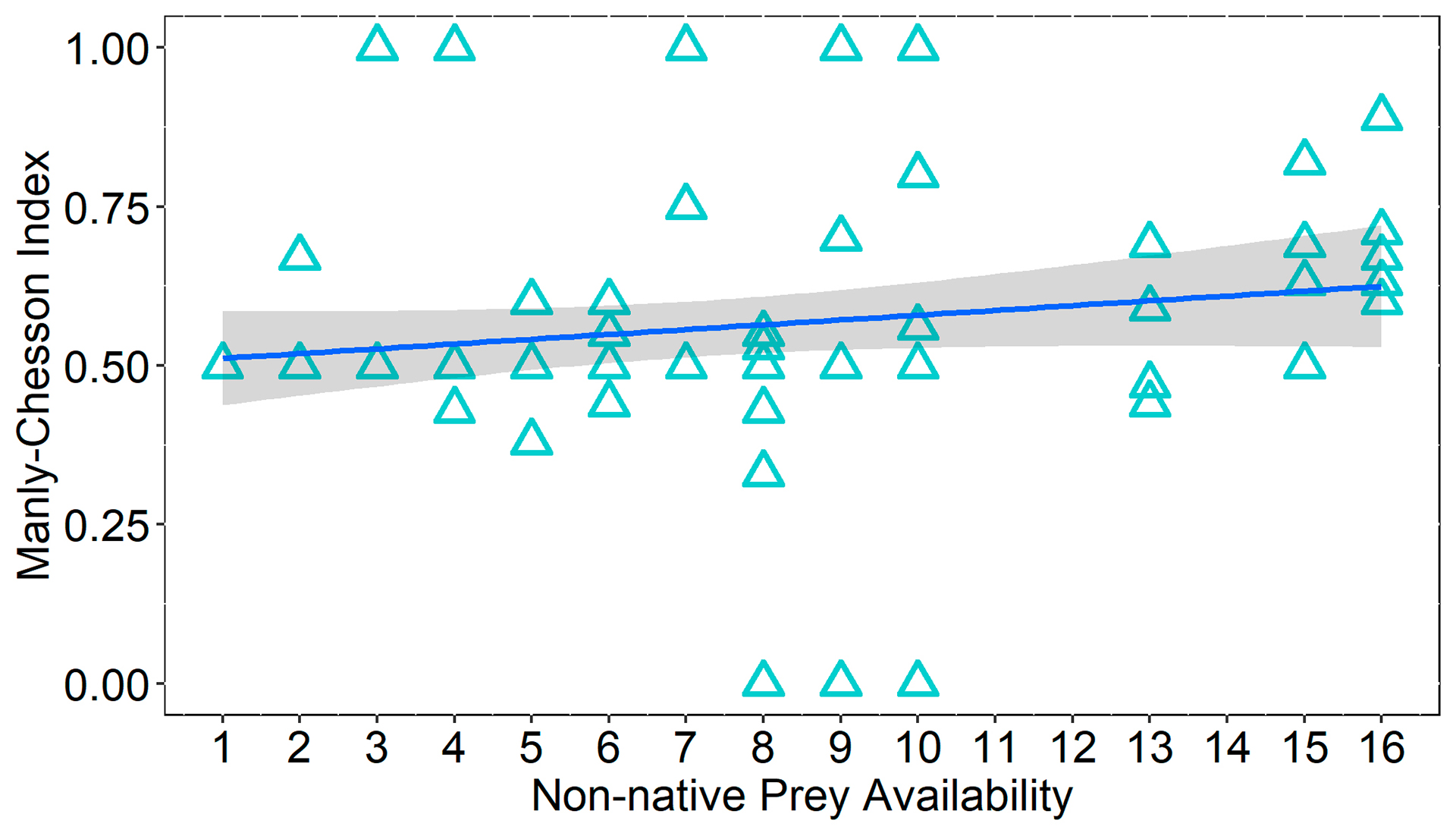
FIGURE 2 | Relationship between the Manly-Chesson selectivity and prey availability for Micropterus salmoides. Higher values indicate preference for non-native species.Shading represents 95% confidence intervals. Note that because the index fluctuates between 0 and 1, with 2 types of prey and equal availability of prey for both types, the result of the index for one prey is exactly the opposite of the other. For this reason, the graph only shows the results of the index for the non-native species. The graph for the other type of prey would be the spectral image of this one.
Discussion
We generated evidence that the invasive M. salmoides consumes a higher number of non-native prey cichlids compared to natives, when offered simultaneously at equal densities, especially at high prey densities. It was indeed expected that an opportunistic and generalist behavior of M. salmoides occur particularly when prey densities were low. Opportunistic behavior at low densities could increase the predation effect on native prey. We emphasize that this refers only to a generalist behavior regarding these two types of prey and it is not possible to generalize this behavior to the entire range of the diet, given the experiment was limited to only three typical preys of the predator. It was clear in our experimental essay that both prey types were vulnerable to active hunting by M. salmoides, also an expected scenario given the absence of shelters and the ecomorphological traits of this predator (Luger et al.,2020). Considering prey size, the results obtained were in line with those obtained by Cuthbert et al.(2020) suggesting that even small and intermediate M. salmoides exhibit higher attack rates in small and intermediate preys. We expected that our results also reflected behavioural differences between the two prey types. The native prey was observed to be less active compared to the non-native prey. Thus, the natives can benefit from an anti-predator behaviour of taking cover, “freezing” or reducing activity during certain times of day to avoid being detected by a chasing predator. This could indicate an important role in trade-off between risk of predation and foraging and other fitness-enhancing activities (Clark, 1994).
It is important to note that heightened consumption on non-native prey may have little impact on the exotic tilapiine populations in Passaúna reservoir as their abundance is far higher than the native prey G. iporangensis (estimated as 4.96 ton*km-2*year-1 compared to 1 ton*km-2*year-1, respectively) as well as their reproductive rates (see Bezerra et al.,2019b). The higher consumption of individuals from the successful populations of O. niloticus and C. rendalli in the reservoirs of the Metropolitan Region of Curitiba may thus indicate a positive effect of the invasive preys in sustaining M. salmoides populations without being affected by top-down control (see Bezerra et al., 2019b). Thus, changes in the biomass of O. niloticus and C. rendalli and its hybrids can result in significant changes in the biomass of M. salmoides (Bezerra et al.,2019b). Our results add evidence for a positive effect between interacting non-native species, which can result in the ‘Invasional Meltdown’ phenomenon (Simberloff, Von Holle, 1999; Kuebbing et al.,2013; Braga et al.,2018). Interestingly, such facilitation scenario was generated by a ‘negative’ predator-prey ecological interaction, which at a first glance seems counterintuitive, but well explained by the biology of the tilapiine species. Given the high reproduction rate and successful colonization in the urban reservoirs (Starling et al., 2002; Sánchez-Botero et al., 2014), we argue that tilapias are not negatively affected by a voracious invasive predator, but instead facilitate the establishment of the invasive predator (see Bezerra et al., 2019b). Such impact of tilapias is still underestimated, particularly in Brazil where there is a strong movement of denial against tilapias impacts (Charvet et al., 2021). Finally, it is important to note that results became even more relevant given the preys and the predator studied here are the most abundant fish species in the Passaúna reservoir (Frehse et al., 2021).
Indeed, the present study has contributed to our understanding of trophic relationships between native and non-native species. Taken together, patterns were in accordance with others that highlight the opportunistic behaviour of M. salmoides at low densities (Hodgson, Kitchell, 1987) and depending of prey characteristics (morphology, behaviour, abundance, availability), which consequently dictate its feeding strategy at high densities (Taylor et al.,2019; Luger et al.,2020). Further, invaders such as M. salmoides have a great phenotypic plasticity, even acting at different trophic levels (Almeida et al.,2012). The trophic level at which opportunistic invasive species establish can be determined by the ecological characteristics of invaded communities. For example, Costantini et al.(2018) showed that M. salmoides can change their feeding habits and their trophic level depending on the availability of prey. Additionally, this predator played an important role within trophic networks, as changes in their populations could generate cascading effects (Schindler et al.,1997).
As a result of the current dynamics of globalization, most ecosystems have suffered from simultaneous introduction of several invasive species, adding complexity in the understanding of species interactions (Simberloff, Von Holle, 1999). Most research, however, remains on the study of single-species invasions (Magalhães, Ratton, 2005; Hudina et al.,2011; Costa-Novaes, Carvalho, 2012; Kuebbing et al.,2013). Quantifying and predicting the negative impacts of multiple non-native invasive species are highly important for conservation in diverse regions such as Neotropics; particularly in human altered environments. Here we have shown how the presence of multiple invasive fish with high reproductive rates may result in synergistic effects, with greater potential damage to natural ecosystems. We also provided an approach of experimentation and analysis to address the scenario of multiple invasions, especially in Neotropical water bodies. We highlighted the probability of increased impact when invasive species coexist in the same ecosystem, even when the predominant interaction between invasive species is a ‘negative’ ecological interaction, such as predator-prey.
Acknowledgments
To the Universidade Federal de Paraná, especially to the Pós-Graduação em Ecologia e Conservação and to the laboratories of Análise e Síntese em Biodiversidade (LASB) and Ecologia e Conservação (LEC). To the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for a Master scholarship granted to LPCM (Financial code 001) and to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the financial support. AAP and JRSV are grateful for grants from CNPq (PQ Process numbers: 310850/2012–6; 303776/2015–3; 307984/2015–0; 402828/2016–0; 301867/2018–6). We are also grateful for anonymous reviewers and the editors for valuable comments in previous drafts.
References
Abekura K, Hori M, Takemon Y. Changes in fish community after invasion and during control of alien fish populations in Mizoro-ga-ike, Kyoto City. Global Environ Res. 2004; 8(2):145–54.
Alexander ME, Dick JTA, Weyl OLF, Robinson TB, Richardson DM. Existing and emerging high impact invasive species are characterized by higher functional responses than natives. Biol Lett. 2014; 10(2):e20130946. https://doi.org/10.1098/rsbl.2013.0946
Almeida D, Almodóvar A, Nicola GG, Elviraa B, Grossmanc GD. Trophic plasticity of invasive juvenile largemouth bass Micropterus salmoides in Iberian streams. Fish Res. 2012; 113(1):153–58. https://doi.org/10.1016/j.fishres.2011.11.002
American Veterinary Medical Association (AVMA). Guidelines for euthanasia of animals. ed. Schaumburg: American Veterinary Medical Association. 2013.
Argolo LA, López-Fernández H, Batalha-Filho H, Mello Affonso PRA. Unraveling the systematics and evolution of the ‘Geophagus’ brasiliensis (Cichliformes: Cichlidae) species complex. Mol Phylogenet Evol, 2020; 150:106855. https://doi.org/10.1016/j.ympev.2020.106855
Beltrán Alvarez R, Sánchez Palacios J, Ramírez Lozano JP, Ortega Salas AA. Reproducción de Micropterus salmoides (Pisces: Centrarchidae), en el embalse Gustavo Díaz Ordaz, Sinaloa, México. Rev Biol Trop. 2013; 61(3):1313–25. Available from: https://www.scielo.sa.cr/pdf/rbt/v61n3/a25v61n3.pdf
Bezerra LAV, Freitas MO, Daga VS, Occhi TVT, Faria L, Costa APL, Padial AA, Prodocimo, Vitule JRS. A network meta-analysis of threats to South American fish biodiversity. Fish Fish. 2019a; 20(4):620–39. https://doi.org/10.1111/faf.12365
Bezerra LAV, Ribeiro VM, Freitas MO, Kaufman L, Padial AA, Vitule JRS. Benthification, biotic homogenization behind the trophic downgrading in altered ecosystems. Ecosphere. 2019b; 10(6):e02757. https://doi.org/10.1002/ecs2.2757
Borstein SR. dietr: an R package for calculating fractional trophic levels from quantitative and qualitative diet data. Hydrobiologia. 2020; 847:4285–94. Available from: https://github.com/sborstein/dietr
Braga RR, Gómez-Aparicio L, Heger T, Vitule JRS, Jeschke JM. Structuring evidence for invasional meltdown: broad support but with biases and gaps. Biol Invasions. 2018; 20:923–36. https://doi.org/10.1007/s10530-017-1582-2
Brito MFG, Daga VS, Vitule JRS. Fisheries and biotic homogenization of freshwater fish in the Brazilian semiarid region. Hydrobiologia. 2020; 847:3877–95.
Burkhead NM. Extinction rates in north american freshwater fishes, 1900–2010. Bioscience. 2012; 62(9):798–808. https://doi.org/10.1525/bio.2012.62.9.5
Canonico GC, Arthington A, Mccrary JK, Thieme ML. The effects of introduced tilapias on native biodiversity. Aquat Conserv Mar Freshwater Ecosyst. 2005; 15(5):463–83. https://doi.org/10.1002/aqc.699
Cassemiro FAS, Bailly D, Graça WJ, Agostinho AA. The invasive potential of tilapias (Osteichthyes, Cichlidae) in the Americas. Hydrobiologia. 2018; 817:133–54.
Cattau CE, Fletcher Jr. RJ, Reichert BE, Kitchens WM. Counteracting effects of a non-native prey on the demography of a native predator culminate in positive population growth. Ecol Appl. 2016; 26(7):1952–68. https://doi.org/10.1890/15-1020.1
Champneys T, Genner MJ, Ioannou CC. Invasive Nile tilapia dominates a threatened indigenous tilapia in competition over shelter. Hydrobiologia. 2021; 848:3747–62. https://doi.org/10.1007/s10750-020-04341-8
Charvet P, Occhi TVT, Faria L, Carvalho B, Pedroso CR, Carneiro L, Freitas M, Petrere-Junior M, Vitule JRS. Tilapia farming threatens Brazil’s waters. Science. 2021; 371(6527):356. https://doi.org/10.1126/science.abg1346
Chesson J. The estimation and analysis of preference and its relationship to foraging models. Ecology. 1983; 64(5):1297–304. https://doi.org/10.2307/1937838
Clark CW. Antipredator behaviour and the asset-protection principle. Behav Ecol. 1994; 5(2):159–70. https://doi.org/10.1093/beheco/5.2.159
Cochran-Biederman JL, Vondracek B. Seasonal feeding selectivity of brown trout Salmo trutta in five groundwater-dominated streams. J Freshwater Ecol. 2017; 32(1):653–73. https://doi.org/10.1080/02705060.2017.1386595
Confer JL, Moore MV. Interpreting selectivity indices calculated from field data or conditions of prey replacement. Can J Fish Aquat Sci. 1987; 44(9):1529–33. https://doi.org/10.1139/f87-184
Costa-Novaes JL, Carvalho ED. Reproduction, food dynamics and exploitation level of Oreochromis niloticus (Perciformes: Cichlidae) from artisanal fisheries in Barra Bonita Reservoir, Brazil. Rev Biol Trop. 2012; 60(2):721–34. Available from: https://repositorio.unesp.br/handle/11449/73365
Costantini ML, Carlino P, Calizza E, Careddu G, Cicala D, Caputi SS, Fiorentino F, Rossi L. The role of alien fish (the centrarchid Micropterus salmoides) in lake food webs highlighted by stable isotope analysis. Freshw Biol. 2018; 63(9):1130–42. https://doi.org/10.1111/fwb.13122
Cucherousset J, Olden JD. 2011. Ecological Impacts of Nonnative Freshwater Fishes. Fisheries. 2011; 36(5): 215–30. https://doi.org/10.1080/03632415.2011.574578
Cuthbert RN, Wasserman RJ, Dalu T, Kaiser H, Weyl OLF, Dick JTA, Sentis A, McCoy MW, Alexander ME. Influence of intra- and interspecific variation in predator–prey body size ratios on trophic interaction strengths. Ecol Evol. 2020; 10(12):5946–62. https://doi.org/10.1002/ece3.6332
Daga VS, Debona T, Abilhoa V, Gubiani EA, Vitule JRS. Non-native fish invasions of a Neotropical ecoregion with high endemism: a review of the Iguaçu River. Aquat Invasions. 2016; 11(2):209–23. http://dx.doi.org/10.3391/ai.2016.11.2.10
Daga VS, Skóra F, Padial AA, Abilhoa V, Gubiani EA, Vitule JRS. Homogenization dynamics of the fish assemblages in Neotropical reservoirs: comparing the roles of introduced species and their vectors. Hydrobiologia. 2015; 746:327–47. https://doi.org/10.1007/s10750-014-2032-0
Ellender BR, Woodford DJ, Weyl OLF, Cowx IG. Managing conflicts arising from fisheries enhancement based on non-native fishes in southern Africa. J Fish Biol. 2014; 85(6):1890–906. https://doi.org/10.1111/jfb.12512
Franco ACS, Santos LN, Petry AC, García-Berthou E. Abundance of invasive peacock bass increases with water residence time of reservoirs in southeastern Brazil. Hydrobiologia. 2018; 817:155–66. https://doi.org/10.1007/s10750-017-3467-x
Frehse FA, Braga RR, Nocera GA, Vitule JRS. Non-native species and invasion biology in a megadiverse country: scientometric analysis and ecological interactions in Brazil. Biol Invasions. 2016; 18:3713–25. https://doi.org/10.1007/s10530-016-1260-9
Frehse FA, Weyl OLF, Vitule JRS. Differential use of artificial habitats by native and non-native fish species in Neotropical reservoirs. Hydrobiologia. 2021; 848: 2355–67. https://doi.org/10.1007/s10750-021-04564-3
García-Berthou E, Moreno-Amich R. Introduction of exotic fish into a Mediterranean lake over a 90-year period. Arch Hydrobiol. 2000; 149(2):271–84. https://doi.org/10.1127/archiv-hydrobiol/149/2000/271
Garcia DAZ, Costa ADA, Leme GA, Orsi ML. Biology of black bass Micropterus salmoides (Lacepède, 1802) fifty years after the introduction in a small drainage of the Upper Paraná River basin, Brazil. Biodiversitas. 2014; 15:180–85. https://doi.org/10.13057/BIODIV%2FD150209
Hodgson JR, Kitchell JF. Opportunistic foraging by largemouth bass (Micropterus salmoides). Am Midl Nat. 1987; 118(2):323–36. https://doi.org/10.2307/2425789
Hudina S, Galić N, Roessink I, Hock K. Competitive interactions between co-occurring invaders: identifying asymmetries between two invasive crayfish species. Biol Invasions. 2011; 13:1791–803. https://doi.org/10.1007/s10530-010-9933-2
Invasive Species Specialist Group (ISSG). 100 of the world’s worst invasive alien species. Global Invasive Species Database. University of Auckland, New Zealand. 2013. Available from: http://www.iucngisd.org/gisd/100_worst.php
Johnson PTJ, Olden JD, Solomon CT. Interactions among invaders: community and ecosystem effects of multiple invasive species in an experimental aquatic system. Oecologia. 2009; 159:161–70. https://doi.org/10.1007/s00442-008-1176-x
Khosa D, Marr SM, Wasserman RJ, Zengeya TA, Weyl OLF. An evaluation of the current extent and potential spread of black bass invasions in South Africa. Biol Invasions. 2019; 21:1721–36. https://doi.org/10.1007/s10530-019-01930-0
Ko M-H, Kwan Y-S, LeeW-K, Won Y-J. Impact of human activities on changes of ichthyofauna in Dongjin River of Korea in the past 30 years. Anim Cells Syst. 2017; 21(3):207–16. https://doi.org/10.1080/19768354.2017.1330223
Kuebbing SE, Nuñez MA, Simberloff D. Current mismatch between research and conservation efforts: The need to study co-occurring invasive plant species. Biol Conserv. 2013; 160:121–29. https://doi.org/10.1016/j.biocon.2013.01.009
Luger AM, South J, Alexander ME, Ellender BR, Weyl OLF, Nagelkerke LAJ. Ecomorphology of largemouth bass relative to a native trophic analogue explains its high invasive impact. Biol Invasions. 2020; 22:2223–33. https://doi.org/10.1007/s10530-020-02252-2
Magalhães ALB, Daga VS, Bezerra LAV, Vitule JRS, Jacobi CM, Silva LGM. All the colors of the world: biotic homogenization-differentiation dynamics of freshwater fish communities on demand of the Brazilian aquarium trade. Hydrobiologia. 2020; 847:3897–915. https://doi.org/10.1007/s10750-020-04307-w
Magalhães ALB, Ratton TF. Reproduction of a South American population of pumpkinseed sunfish Lepomis gibbosus (Linnaeus) (Osteichthyes, Centrarchidae): a comparison with the European and North American populations. Rev Bras Zool. 2005; 22(2):477–83. https://doi.org/10.1590/S0101-81752005000200025
Neuhaus EB, Antonetti DA, Schulz UH. The invasive fish Acestrorhynchus pantaneiro Menezes, 1992 in Southern Brazil: Occurrence and food niche overlap with two native species. J Appl Ichthyol. 2016; 32(6):1107–12. https://doi.org/10.1111/jai.13147
Pereira FW, Vitule JRS. The largemouth bass Micropterus salmoides (Lacepède, 1802): impacts of a powerful freshwater fish predator outside of its native range. Rev Fish Biol Fish. 2019; 29:639–52. https://doi.org/10.1007/s11160-019-09570-2
Pinnegar JK, Trenkel VM, Tidd AN, Dawson WA, Du buit MH. Does diet in Celtic Sea fishes reflect prey availability? J Fish Biol. 2003; 63(1):197–212. https://doi.org/10.1111/j.1095-8649.2003.00204.x
Price EPF, Spyreas G, Matthews JW. Biotic homogenization of regional wetland plant communities within short time-scales in the presence of an aggressive invader. J Ecol. 2018; 106(3):1180–90. https://doi.org/10.1111/1365-2745.12883
Pyke GH. Optimal foraging theory: A critical review. Annu Rev Ecol Syst. 1984; 15:523–75. https://doi.org/10.1146/annurev.es.15.110184.002515
R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. 2018. http://www.r-project.org.
Ribeiro VM, Braga RR, Abilhoa V, Vitule JRS. Evaluation of three capture techniques for invasive Micropterus salmoides (Lacépède, 1802) in a Neotropical reservoir: implications for population control and management. J Appl Ichthyol. 2015; 31(6):1127–29. https://doi.org/10.1111/jai.12904
Ricciardi A, Rasmussen JB. Extinction rates of North American freshwater fauna. Conserv Biol J. 1999; 13(5):1220–22. https://doi.org/10.1046/j.1523-1739.1999.98380.x
Sánchez-Botero JI, Reis VC, Chaves FDN, Garcez DS. Fish assemblage of the Santo Anastácio Reservoir (Ceará State, Brazil). Bol Inst Pesca. 2014; 40(1):1–15. Available from: https://www.pesca.sp.gov.br/boletim/index.php/bip/article/view/1015
Schindler DE, Carpenter SR, Cole JJ, Kitchell JF, Pace ML. Influence of food web structure on carbon exchange between lakes and the atmosphere. Science. 1997; 277(5323):248–51. https://doi.org/10.1126/science.277.5323.248
Schulz UH, Leal ME. Growth and mortality of black bass, Micropterus salmoides (Pisces, Centrarchidae, Lacépède, 1802) in a reservoir in southern Brazil. Braz J Biol. 2005; 65(2):363–69. https://doi.org/10.1590/S1519-69842005000200021
Shelton JM, Day JA, Griffiths CL. Influence of largemouth bass, Micropterus salmoides, on abundance and habitat selection of Cape galaxias, Galaxias zebratus, in a mountain stream in the Cape Floristic Region, South Africa. Afr J Aquat Sci. 2008; 33(3):201–10. https://doi.org/10.2989/AJAS.2008.33.3.2.614
Simberloff D, Von Holle B. Positive interactions of nonindigenous species: Invasional meltdown? Biol Invasions. 1999; 1:21–32. https://doi.org/10.1023/A:1010086329619
Skein L, Alexander ME, Robinson TB. Co-occurring predators increase biotic resistance against an invasive prey. Mar Environ Res. 2020; 157:104929. https://doi.org/10.1016/j.marenvres.2020.104929
Sotiri K, Hilgert S, Mannich M, Bleninger T, Fuchs S. Implementation of comparative detection approaches for the accurate assessment of sediment thickness and sediment volume in the Passaúna Reservoir. J Environ Manage. 2021; 287:112298. https://doi.org/10.1016/j.jenvman.2021.112298
Starling F, Lazzaro X, Cavalcanti C, Moreira R. Contribution of omnivorous tilapia to eutrophication of a shallow tropical reservoir: evidence from a fish kill. Freshw Biol. 2002; 47(12):2443–52. https://doi.org/10.1046/j.1365-2427.2002.01013.x
Subhadra B, Lochmann R, Rawles S, Chen R. Effect of dietary lipid source on the growth, tissue composition and hematological parameters of largemouth bass (Micropterus salmoides). Aquaculture. 2006; 255(1–4):210–22. https://doi.org/10.1016/j.aquaculture.2005.11.043
Taylor GC, Hill JM, Weyl OLF. The diet and trophic ecology of non-native Micropterus salmoides in two South African impoundments. Afr J Aquat Sci. 2019; 44(2):143–53. https://doi.org/10.2989/16085914.2019.1612318
Twardochleb LA, Novak M, Moore JW. Using the functional response of a consumer to predict biotic resistance to invasive prey. Ecol Appl. 2012; 22(4):1162–71. https://doi.org/10.1890/11-0871.1
Vitule JRS, Freire CA, Simberloff D. Introduction of non-native freshwater fish can certainly be bad. Fish Fish. 2009; 10(1):98–108. https://doi.org/10.1111/j.1467-2979.2008.00312.x
Vitule JRS, Occhi TVT, Kang B, Matsuzaki S-I, Bezerra LA, Daga VS, Faria L, Frehse FA, Walter F, Padial AA. Intra-country introductions unraveling global hotspots of alien fish species. Biodivers Conserv. 2019; 28:3037–43. https://doi.org/10.1007/s10531-019-01815-7
Vitule JRS, Skóra F, Abilhoa V. Homogenization of freshwater fish faunas after the elimination of a natural barrier by a dam in Neotropics. Divers Distrib. 2012; 18(2):111–20. https://doi.org/10.1111/j.1472-4642.2011.00821.x
Vörösmarty CJ, Green P, Salisbury J, Lammers RB. Global water resources: vulnerability from climate change and population growth. Science. 2000; 289:284–88.
Young DD, Cockcroft VG. Diet of common dolphins (Delphinus delphis) off the south-east coast of southern Africa: opportunism or specialization? J Zool. 1994; 234(1):41–53. https://doi.org/10.1111/j.1469-7998.1994.tb06055.x
Authors

![]() Liliana Paola Cárdenas-Mahecha1,
Liliana Paola Cárdenas-Mahecha1, ![]() Jean Ricardo Simões Vitule1,2 and
Jean Ricardo Simões Vitule1,2 and ![]() Andre Andrian Padial1,3,4
Andre Andrian Padial1,3,4 ![]()
[1] Laboratório de Análise e Síntese em Biodiversidade, Departamento de Botânica. Programa de Pós-Graduação em Ecologia e Conservação, Setor de Ciências Biológicas, Universidade Federal do Paraná, Av. Cel. Francisco H. dos Santos, 100, Jardim das Américas, 81531-980 Curitiba, PR, Brazil. (LPCM) lilipa6000@gmail.com, (JRSV) biovitule@gmail.com, (AAP) aapadial@gmail.com (corresponding author).
[2] Laboratório de Ecologia e Conservação, Departamento de Engenharia Ambiental, Setor de Tecnologia, Universidade Federal do Paraná, Av. Cel. Francisco H. dos Santos, 100, Jardim das Américas, 81531-980 Curitiba, PR, Brazil.
[3] Programa de Pós-graduação em Botânica, Departamento de Botânica, Setor de Ciências Biológicas, Universidade Federal do Paraná, Av. Cel. Francisco H. dos Santos, 100, Jardim das Américas, 81531-980 Curitiba, PR, Brazil.
[4] Programa de Pós-graduação em Ecologia de Ambientes Aquáticos Continentais, Núcleo de Pesquisa em Limnologia, Ictiologia e Aquicultura, Universidade Estadual de Maringá, Av. Colombo, 5790, Bloco G-90, 87020-900 Maringá, PR, Brazil.
Authors’ Contribution 

Liliana Paola Cárdenas-Mahecha: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Validation, Visualization, Writing-original draft, Writing-review and editing.
Jean Ricardo Simões Vitule: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Resources, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Andre Andrian Padial: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Ethical Statement
The experiment was carried out following the protocols of Conselho Nacional de Controle de Experimentação Animal (CEUA) and the American Veterinary Medical Association (AVMA, 2013); and all animals used were euthanized according to the procedures of Divisão de Gestão Ambiental da Universidade Federal do Paraná (UFPR). The use of animals in the experiments and the procedures performed were authorized by the certificates N° 1027 and 1199 of CEUA from UFPR.
Competing Interests
The authors declare no competing interests.
How to cite this article
Cárdenas-Mahecha LP, Vitule JRS, Padial AA. Prey selectivity of the invasive largemouth bass towards native and non-native prey: an experimental approach. Neotrop Ichthyol. 2022; 20(2):e210123. https://doi.org/10.1590/1982-0224-2021-0123
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2022 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted May 22, 2022 by Ana Petry
Accepted May 22, 2022 by Ana Petry
![]() Submitted July 28, 2021
Submitted July 28, 2021
![]() Epub June 24, 2022
Epub June 24, 2022