Karla D. A. Soares1 ![]()
![]() , Renan A. Moreira1
, Renan A. Moreira1 ![]() and Ulisses L. Gomes2
and Ulisses L. Gomes2 ![]()
PDF: EN XML: EN | Cite this article
Abstract
Rioraja agassizii belongs to the monotypic genus Rioraja and differs from the other Riorajini species of the genus Atlantoraja by the shape of anterior nasal flap, squamation, clasper skeleton, body measurements and color pattern. Although R. agassizii is a well-known species, commonly captured in inshore waters of the southwestern Atlantic, its external morphology and internal anatomy have never been fully described and little is known on intraspecific variation within the species. Juvenile and adult, male and female specimens were examined, measured and dissected. Herein, we redescribe the species and present detailed descriptions and illustrations of external morphology, neurocranium, jaws, hyoid and gill arches, and pelvic fin and girdle. Additionally, notes on sexual dimorphism and ontogenetic differences in coloration, body and cranial measurements, and dermal denticles are provided.
Keywords: Anatomy, Sexual dimorphism, Taxonomy.
Rioraja agassizii pertence ao gênero monotípico Rioraja e difere das demais espécies da tribo Riorajini classificadas no gênero Atlantoraja pelo formato da aba nasal anterior, escamas, esqueleto do clásper, medidas corpóreas e padrão de coloração. Embora R. agassizii seja uma espécie bem conhecida, comumente capturada em águas costeiras do Atlântico Sul Ocidental, sua morfologia externa e anatomia interna não foram totalmente descritas e pouco se sabe sobre a variação intraespecífica nesta espécie. Espécimes juvenis, adultos, machos e fêmeas foram examinados, medidos e dissecados. Neste estudo, a espécie é redescrita e descrições detalhadas e ilustrações da morfologia externa, neurocrânio, maxilas, arcos hioide e branquiais, nadadeira e cintura pélvicas são apresentadas. Adicionalmente, notas sobre dimorfismo sexual e variação ontogenética na coloração, medidas corpóreas e craniais, e dentículos dérmicos são fornecidas.
Palavras-chave: Anatomia, Dimorfismo sexual, Taxonomia.
Introduction
The genus Rioraja Whitley, 1929 belongs to the tribe Riorajini McEachran & Dunn, 1998 together with other three species of Atlantoraja: the La Plata skate, A. platana (Günther, 1880), the eyespot skate, A. cyclophora (Regan, 1903), and the spotback skate, A. castelnaui (Miranda Ribeiro, 1907). Representatives of this tribe are endemic to the inshore waters in the southwestern Atlantic, from Espírito Santo (Brazil) to Argentina (Miranda Ribeiro, 1907, 1923; Evermann, Kendall, 1907; Ruschi, 1965; Menni, 1973; Roux, 1977; Menni et al., 1984; Cousseau et al., 2000, 2007; Meneses, Paesch, 2003; Last et al., 2016).
Rioraja is a monotypic genus represented by the Rio skate, Rioraja agassizii (Müller & Henle, 1841) originally described based only on external morphological characters such as disc shape and position of the first dorsal fin. Those characters do not unequivocally distinguish this species from other skates, mainly from the genus Atlantoraja.
Varieties of Rioraja agassizii were reported by Garman (1913) and Miranda Ribeiro (1903, 1907) but only in the work of Menni (1972), Atlantoraja was proposed to allocate A. cyclophora, A. castelnaui and A. platana based on clasper features. According to recent studies, Rioraja differs from Atlantoraja species by having the distance from the origin of first dorsal fin to the end of caudal fin equal to or greater than the distance of the origin of first dorsal fin to the origin of the posterior lobe of pelvic fin, a nasal flap falciform, one interdorsal thorn, ventral surface of the disc lighter with few small dark spots, apex of snout with a dark blotch ventrally, triangular dorsal terminal 1 cartilage on clasper, and caudal fin absent (Menni, 1973; Barbosa, Gomes, 1998; Gomes et al., 2010, 2019; Moreira et al., 2017). However, individuals of R. agassizii have already been misidentified with A. platana by the presence of dark spots on pectoral fins (Barbosa, Gomes, 1998) and little is known on intraspecific variation in color pattern in these species. Descriptions of some anatomical components of Rioraja agassizii are found in the literature, such as the cervicothoracic synarcual cartilage (Gomes et al., 1997), mucous pores of the ampullae of Lorenzini (Gomes, Paragó, 2005), egg capsules (Oddone et al., 2008), claspers (Moreira et al., 2017) and scapulocoracoid (Soares et al., 2020). Characters from squamation and neurocranium morphology (i.e., alar thorns strongly reclined laterally and preorbital process continuous with supraorbital crest; McEachran, Dunn, 1998) were proposed to diagnose the tribe Riorajini but detailed accounts on these structures and their variation within Rioraja are scarce.
Here, we redescribe Rioraja agassizii, providing detailed descriptions and illustrations of external morphology, neurocranium, visceral skeleton and pelvic girdle. Notes on sexual dimorphism and ontogenetic differences in coloration, body and cranial measurements, and dermal denticles are also presented.
Material and methods
A total of 105 specimens of Rioraja agassizii were examined in this study; 84 were obtained from the commercial bottom fishing (continental shelf) from Southeastern and Southern Brazil (from Rio de Janeiro to Rio Grande do Sul states) and 21 corresponded to museum specimens. The material examined was fixed in formaldehyde 7% and preserved in ethanol 70%. In addition, 32 specimens of Atlantoraja species were examined as comparative material. No licenses were needed for the realization of this study.
Museum abbreviations are as follow: Brazil: Chondrichthyes Anatomic Collection from Universidade do Estado do Rio de Janeiro, Rio de Janeiro (AC.UERJ); Fundação Universidade do Rio Grande, Rio Grande (FURG); Museu de Ciências e Tecnologia da Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre (MCP); Museu Nacional do Rio de Janeiro, Rio de Janeiro (MNRJ); Museu de Zoologia, Universidade de São Paulo, São Paulo (MZUSP); Universidade do Estado do Rio de Janeiro, Rio de Janeiro (UERJ); Universidade Santa Úrsula, Rio de Janeiro (USU); Argentina: Museu Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires (MACN); France: Museum National d’Histoire Naturelle, Paris (MNHN). A list of the material examined is provided below. The abbreviations DW and TL used throughout the text refer to disc width and total length, respectively. Information on sex of each specimen is also presented.
Specimens were manually dissected with the aid of scissors, scalpels and forceps, sometimes placed into hot water (around 60 ºC) in a solution of potassium carbonate (K2CO3) to facilitate the removal of skin and muscles. Of the 105 specimens examined, 38 were measured and 64 dissected for the examination of the neurocranium, visceral arches, pelvic girdle and fin. Maturity stages of specimens were determined following the works of Oddone, Vooren (2008) and Oddone et al. (2007a,b). Photographs of external morphology and skeletal components were taken with a digital camera (Canon Power Shot SX610 HS), digitized and edited with the aid of Adobe Photoshop CS6. Drawings of skeletal structures were made in the Adobe Illustrator CC and edited in Adobe Photoshop CS6. Measurements and counts follow Hubbs, Ishiyama (1968), McEachran, Compagno (1979) and Leible (1988). Descriptions of the neurocranium and pelvic girdle were based in Hulley (1972), Leible (1988) and Nishida (1990); visceral skeleton follows Gillis et al. (2009); dermal denticles, Leible (1988) and Cappetta (2012).
Thorns are defined as derivative dermal denticles varying from 2 mm to 10 mm with a star-shaped or circular base and acute tip (Leible, 1988) and named according to their location (Fig. 1): rostral (from the anterior tip of rostrum to the orbital region), preorbital (close to the anterior margin of orbits), interorbital (on the interspace between orbits), midorbital (close to the medial margin of orbits), postorbital (on the posterior margin of orbits and delimited by the spiracle), spiracular (on the internal margin of spiracle), interspiracular (on the interspace between spiracles), nuchal (between interspiracular and scapular thorns), scapular (on the scapular region), middorsal (on the midline posterior to scapulocoracoid and anterior to the pelvic girdle), mediocaudal (on the midline of caudal dorsal surface), interdorsal (between dorsal fins), and alar (curved tips and on the lateral tip of pectoral fins of adult males). Prickles are defined as diminute dermal denticles, smaller than 1 mm with narrow base and acute edges, randomly distributed around the body.
Results
Rioraja Whitley, 1929
Rioraja agassizii (Müller & Henle, 1841)
(Figs. 2–9; Tabs. 1–3)
Uraptera agassizii Müller, Henle, 1841:155–156 (original description; type locality: “Brazil”). —Garman, 1913:367–368, figs. 53, 68 (systematic account). —Marini, 1929:452 (listed).
Raja agassizii Günther, 1870:465 (British Museum; catalogue). —Miranda Ribeiro, 1923:34 (Rio de Janeiro, Brazil; catalogue). —Ringuelet, Aramburu, 1960:46 (Argentina; catalogue). —Gomes et al., 1997:96 (cervicothoracic synarcual cartilage). —Gadig, 1998:50 (listed). —Andreata et al., 2002:1142 (listed).

FIGURE 1| Terminology of thorns. alt, alar; idt, interdorsal; iot, interorbital; ist, interespiracular; mct, mediocaudal; mdt, middorsal; mot, midorbital; nut, nuchal; pet, preorbital; pot, postorbital; rst, rostral; sct, scapular; spt, spiracular.
Raja agassizii Miranda Ribeiro, 1961:4 (Rio de Janeiro; catalogue). —Soares et al., 1999:201 (São Paulo, Brazil; diet). —Estalles et al., 2009:60 (Argentina; reproductive biology). —Muto et al., 2001:217 (São Paulo, Brazil; diet). —Costa, Chaves, 2006:1 (Paraná and Santa Catarina, Brazil; listed).
Raja (Rioraja) agassizii Menni, 1973:419, 422, fig. 2 (Argentina; catalogue). —Roux, 1977:118 (Brazil and Argentina; listed). —Menni et al., 1984:63 (Argentina; catalogue). —Andreata, Séret, 1995:581 (Espírito Santo, Brazil; listed). —Deynat, Séret, 1996:81 (squamation). —Barbosa, Gomes, 1998:127 (juvenile external morphology).
Rioraja agassizii Whitley, 1929:254 (original description; type locality: Rio de Janeiro, Brazil). —McEachran, Dunn, 1998:286 (systematic study). —Compagno, 1999:489 (listed). —Mazzoleni, Schwingel, 1999:63 (Itajaí, Santa Catarina, Brazil; listed). —Menni, Stehmann, 2000:88 (Argentina, Uruguay and Brazil; listed). —Gomes, Gadig, 2003:29 (São Paulo, Brazil; listed). —McEachran, Aschliman, 2004:101 (systematic study). —Compagno, 2005:533 (listed). —Gomes, Paragó, 2005:55–62, figs. 1, 5, 9 (mucous pores canals). —Oddone et al., 2006:43 (Rio Grande do Sul, Brazil; reproductive biology). —Colonello et al., 2007:277 (Uruguay and Argentina; reproductive biology). —Cousseau et al., 2007:14 (Mar del Plata, Argentina; catalogue). —Ebert, Compagno, 2007:116 (listed). —Oddone et al., 2007a:415 (reproductive biology). —Oddone et al., 2007b:594 (reproductive biology). —Oddone, Vooren, 2008:5 (egg capsules). —Gomes et al., 2010:166–167, fig. 263 (Rio de Janeiro, Brazil; catalogue). —Last et al., 2016:944 (listed). —Moreira et al., 2017:1–12 (clasper morphology). —Gomes et al., 2019:297, fig. 286 (Rio de Janeiro, Brazil; catalogue). —Coelho et al., 2020:1–16 (Brazil; ecological niche model). —Soares et al., 2020:493–500 (scapulocoracoid morphology).
Raja agassizii meta (in part) Miranda-Ribeiro, 1953:406 (type catalogue, Museu Nacional, Rio de Janeiro, Brazil).
Raja agassizii picta Miranda Ribeiro, 1953:406 (type catalogue, Museu Nacional, Rio de Janeiro, Brazil).
Raja platana Miranda Ribeiro, 1961:4 (Toku Maru; listed).
Lectotype. MNHN 2430, male, 405 mm TL, 259 mm DW (Brazil) [designated herein].
Diagnosis. Rioraja agassizii distinguishes from Atlantoraja species by lacking caudal fin (vs. present in Atlantoraja spp.); posterior margin of anterior nasal flaps straight (vs. rounded); one interdorsal thorn (vs. none); ventral surface of the disc cream in color with few small dark spots (vs. dark ventral surface); dark blotch on the ventral surface of snout (vs. absent); mucous pore canals less numerous and rounded (vs. more numerous and elongated); distance between the first dorsal fin and posterior lobe of pelvic fin somewhat similar to the distance between the first dorsal fin and caudal tip (vs. first dorsal fin closer to the caudal tip in Atlantoraja castelnaui); clasper with a dorsal terminal 1 cartilage triangular and with a conspicuous proximal shelf (vs. inverted U-shape and shelf absent); shape of the ventral terminal cartilage like an elongated spoon (vs. Y-shape in A. cyclophora, flattened in A. platana and wrench-shaped in A. castelnaui).

FIGURE 2| A. Drawing of Rioraja agassizii provided by Müller, Henle (1841). B. Dorsal view of MNHN 2430, male, 405 mm TL (lectotype designated herein and modified from MNHN database). Scale bar = 20 mm.
External description. Proportional dimensions of males and females are presented in Tab. 1. Disc rhomboidal with straight or sinuous pectoral anterior margins (in adult males) according to the degree of maturity and posterior pectoral margins slightly convex; 1.2–1.3 times wider than long (Figs. 2–3). Snout long and obtuse, slightly projecting beyond anterior disc margin and 0.2 times the disc length (Figs. 2–4). Posterior lobe of disc covering the anterior portion of the anterior lobe of pelvic fin. Preorbital length 1.2–1.6 times greater than prenasal length and 1.0–1.2 times the preoral length.
Eye and spiracle closely set; spiracles projecting obliquely from midline (Fig. 2B, 3). Eye length greater than spiracle length and goes 1.3–2.5 in times the interorbital distance; adult males with slightly greater eyes than females and juveniles.
Anterior nasal flap broad, not fused medially, about four times the lateral flap and covering the mouth commissure (Fig. 4); lateral margin slightly sigmoid with an anterior notch close to the nasal aperture, posterior margin straight and smooth, and inner margin nearly straight. Lateral flap tube-like, not fringed and outlining the incurrent aperture.
Mouth shape strongly arched and displaying a V-shape in males (Fig. 4). Distance between incurrent apertures 0.9–1.2 times the distance between mouth corners. Teeth with a flat oval base and acute single cusp. Total tooth count in upper jaw 39–51 arranged in four rows and in lower jaw 36–48 arranged in five rows; higher tooth counts in adult females than in males (Tab. 2).
Gill openings slitlike and medially situated to the propterygia; first one 1.5–1.9 times greater than fifth and third slightly larger than the first (Fig. 4). Distance between first gill slits 1.7–2.2 times the distance between fifth gill slits (Tab. 1); females presenting the last gill slits more widely spaced (6.4–8.3% TL) than males (5.4–6.0% TL).
TABLE 1 | Body measurements of males and females of Rioraja agassizii. Total length (TL) in millimeters and other measurements as percentages of total length (% TL).
Measurements | Males (n = 13) | Females (n = 25) | ||
Range | Mean | Range | Mean | |
Total length (mm) | 355–470 | 418.0 | 364–561 | 471.0 |
Disc width | 58.0–68.0 | 64.3 | 55.9–70.3 | 62.9 |
Disc length | 48.0–55.7 | 51.2 | 44.2–60.4 | 55.6 |
Prenasal length | 8.0–11.4 | 9.7 | 9.2–13.8 | 10.9 |
Preoral length | 10.7–13.7 | 12.9 | 11.6–15.7 | 13.5 |
Preorbital length | 12.6–15.1 | 12.7 | 12.8–15.2 | 13.7 |
Eye length | 2.4–4.8 | 3.3 | 1.8–3.2 | 2.5 |
Interorbital distance | 4.5–6.4 | 5.9 | 4.6–7.7 | 6.0 |
Eye-spiracle distance | 0.6–0.9 | 0.7 | 0.5–0.9 | 0.7 |
Spiracle diameter | 2.3–2.8 | 2.5 | 1.8–3.0 | 2.7 |
Interespiracular distance | 5.1–6.0 | 5.6 | 5.4–6.2 | 5.8 |
Anterior nasal flap length | 3.1–3.6 | 3.4 | 2.6–4.3 | 3.5 |
Internarial distance | 4.5–7.1 | 6.0 | 5.4–7.8 | 6.5 |
Mouth width | 5.5–7.6 | 6.2 | 5.8–8.4 | 7.2 |
First gill width | 1.2–1.8 | 1.2 | 1.4–2.3 | 1.7 |
Third gill width | 1.2–1.9 | 1.4 | 1.1–2.8 | 1.9 |
Fifth gill width | 0.8–1.3 | 1.1 | 0.9–1.2 | 1.0 |
Distance between first gill openings | 11.6–13.1 | 11.9 | 12.1–14.6 | 12.5 |
Distance between fifth gill openings | 5.4–6.0 | 5.7 | 6.4–8.3 | 6.6 |
Pelvic anterior lobe length | 10.2–11.6 | 10.9 | 9.1–12.3 | 10.9 |
Pelvic posterior lobe length | 9.5–16.0 | 12.8 | 10.2–12.7 | 11.8 |
Clasper outer length | 12.7–15.2 | 14.1 | – | – |
Clasper inner length | 20.7–23.2 | 21.5 | – | – |
Clasper base length | 2.5–3.3 | 2.8 | – | – |
Tail width at axil of pelvics | 3.7–4.0 | 3.8 | 2.9–4.1 | 3.4 |
Snout-cloaca length | 43.9–48.2 | 46.1 | 41.2–50.4 | 45.5 |
Cloaca-caudal length | 51.5–54.0 | 52.3 | 47.8–54.1 | 51.0 |
Distance between pelvic and first dorsal fins | 24.5–29.6 | 26.2 | 22.1–27.5 | 24.9 |
Distance between first dorsal and caudal fins | 19.6–25.2 | 23.1 | 15.9–24.3 | 22.6 |
Interdorsal distance | 3.9–6.8 | 5.4 | 3.0–4.3 | 3.5 |
First dorsal fin base | 3.2–3.9 | 3.6 | 3.2–4.0 | 3.5 |
First dorsal fin height | 2.3–3.0 | 2.5 | 2.2–2.8 | 2.4 |
Second dorsal fin base | 3.2–4.2 | 3.5 | 2.5–4.4 | 3.9 |
Second dorsal fin height | 2.0–2.5 | 2.2 | 1.8–2.4 | 2.2 |
Pelvic fin deeply concave outwardly with anterior and posterior portions continuous externally but forming distinct lobes (Fig. 3). Anterior lobe similar in size to the posterior one in juveniles and females; 1.4 times posterior lobe in adult males. Anterior margin of the anterior lobe nearly straight close to its origin and convex distally. Origin of anterior lobe slightly anterior and ventral to pectoral fin insertion. Distal tips of radials well-marked and prominent along the posterior margin of anterior lobe. Posterior lobe with strongly convex lateral margin and straight inner margin. Clasper inner length 1.4–2.2 times pelvic posterior lobe length and 1.5–1.6 times clasper outer length.

FIGURE 3| External morphology and color pattern of Rioraja agassizii. A. dorsal view and B. ventral view of MZUSP (uncatalogued), 424 mm TL; C. dorsal view and D. ventral view of UERJ 1569, female, 466 mm TL; E. MZUSP 117280, female, 543 mm TL; F. MNRJ 50512, female, 505 mm TL. Scale bars = 10 cm.
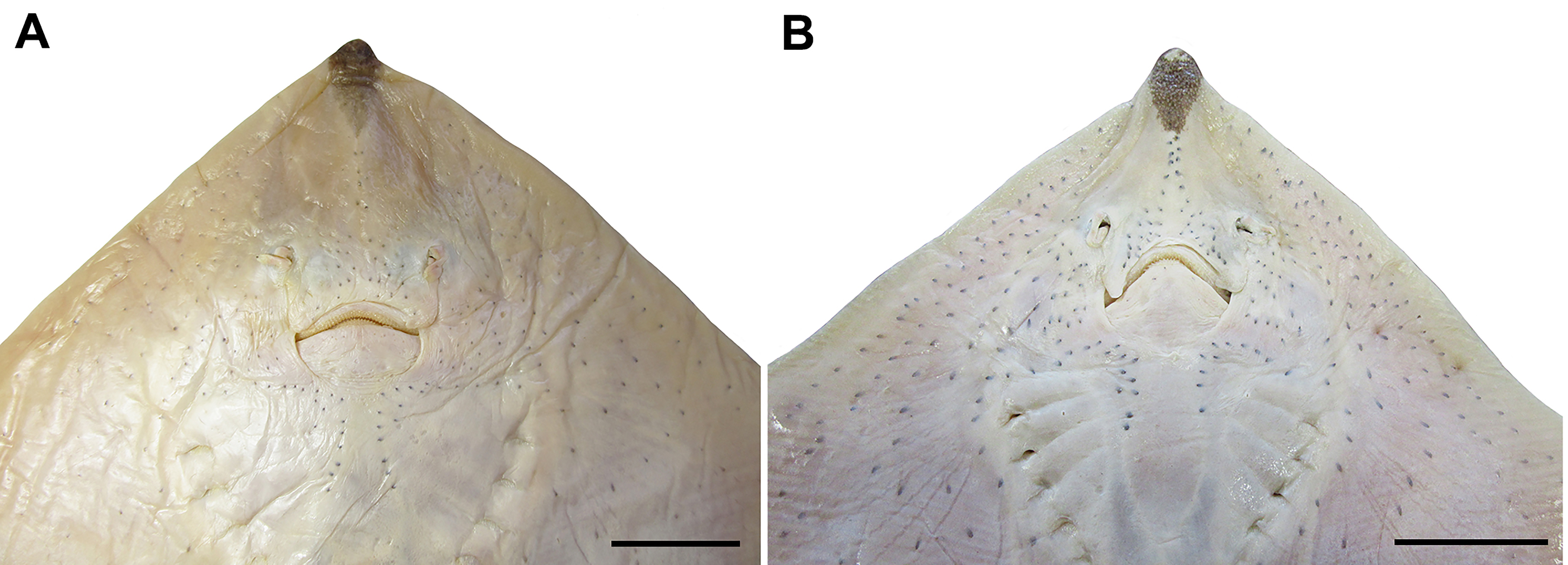
FIGURE 4| Ventral view of the head of Rioraja agassizii. A. MZUSP 117280, female, 543 mm TL. B. USP (uncatalogued), male. Scale bars = 20 mm.
Dorsal fins rounded, interdorsal space corresponding to 3.0–6.8% of total length, slightly greater in adult males than in adult females (Tab. 1). First dorsal fin 1.1–1.2 times higher than the second. Distance from axil of the pelvic fin to first dorsal fin origin 1.2–1.4 times the distance from first dorsal fin origin to caudal tip.
Caudal region slender and elongate, clearly distinct from disc and tapering from pelvic base to its distal tip. Caudal region greater than the disc; distance from cloaca to tail distal tip 1.1–1.2 times the distance from the snout to cloaca. Caudal fin absent.
Squamation. Skin in juveniles smooth on dorsal and ventral surfaces; prickles and thorns more conspicuous in adult specimens. Preorbital, interorbital and midorbital thorns absent in all examined specimens. Mediocaudal and interdorsal thorns more abundant in adult females than in adult males and juveniles of both sexes (Tab. 2). Rostral, postorbital, spiracular, interspiracular, nuchal and scapular thorns in adult females, absent in adult males and juveniles. Alar thorns in adult males varying from 72 to 76 arranged in three horizontal rows (8–23).
Coloration. Color pattern light brown dorsally and whitish ventrally with a dark spot on the anterior tip of snout (Fig. 3). Diffuse dark ocellus with a paler center on the medial portion of pectoral fins in most specimens (Figs. 3A, E, F). Dorsal surface with light and diffuse and scattered spots mostly in females (Fig. 3C). Dark saddles on the caudal region in juveniles, absent in adult specimens. Ventral surface of disc with sensory and mucous pores as black streaks and spots.
Neurocranium. Largest width at nasal capsules, 1.0–1.1 times in the nasobasal length (Tab. 3). Rostrum delimitated posteriorly by the intersection between nasal capsules and rostral cartilages at ophthalmic foramen and extends anteriorly to the tip of rostral appendix (Figs. 5–6). Rostral appendix fused to the rostral cartilage, wider at the rostral node, tapering posteriorly, its length 0.9–1.6 times in rostral cartilage length and 2.4–2.8 times rostral appendix width. Rostral cartilage not flexible, large dorsally from its base to ophthalmic foramina, narrowing towards its anterior tip, its length 1.0–1.4 times in nasobasal length. Furrow present on dorsal surface of rostral cartilage, extending from the anterior region to the anterior margin of anterior fontanelle, extending throughout rostral cartilage in lateral view.
TABLE 2 | Meristic counts of males and females, juveniles and adult specimens of Rioraja agassizii.
Counts | Males | Females | ||
Juveniles | Adults | Juveniles | Adults | |
Mediocaudal thorns | 7–10 | 10–27 | 8–13 | 23–31 |
Interdorsal thorns | 1–4 | 1 | 1–4 | 1–4 |
Rostral thorns | – | – | – | 1–3 |
Postorbital thorns | – | – | – | 1–2 |
Spiracular thorns | – | – | – | 1–2 |
Interspiracular thorns | – | – | – | 1 |
Nuchal thorns | – | – | – | 4–5 |
Scapular thorns | – | – | – | 2–3 |
Alar thorns | – | 72–76 | – | – |
Upper tooth counts | – | 40–45 | – | 39–51 |
Lower tooth counts | – | 37–42 | – | 36–48 |

FIGURE 5| Neurocranium of Rioraja agassizii, MZUSP 117280, female, 543 mm TL. A. dorsal, B. ventral views. af, anterior fontanelle; bp, basal plate; epb, epiphysial bridge; end, endolymphatic foramen; icf, internal carotid artery foramen; inc, inner nasal cartilage; ins, internasal space; ja, jugal arch; na, nasal aperture; nc, nasal capsule; oc, otic capsule; occ, occipital condyle; onc, outer nasal cartilage; opf, ophthalmic foramen; peri, perilymphatic foramen; pf, posterior fontanelle; pnc, posterior nasal cartilage; pos, postorbital process; pre, preorbital process; ptp, pterotic process; ra, rostral appendix; rb, rostral base; rn, rostral node; rs, rostrum. Scale bar = 10 mm.
TABLE 3 | Neurocranial measurements as percentages of neurocranium total length (% NTL).
Measurements | Males (n = 26) | Females (n = 36) | ||
Range | Mean | Range | Mean | |
Nasobasal length | 52.0–72.4 | 55.8 | 48.3–80.3 | 56.2 |
Rostral length | 39.1–49.5 | 45.7 | 24.6–52.3 | 45.7 |
Rostral appendix length | 47.5–67.6 | 56.3 | 43.6–54.8 | 49.8 |
Rostral appendix width | 28.2–55.0 | 45.1 | 28.6–57.7 | 43.4 |
Rostral base width | 7.9–17.2 | 14.3 | 10.2–20.4 | 14.9 |
Width across nasal capsules | 46.2–60.4 | 50.2 | 44.2–65.6 | 51.3 |
Nasal aperture width | 10.3–22.4 | 17.8 | 15.4–23.0 | 18.4 |
Internasal distance | 8.0–14.6 | 10.3 | 7.9–15.2 | 11.1 |
Interorbital distance | 13.2–25.3 | 18.4 | 16.2–24.6 | 18.9 |
Anterior fontanelle length | 17.1–31.0 | 23.5 | 21.2–34.4 | 25.5 |
Anterior fontanelle width | 15.6–22.1 | 17.6 | 14.3–16.7 | 15.5 |
Epiphyseal bridge width | 2.1–4.5 | 2.4 | 2.8–4.1 | 2.9 |
Posterior fontanelle length | 21.3–32.8 | 24.9 | 19.8–34.4 | 24.4 |
Posterior fontanelle width | 5.1–10.3 | 7.1 | 5.8–11.1 | 7.6 |
Basal plate width | 10.7–26.0 | 13.7 | 10.2–18.5 | 13.8 |
Width across otic capsules | 22.0–31.0 | 25.0 | 17.1–36.1 | 25.3 |
Nasal capsule obliquely oriented to the cranial roof and perpendicular to the rostrum, with anterior edge slightly anterior to the rostral cartilage base (Figs. 5A, 6A). Foramen of ethmoidal nerve on anterodorsal surface of the nasal capsule, and lateral to the foramen lies the antorbital facet which articulates to the antorbital cartilage. Nasal aperture wide with a rounded anterior margin and nearly straight posterior margin; ophtalmicus foramen anteriorly and oronasal canal more posteriorly. Internasal space 4.0–4.4 times in the greatest cranium width. Outer nasal cartilage triangular, dividing incurrent and excurrent apertures laterally oriented (Figs. 5B, 6B). Inner nasal cartilage rounded and laterally expanded, its length two-thirds of its width, it lies on posterior margin of nasal apertures, extending from the internasal septum to the lateral margin of nasal fontanelle. Posterior nasal cartilage broad anteriorly, tapering posteriorly, curved distally, oblique to anteroposterior axis, its posterior tip more laterally situated than the anterior one (Figs. 5B, 6B).
Cranial roof extends from the posterior margin of the anterior fontanelle to the parietal fossa, slightly posterior to the posterior fontanelle and limited laterally by the supraorbital crest (Figs. 5A, C). Two dorsal fontanelles; anterior fontanelle wider than the posterior, teardrop-shaped beginning at posterior third of rostral cartilage, extending to anterior edge of supraorbital crest (Figs. 5A, 6A), 1.4–3 times longer than wide and relatively wider in males than in females. Posterior fontanelle almost the same length of the anterior one, extending from the epiphysial bridge to the parietal fossa. Epiphysial bridge slender with a vertical extension on its median portion; bridge corresponding to 16–21.6 times in nasobasal length. Preorbital process well-developed on the posterior third of nasal capsule and continuous to the supraorbital crest.
Basal plate begins anteriorly at the shallow ethmoidal fossa right after the nasal capsules, runs to the posteroventral end of the optic region and ends at the ventral margin of the foramen magnum (Figs. 5B, 6B). A single foramen for the internal carotid artery is situated on the posterior third at the midline. Basal plate narrower than cranial roof, its width 5.4–9.4 times in nasobasal length (Tab. 3).
Orbital region delimited anteriorly by the posterior margin of the nasal capsules, posteriorly by the anterior margin of otic capsules, dorsally by the cranial roof, ventrally by the basal plate, and medially by the orbital wall, which is perforated by several foramina to the passage of cranial nerves and blood vessels (Fig. 6C). Oronasal canal ventrally close to the basal plate. Foramina for the anterior cerebral vein on the same vertical line between the oronasal canal and the foramina for the ophtalmicus nerve. Large foramen for the optic nerve (II) posterior to the foramina for the anterior cerebral vein and ventral to a single foramen for the trochlear nerve (IV). Foramen for the oculomotor nerve (III) posterior to the optic nerve and anterior and dorsal to optic pedicel. Foramen for the afferent pseudobranchial artery ventral to the optic pedicel and close to the basal plate. Prootic foramen for the passage of the trigeminus and facialis nerves (V+VII) as the second largest aperture of the orbital wall and slightly posterior to the foramen for the abducens nerve (VI). Prootic foramen separated posteriorly from the hyomandibular ramus of the facialis nerve by the prefacial commissure, close to the otic region (Fig. 6C).
Otic region as the second widest region of the neurocranium (Figs. 5–6), composed of otic capsules, impressions of semicircular canals, parietal fossa and foramina for the endolymphatic (smaller aperture) and perilymphatic canals. Behind the sphenopterotic crest, there is a well-developed opistotic crest superior that runs dorsally to the otic capsule and ends close to the occipital region. The ventral portion of the otic region is delimited laterally by the opistotic crest inferior and the hyomandibular facet.

FIGURE 6| Neurocranium of Rioraja agassizii, MNRJ 50512, female, 505 mm TL. A. dorsal, B. ventral, C. lateral views. II, foramen for the optic nerve; III, foramen for the oculomotor nerve; IV, foramen for the trochlear nerve; VI, foramen for the abducens nerve; acvf, anterior canal vein foramen; af, anterior fontanelle; antf, antorbital facet; apba, foramen for the afferent pseudobranchial artery bp, basal plate; epb, epiphysial bridge; end, endolymphatic foramen; ethn, foramen for the passage of the ethmoidal nerve; fm, foramen magnum; hf, hyomandibular facet; hVII, foramen for the passage of the hyomandibular ramus of the facialis nerve; icf, internal carotid artery foramen; inc, inner nasal cartilage; ins, internasal space; ja, jugal arch; na, nasal aperture; nc, nasal capsule; oc, otic capsule; occ, occipital condyle; onc, outer nasal cartilage; opc, opistotic crest; opd, optic pedicel; opf, ophthalmic foramen; ornc oronasal canal; peri, perilymphatic foramen; pf, posterior fontanelle; pfc, prefacial commissure; pnc, posterior nasal cartilage; pos, postorbital process; pre, preorbital process; prf, parietal fossa; prof, prootic formamen; ptp, pterotic process; ra, rostral appendix; rb, rostral base; rn, rostral node; rs, rostrum; soc, supraorbital crest; sopf, superficial ophtalmic foramen.
Occiptal region on the posterior surface of the chondrocranium, composed by the large foramen magnum and the lateral edges of the hyomandibular facet. On each side of the foramen magnum lies the occipital condyle and lateral to it the foramen for the vagus nerve (X). Lateral to the foramen for the vagus nerve lies the foramen for the glossopharyngeal nerve (IX).
Visceral skeleton. Upper jaw concave medially; lower jaw strongly convex (Fig. 7A). Both jaws tapering towards midline. Hyomandibular cartilage tapering anteriorly. Dorsal (epi-) and ventral (cerato-) pseudohyoid rodlike, laterally to the basihyal cartilage (Figs. 7B–D). Basihyal cartilage wide, slender, strongly arched, with well-developed anterolateral projections. Hypobranchial 2 one-third the length of posterior hypobranchials, rounded and restricted to the medial portion of ceratobranchials 1 and 2. Hypobranchials 3 and 4 fused, not reaching the anterior portion of basibranchial copula. Ceratobranchials 1–4 overlapping medially; ceratobranchial 5 articulating directly to the basibranchial copula. Epibranchials 1 and 5 longer than medial ones. Pharyngobranchials 2 and 3 longer than the first; gill pickax half-length of the pharyngobranchial 1. Anterior processes of basibranchial copula straight, tapering anteriorly and shorter than the posterior portion of basibranchial copula. Cardiobranchial cartilage elongate; accessory cartilage of the basibranchial copula triangular and corresponding to one-fifth of the cardiobranchial cartilage.

FIGURE 7| Visceral arches of Rioraja agassizii, MNRJ 50512, female, 505 mm TL. A. jaws; B, D. dorsal and C, E. ventral views of hyoid and gill arches. abc, anterior portion of basibranchial copula; bbc, basibranchial copula; bh, basihyal; cb 1–5, ceratobranchials I–V; mck, Meckel’s cartilage; cph, cerato-pseudohyoid; eb 1–5, epibranchials I–V; eph, epi-pseudohyoid; hb 2–4, hypobranchials II–IV; lpbh, lateral projection of basihyal; pb 1–5, pharyngobranchials I–V; pq, palatoquadrate. Scale bar = 5 mm.
Pelvic girdle and fin. Pelvic bar transverse and nearly straight, with a process extending forward from the outer corner at each end (Fig. 8). Median ischio-pubic region and lateral iliac regions with obturatorial nerve foramina for diazonal nerves. Lateral prepelvic process triangular and directed anteriorly. Condyle for articulation ventrolateral, with enlarged anterior radial; obturator foramina posterior to the puboischiadic condyle. Pelvic anterior lobe with six radial segments; first radial segment stout, elongate, with very concave articular surface for puboischiadic condyle. Posterior lobe with 20 radial segments.
Geographic distribution. This species is endemic to the coastal waters of southwestern Atlantic occurring from Espírito Santo to Rio Grande do Sul, Brazil, Uruguay and Northern Argentina (Fig. 9; see also, Menni, 1973; Menni et al., 1984; McEachran, Miyake, 1990; Andreata, Serét, 1995; Gadig, 1998; Mazzoleni, Schwingel, 1999; Cousseau et al., 2000, 2007; Meneses, Paesch, 2003; Coelho et al., 2020). It has been reported in temperate waters (Menni, 1973; Bellisio et al., 1979; Menni et al., 1984; Menni, Stehmann, 2000) at depths up to 150 m (Andreata, Séret, 1995; Vooren, 1997; Cousseau et al., 2000, 2007) although rare over 120 meters depth (Oddone et al., 2007a).
Ecological notes. Largest female and male specimens recorded in Argentina, 710 mm and 629 mm in TL, respectively (Cousseau et al., 2007; Estalles et al., 2009). Females become sexually mature at 200 mm DW and the first sexual maturation takes with 250 mm DW, while males become mature at 200 mm DW (495–577 mm TL in females and 420–520 mm TL in males, Cousseau et al., 2007; 320 mm TL in Oddone et al., 2007a,b; 570 mm and 504 mm in TL for females and males respectively, Estalles et al., 2009). Ovulation, egg-laying and presence of sperm in the seminal vesicle all year round and at least one peak in the sexual activity (Oddone et al., 2007a,b).

FIGURE 8| Pelvic girdle and fin skeleton of Rioraja agassizii, MNRJ 50512, female, 505 mm TL. 1pvr, first pelvic radial segment; 1pvrc, condyle for the first pelvic radial segment; bpt, basipterygium; lpp, lateral pelvic process; obf, obturator foramen; pib, pubischiadic bar; pvr, pelvic radial segments. Scale bar = 10 mm.
Siqueira (1999) found that in specimens of Rioraja agassizii captured in Santos, São Paulo State, the egg-laying starts by the right uterus. Egg capsules bright and light brown, laterally keeled and symmetrically convex with a smooth surface. Mean total length of the egg capsules 47 mm and mean width 31 mm (Oddone, Amorim, 2006). Distance from anterior apron to apex of anterior horn and flange with less than 50% of the length of egg case with well-defined filaments and strongly adhered to the lateral keel. Lateral edge of 61 to 68 mm (Mabragaña et al., 2009).
Stomach contents have been reported to include crustaceans (Alpheidae, Amphipoda, Caridea, Copepoda, Cirripedia, Isopoda, Stomatopoda, Ogyrididae, Penaeidae, Solenoceridae, Sicyonidae), nematodes, polychaetes, and teleost fishes (Bothidae, Ophidiidae, Scorpedinidae, Serranidae, Bathracoididae, Ophichthidae, Bragmacerotidae) (Muto et al., 2001; Pasquino et al., 2011; Motta et al., 2016).
Common name. Rio skate.
Conservation status. This species is categorized as ‘Vulnerable’ (VU) (Pollom et al., 2020).
Remarks. Müller, Henle (1841) described the species Uraptera agassizii based on four specimens of which one is housed in Berlin (ZMB, not examined) and three in Paris (MNHN) but only one was found in alcohol and examined in this study (MNHN 2430, Fig. 2B).

FIGURE 9| Map showing the geographic distribution of Rioraja agassizii based on material examined in this study (red) and literature (blue).
Whitley (1929) designated the genus Rioraja in substitution of Uraptera because it was already pre-occupied by Uraptera Billberg, 1820, a genus of Lepidoptera (Insecta). However, subsequent authors used the species name Raja agassizi (e.g., Miranda Ribeiro, 1907, 1923; Bigelow, Schroeder, 1953; Ruschi, 1965; Menni, 1972b, 1973; Sadowsky, 1973; Figueiredo, 1977; Lucena, Lucena, 1981; Menni et al., 1984; Gomes et al., 1997; Barbosa, Gomes, 1998). Later, Menni (1973) synonymized the name Uraptera in Rioraja as proposed by Whitley (1929).
According to Roux (1977:118), a specimen of Rioraja agassizii was collected in Fernando de Noronha, which represents the northernmost record for the species. The specimen is deposited in the MNHN under the catalog number 1984-0019 (311 mm TL, 202 mm DW) and its identification was confirmed by the last author (ULG). Nevertheless, its catch locality remains uncertain and may have been mislabeled, since the species is only found in the continental waters of southwestern Atlantic.
Material examined. Argentina. MACN 3825, male, 316 mm TL, 222 mm DW, Puerto Quenquén. Brazil. Paraná: AC.UERJ 1105, male, 26°20’S 47°52’W. AC.UERJ 1106, male, 25°3’S 46°16’W. Rio de Janeiro: AC.UERJ 1360, male, 295 mm TL, Marambaia Restinga. AC. UERJ 1361, male, 310 mm TL, Marambaia Restinga. AC. UERJ 1362, male, 297 mm TL, Marambaia Restinga. AC.UERJ 1363, male, 267 mm TL, Marambaia Restinga. AC.UERJ 1367, male, 315 mm TL, Marambaia Restinga, Angra dos Reis. AC.UERJ 1380, female, 474 mm TL, 320 mm DW, Rio de Janeiro. AC.UERJ 1386, male, 440 mm TL, Rio de Janeiro. MNRJ 578, male, 367 mm TL, 230 mm DW, Farol de Santana. MNRJ 591, 5 females, 270–441 mm TL, 2 males, 311–381 mm TL, Rio de Janeiro. MNRJ 593, female, 450 mm TL, 282 mm DW, Farol de Santana. UERJ 695, female, 385 mm TL, 329 mm DW, Barra de Guaratiba. UERJ 741, female, 177 mm TL, 128 mm DW, Ilha Grande. UERJ 743, female, 270 mm TL, 168 mm DW, Ilha de Santana. UERJ 747, female, 277 mm TL, 190 mm DW, Ilha de Santana. UERJ 782, female, 430 mm TL, 313 mm DW, Ilha Rasa. UERJ 788, male, 306 mm TL, 182 mm DW. UERJ 943, 3, 90–149 mm TL, between Macaé and Niterói. UERJ 1178, 2 males, 360–436 mm TL, Macaé. UERJ 1179, female, 490 mm TL, 350 mm DW, Macaé. UERJ 1428, male, 188 mm TL, 116 mm DW, Ilha Grande. UERJ 1525, female, 294 mm TL, 190 mm DW, Maricá. UERJ 1569, female, 466 mm TL, 332 mm DW, Barra de Guaratiba. UERJ 1580, male, 532 mm TL, 345 mm DW, Barra de Guaratiba. UERJ 1581, female, 466 mm TL, 342 mm DW, Barra de Guaratiba. UERJ 1716, female, 153 mm TL, 95 mm DW, Ilha Grande. USU 119, 2 males, 318–335 mm TL, 2 females, 395–434 mm TL, Rio de Janeiro. USU 598, 4 females, 356–503 mm TL, Guaratiba. USU 2216, male, 495 mm TL, 419 mm DW. Rio Grande do Sul: MCP 7120, male, 260 mm TL, 159 mm DW, Torres. MCP 7122, male, 237 mm TL, 152 mm DW, Torres. MCP 7156, female, 586 mm TL, 381 mm DW, Torres. MCP 7158, female, 277 mm TL, 172 mm DW, Torres. MCP 7444, male, 421 mm TL, 263 mm DW, between Farol da Solidão and Farol da Conceição. UERJ 879, male, 356 mm TL, 239 mm DW, between Ilha do Bom Abrigo, São Paulo, and Paranaguá, Paraná. UERJ 880, male, 323 mm TL, 213 mm DW, between Ilha do Bom Abrigo, São Paulo, and Paranaguá, Paraná. UERJ 892.1, female, 90 mm TL, 60 mm DW, between Southern Ilha Bela, São Paulo, and Ilha Grande, Rio de Janeiro. UERJ 892.2, male, 172 mm TL, 102 mm DW, between Southern Ilha Bela, São Paulo, and Ilha Grande, Rio de Janeiro. Santa Catarina: AC.UERJ 1107, male, 27°20’S 48°2’W. MCP 1523, male, 318 mm TL, 205 mm DW, Florianópolis. MCP 4834, female, 360 mm TL, 262 mm DW, Ponta das Canas, Florianópolis. São Paulo: AC.UERJ 1044, male, 245 mm DW, Santos. AC.UERJ 1047, male, 355 mm TL, 240 mm DW, Santos. AC.UERJ 1048, male, 365 mm TL, 240 mm DW, Santos. AC.UERJ 1050, male, 360 mm TL, 245 mm DW, Santos. AC.UERJ 1051, male, 250 mm DW, Santos. AC.UERJ 1052, male, 390 mm TL, 265 mm DW, Santos. AC.UERJ 1053, male, 380 mm TL, 240 mm DW, Santos. AC.UERJ 1054, male, 230 mm DW, Santos. AC.UERJ 1055, male, 250 mm DW, Santos. AC.UERJ 1056, male, 240 mm DW, Santos. AC.UERJ 1057, male, 220 mm DW, Santos. AC.UERJ 1070, male, 390 mm TL, 255 mm DW, Santos. AC.UERJ 1071, male, 350 mm TL, 240 mm DW, Santos. AC.UERJ 1073, male, 395 mm TL, 250 mm DW, Santos. AC.UERJ 1074, male, 209 mm DW, Santos. AC.UERJ 1075, 231 mm DW, Santos. AC.UERJ 1102, male, Santos. AC.UERJ 1103, male, Santos, 24°11’S 46°29’W. AC.UERJ 1104, male, Santos, 24°39’S 46°17’W. AC.UERJ 1108, male, Santos 24°48’S 47°23’W. AC.UERJ 1109, male, Santos. AC.UERJ 1110, male, Santos. AC.UERJ 1111, male, Santos. AC.UERJ 1112, male, Santos. AC.UERJ 1113, male, Santos. AC.UERJ 1114, male, Santos. AC.UERJ 1115, male, Santos. AC.UERJ 1116, male, Santos. AC.UERJ 1117, male, Santos. AC.UERJ 1381, female, 567 mm TL, 365 mm DW, Santos. MNRJ 588, male, 356 mm TL, 226 mm DW. MNRJ 50512, female, 505 mm TL, 23º11’30”S 41º2’55”W. MZUSP (uncatalogued), male, 495 mm TL, 419 mm DW. MZUSP (uncatalogued), 3, 410–561 mm TL. MZUSP 117280, female, 543 mm TL. UERJ 830, male, 138 mm TL, 85 mm DW, Santos. UERJ 897, 4, 112–202 mm TL, Ilha do Trigo. UERJ 898, 5, 102–135 mm TL. UERJ 1719, female, 472 mm TL, 323 mm DW, Ubatuba.
Discussion
Differences between adult males and females of Rioraja were observed in mouth shape, distance between fifth gills slits, interdorsal space, occurrence and counts of thorns (Tabs. 1–2). Adult males of Rioraja present a mouth strongly arched and acute teeth vs. mouth slightly arched and rhomboid teeth in females and juveniles, as already described for other skates (e.g., Bigelow, Schroeder, 1953; Hulley, 1972; McEachran, 1977; Leible, 1988). No ontogenetic or sexual difference in counts of thorns was reported by Barbosa, Gomes (1998) who also described lower numbers of mediocaudal (9–13 vs. 7–31) and interdorsal thorns (1 vs. 1–4) in comparison to our observations. Oddone et al. (2007b) described up to five rows of alar thorns in adult males of Rioraja agassizii; differences from our observations could be explained by the maturity stage of the specimens examined in that study.
The distance between the last gill slits is wider in females than in males of Rioraja andmay be related to the width of scapulocoracoid (Soares et al., 2020). Sexual dimorphism has also been reported for total weight and liver size with higher values for adult females (Colonello et al., 2007; Estalles et al., 2009). Estalles et al. (2009) also observed a significant difference in disc width (larger in females than in males) but this was not tested statistically herein. No difference was observed in the cranial range measurements of males and females, except for slight differences in the anterior fontanelle width.
Menni (1973), Figueiredo (1977) and Menni et al. (1984) used the position of the first dorsal fin in relation to the posterior lobe of pelvic fin and caudal tip as a distinctive character between Rioraja and Atlantoraja. However, according to Barbosa, Gomes (1998) and our observations, this is not a reliable character since some specimens of A. cyclophora and A. platana display the origin of first dorsal somewhat equidistant to the pelvic fin and caudal tip as in Rioraja. Morphometrics of Riorajini species need to be further investigated in order to enlarge our knowledge on the intraspecific variation within genus and species of this tribe.
A taxonomic study and anatomical investigation of Atlantoraja species is in progress. We hope that morphological data presented here for Rioraja agassizii, mainly on neurocranium and visceral arches, can be potentially useful in future inferences of the interrelationships among Riorajini species and other rajiforms.
Comparative material examined. All specimens from Brazil. Atlantoraja castelnaui: AC.UERJ 747, female, 200 mm TL, 133 mm DW, no locality data; AC.UERJ 941, female, 713 mm TL, Guarujá, São Paulo; AC.UERJ 956, female, 850 mm TL, no locality data; UERJ 692, female, 672 mm TL, 480 mm DW, Copacabana Beach, Rio de Janeiro; UERJ 876, male, 247 mm TL, 161 mm DW, Guaratiba, Rio de Janeiro; UERJ 877.1, male, 243 mm TL, 165 mm DW, Guaratiba, Rio de Janeiro; UERJ 1686, female, 270 mm TL, 180 mm DW, Guarujá, São Paulo; UERJ 1628, male, 220 mm TL, 141 mm DW, Santos; UERJ 1629, female, 391 mm TL, 265 mm DW, Santos, São Paulo. Atlantoraja cyclophora: UERJ 378, female, 130 mm TL, 96 mm DW, São Paulo; UERJ 395, 4, 225–247 mm TL, Itajaí, Santa Catarina; UERJ 396, male, 325 mm TL, 257 mm DW, Itajaí, Santa Catarina; UERJ 399, female, 517 mm TL, 389 mm DW, Itajaí, Santa Catarina; UERJ 740, male, 231 mm TL, 176 mm DW, Maricá, Rio de Janeiro; UERJ 790, female, 106 mm TL, 71 mm DW, Ilha Rasa, Rio de Janeiro; UERJ 830, female, 359 mm TL, 270 mm DW, Itajaí, Santa Catarina; UERJ 890, female, 121 mm TL, 91 mm DW, Saquarema, Rio de Janeiro; UERJ 894, 4, 115–216 mm TL, Guaratiba, Rio de Janeiro; UERJ 1625, male, 156 mm TL, 110 mm DW, Santos, São Paulo. Atlantoraja platana: UERJ 1502, 2, 162–241 mm TL, Niterói, Rio de Janeiro; UERJ 1786, female, 763 mm TL, 623 mm DW, Rio Grande, Rio Grande do Sul; UERJ 1788, female, 740 mm TL, 606 mm DW, Rio Grande, Rio Grande do Sul; UERJ 1849, female, 739 mm TL, 577 mm DW, Santos, São Paulo; UERJ 1850, male, 666 mm TL, 525 mm DW, Santos, São Paulo; UERJ 2162, female, 530 mm TL, 440 mm DW, Santos, São Paulo.
Acknowledgments
The authors wish to thank Calorus Vooren (in memorian) and Maria Cristina Oddone (FURG), Gustavo Chiaramonte (MACN), Carlos Lucena and Margarete Lucena (MCP), Paulo Buckup (MNRJ), Bernard Séret (MNHN) and José Lima Figueiredo (MZUSP) for permission to examine specimens under their care. To Hugo Santos for all the help and valuable comments for this study. The first author was supported by CAPES (Finance code 001).
References
Andreata JV, Meurer BC, Baptista MGS, Manzano FV, Teixeira DE, Longo MM, Freret N. Composição da assembléia de peixes da Baía da Ribeira, Angra dos Reis, Rio de Janeiro, Brasil. Rev Bras Zool. 2002; 19(4):1139–46. http://dx.doi.org/10.1590/S0101-81752002000400019
Andreata JV, Séret B. Relação dos peixes coletados nos limites da plataforma continental e nas montanhas submarinas Vitória, Trindade e Martin Vaz, durante a campanha oceanográfica MD-55 Brasil. Rev Bras Zool. 1995; 12(3):579–94. http://dx.doi.org/10.1590/S0101-81751995000300014
Barbosa FS, Gomes UL. Morfologia juvenil de quatro espécies do gênero Raja (Linnaeus, 1958) das regiões sudeste e sul do Brasil (Chondrichthyes, Batoidei, Rajidae). Biociências. 1998; 6(2):125–46.
Bellisio NB, Lopez RB, Torno A. Peces marinos patagónicos. Buenos Aires: Codex; 1979.
Bigelow HB, Schroeder WC. Fishes of the Western North Atlantic. New Haven: Sears Foundation for Marine Research, Yale University; 1953. https://doi.org/10.5962/bhl.title.7464
Cappetta H. Vol, 3E: Chondrichthyes, Mesozoic and Cenozoic Elasmobranchii: Teeth. In: Schultze, H-P, editor. Handbook of Paleoichthyology: München: Verlag Dr. Friedrich Pfeil; 2012.
Coelho JFR, Lima SMQ, Petean FF. Phylogenetic conservatism of abiotic niche in sympatric Southwestern Atlantic skates. Mar Biol Res. 2020; 16(6–7):458–73. https://doi.org/10.1080/17451000.2020.1837883
Compagno LJV. Checklist of living elasmobranchs. In: Hamlett WC, editor. Sharks, skates, and rays, the biology of elasmobranch fishes. Baltimore: John Hopkins University Press; 1999. p.471–98.
Compagno LJV. Checklist of living Chondrichthyes. In: Hamlett WC, editor. Reproductive biology and phylogeny of Chondrichthyes: sharks, rays and chimaeras, vol. 3. Endfield: Science Publishers; 2005. p.503–48.
Colonello JH, Garcia ML, Lasta CA. Reproductive biology of Rioraja agassizii from the coastal southwestern Atlantic ecosystem between northern Uruguay (34º S) and northern Argentina (42º S). In: Ebert DA, Sulikowski JA, editors. Biology of skates. Dordrecht: Developments in Environmental Biology of Fishes. 2007. p.171–78. https://doi.org/10.1007/978-1-4020-9703-4_12
Costa L, Chaves PTC. Elasmobrânquios capturados pela pesca artesanal na costa sul do Paraná e norte de Santa Catarina, Brasil. Biota Neotrop. 2006; 6(3):BN0270632006. https://doi.org/10.1590/S1676-06032006000300007
Cousseau MB, Figueroa DE, Díaz de Astarloa JM. Clave de identificacion de las rayas del litoral marítimo de Argentina y Uruguay (Chondrichthyes, Familia Rajidae). Mar del Plata: INIDEP, Secretaria de Agricultura, Ganadería, Pesca y Alimentación; 2000. Available from: https://aquadocs.org/handle/1834/2505
Cousseau MB, Figueroa DE, Díaz de Astarloa JM, Mabragaña E, Lucifora LO. Rayas, chuchos y otros batoideos del Atlántico Sudoccidental (34º S–55º S). Mar del Plata: INIDEP, Secretaria de Agricultura, Ganadería, Pesca y Alimentos; 2007. Available from: http://aquaticecology.weebly.com/uploads/8/1/1/9/81193470/batoideos_del_atl%C3%A1ntico_sw_parte_1.pdf
Deynàt PP, Séret B. Le revêtement cutané des raies (Chondrichthyes, Elasmobranchii, Batoidea). I: Morphologie et arrangement des denticules cutanés. Ann Sci Nat, Zool Biol Paris. 1996; 17(2):65–83.
Ebert DA, Compagno LJV. Biodiversity and systematics of skates (Chondrichthyes: Rajiformes: Rajoidei). In: Ebert DA, Sulikowski JA, editors. Biology of skates. Dordrecht: Developments in Environmental Biology of Fishes. 2007. p.5–18. https://doi.org/10.1007/978-1-4020-9703-4_2
Estalles M, Perez Comesaña JE, Tamini LL, Chiaramonte GE. Reproductive biology of the skate, Rioraja agassizii (Müller and Henle, 1841), off Puerto Quequén, Argentina. J Appl Ichthyol. 2009; 25(S1):60–65. https://doi.org/10.1111/j.1439-0426.2008.01103.x
Evermann BW, Kendall WC. Notes on a collection of fishes from Argentina, South America, with descriptions of three new species. Proc U S Natl Mus. 1907; 31(1482):67–108. https://doi.org/10.5479/si.00963801.31-1482.67
Figueiredo JL. Manual de peixes do sudeste do Brasil: I. Introdução, cações, raias e quimeras. São Paulo: Museu de Zoologia, Universidade de São Paulo; 1977. https://doi.org/10.5962/bhl.title.109986
Gadig OBF. Peixes cartilaginosos da costa do Estado de São Paulo. Ceciliana. 1998; 8(9):41–51.
Garman S. The Plagiostomia (sharks, skates, and rays). Mem Mus Comp Zool. 1913; 36:1–515. https://doi.org/10.5962/bhl.title.43732
Gillis JA, Dahn RD, Shubin NH. Chondrogenesis and homology of the visceral skeleton in the little skate, Leucoraja erinacea (Chondrichthyes: Batoidea). J Morphol. 2009; 270(5):628–43. https://doi.org/10.1002/jmor.10710
Gomes UL, Gadig OBF. Família Rajidae. In: Menezes NA, Buckup PA, Figueiredo JL, Moura RL, editors. Catálogo das espécies de peixes marinhos do Brasil. São Paulo: Museu de Zoologia de São Paulo; 2003. p.28–30.
Gomes UL, Paragó C. A utilização da distribuição de poros de canais de muco e da coloração ventral como caracteres taxonômicos em Riorajini (Chondrichthyes, Batoidea, Rajidae). Biociências. 2005; 13(1):55–62.
Gomes UL, Santos HRS, Gadig OBF, Signori CN, Vicente MM. Guia para identificação dos tubarões, raias e quimeras do Estado Rio de janeiro (Chondrichthyes: Elasmobrachii e Holocephali). REVNEBIO. 2019; 27(1):171–368. https://doi.org/10.22478/ufpb.2236-1480.2019v27n1.47122
Gomes UL, Signori CN, Gadig OBF, Santos HRS. Guia de Identificação de Tubarões e Raias do Rio de Janeiro. Rio de Janeiro: Technical Books; 2010.
Gomes UL, Winkelstein C, Souza-Lima W. Estudo da cartilagem sinarcual cérvico-torácica em rajídeos (Batomorphii, Rajiformes) da região sudeste do Brasil. An Acad Bras Cienc. 1997; 69(1):95–107.
Günther A. Catalogue of the Fishes in the British Museum. London: British Museum (Natural History); 1870, vol 8. https://doi.org/10.5962/bhl.title.8809
Hubbs CL, Ishiyama R. Methods for the taxonomic study and description of skates (Rajidae). Copeia. 1968; (3):483–91. https://doi.org/10.2307/1442016
Hulley PA. The origin, interrelationships and distribution of southern African Rajidae (Chondrichthyes, Batoidea). Ann S Afr Mus. 1972; 60:1–103. Available from: https://www.biodiversitylibrary.org/part/92166
Last PR, Stehmann MFW, Seret B, Weigmann S. Soft-nose skates. Family Arhynchobatidae. In: Last PR, White WT, Carvalho MR, Seret B, Stehmann MFW, Naylor GJP, editors. Rays of the world. Melbourne: CSIRO Publishing; 2016. p.364–472.
Leible M. Revision de metodos para estudios taxonomicos de rayas (Rajiformes, Rajidae). Gayana. 1988; 52(1–2):15–93.
Lucena CAS, Lucena ZMS. Catálogo dos peixes marinhos do Museu de Ciências da Pontifícia Universidade Católica do Rio Grande do Sul. Elasmobranchiomorphii. Teleostomi (1ª parte). Comun Mus Ci PUCRS. 1981; 21:1–66.
Mabragaña E, Figueroa DE, Scenna LB, Díaz de Astarloa JM, Colonello JH, Massa AM. Clave de identificación de cápsulas de huevos de condrictios del mar argentino. INIDEP Informe Técnico. 2009; 77:1–14. Available from: https://aquadocs.org/handle/1834/3459
Marini TL. Enumeración de los peces coleccionados en las inmediaciones del Laboratorio de Biologia Marina de Puerto Quenquén. Physis. 1929; 9(34):451–54.
Mazzoleni RC, Schwingel PR. Elasmobranch species landed in Itajaí Harbour, Southern Brazil. Notas Técnicas Facimar. 2009; 3:111–18.
McEachran JD. Variation in Raja garmani and status of Raja lentiginosa (Pisces: Rajidae). Bull Mar Sci. 1977; 27(3):423–39. Available from: https://www.ingentaconnect.com/content/umrsmas/bullmar/1977/00000027/00000003/art00004#
McEachran JD, Aschliman N. Phylogeny of Batoidea. In: Carrier JC, Musick JA, Heithaus MR, editors. Biology of sharks and their relatives. Florida: CRC Press; 2004. p.79–113.
McEachran JD, Compagno LJV. A further description of Gurgesiella furvescens with comments on the interrelationships of Gurgesiellidae and Pseudorajidae (Pisces, Rajoidei). Bull Mar Sci. 1979; 29(4):530–53. Available from: https://www.ingentaconnect.com/contentone/umrsmas/bullmar/1979/00000029/00000004/art00009
McEachran JD, Dunn KA. Phylogenetic analysis of skates, a morphologically conservative clade of elasmobranchs (Chondrichthyes, Rajidae). Copeia. 1998; (2):271–90. https://doi.org/10.2307/1447424
McEachran JD, Miyake T. Phylogenetic interrelationships of skates: a working hypothesis (Chondrichthyes, Rajoidei). NOAA Technical Report NMFS. 1990; 90:285–304. Available from: https://www.st.nmfs.noaa.gov/spo/SPO/tr90opt.pdf
Meneses P, Paesch L. Guía de campo para la identificación de peces cartilaginosos en el Rio de la Plata y su frente oceánico. Frente Marítimo. 2003; 19(B):145–93. Available from: http://www.ctmfm.org/upload/archivoSeccion/meneses-y-paesch-142609731427.pdf
Menni RC. Rajidae del litoral bonaerense I. Especies de los generos Raja, Bathyraja y Sympterygia (Chondrichthyes). Physis. 1973; 32(85):413–39. Available from: http://naturalis.fcnym.unlp.edu.ar/repositorio/_documentos/sipcyt/bfa001732.pdf
Menni RC, Ringuelet RA, Aramburu RH. Peces marinos de la Argentina y Uruguay. Buenos Aires: Hemisferio Sur; 1984. Available from: https://core.ac.uk/download/pdf/301080665.pdf
Menni RC, Stehmann MFW. Distribution, environment and biology of batoid fishes off Argentina, Uruguay and Brazil. A review. Rev Mus Argent Cienc Nat. 2000; 2(1):69–109. Available from: http://revista.macn.gob.ar/ojs/index.php/RevMus/article/view/126/118
Miranda Ribeiro A. Pescas do “Annie.” A lavoura: Bol Soc Nac Agricultura. 1903; (4–7):144–96.
Miranda Ribeiro AM. Fauna brasiliense. Peixes. ii (Desmobranchios). Arch Mus Nac Rio de Janeiro. 1907; 14:131–217. Available from: https://www.biodiversitylibrary.org/bibliography/37958
Miranda Ribeiro AM. Fauna brasiliense. Rio de Janeiro: Imprensa Nacional; 1923, vol. 2, Peixes.
Miranda Ribeiro P. Tipos das espécies e subespécies do Prof. Alipio de Miranda Ribeiro depositados no Museu Nacional. Arch Mus Nac Rio de Janeiro. 1953; 42:389–417.
Miranda Ribeiro PM. Pesca do Toku Maru. Bol Mus Nac Zool Rio de Janeiro. 1961; 288:1–18.
Moreira RA, Gomes UL, Carvalho MR. Clasper morphology of skates of the tribe Riorajini (Chondrichthyes: Rajiformes: Arhynchobatidae) and its systematic significance. J Morph. 2017; 278(9):1185–96. https://doi.org/10.1002/jmor.20703
Motta NS, Della-Fina N, Souza CCA, Rodrigues ES, Amorim AF. Analysis of food habits of skate Rioraja agassizii (Elasmobranchii, Rajidae) from southern Brazil. Braz J Biol. 2016; 76(2):469–75. https://doi.org/10.1590/1519-6984.21414
Müller J, Henle FGJ. Systematische Beschreibung der Plagiostomen. Berlin: Veit; 1841. Available from: https://www.biodiversitylibrary.org/item/30065#page/5/mode/1up
Muto EY, Soares LSH, Goiten R. Food resource utilization of the skates Rioraja agassizii (Müller & Henle, 1841) and Psammobatis extenta (Garman, 1913) on the continental shelf of Ubatuba, South-eastern Brazil. Rev Bras Zool. 2001; 61(2):217–38. https://doi.org/10.1590/S0034-71082001000200005
Nishida K. Phylogeny of the suborder Myliobatidoidei. Mem Fac Fish, Hokkaido Univ. 1990; 37(1–2):1–108. Available from: https://eprints.lib.hokudai.ac.jp/dspace/bitstream/2115/21887/1/37(1_2)_P1-108.pdf
Oddone MC, Amorim AF, Mancini PL, Norbis W. Size composition, monthly condition factor and morphometrics for fishery-dependent samples of Rioraja agassizi (Chondrichthyes: Rajidae) off Santos, Southeast Brazil. Neotrop Ichthyol. 2007a; 5(3):415–24. https://doi.org/10.1590/S1679-62252007000300021
Oddone MC, Amorim AF, Mancini PL, Norbis W, Velasco G. The reproductive biology and cycle of Rioraja agassizi (Müller and Henle, 1841) (Chondrichthyes: Rajidae) in southeastern Brazil, SW Atlantic Ocean. Sci Mar. 2007b; 71(3):593–604. https://doi.org/10.3989/scimar.2007.71n3593
Oddone MC, Mesa A, Amorim AF. The egg capsule of Rioraja agassizi (Müller and Henle, 1841) (Chondrichthyes: Rajidae), endemic to the SW Atlantic. Panam J Aquat Sci. 2006; 1(2):43–48. Available from: https://panamjas.org/pdf_artigos/PANAMJAS_1(2)_43-48.pdf
Oddone MC, Vooren CM. Comparative morphology and identification of egg capsules of skate species of the genera Atlantoraja Menni, 1972, Rioraja Whitley, 1939, and Sympterygia Müller & Henle, 1837. Arq Ciênc Mar. 2008; 41(2):5–13. https://doi.org/10.32360/acmar.v41i2.6056
Pasquino AF, Vaske-Júnior T, Gadig OBF, Barreiros JP. Notes on the feeding habits of the skate Rioraja agassizi (Chondrichthyes, Rajidae) off southeastern Brazil. Cybium. 2011; 35(2):105–09. https://doi.org/10.26028/cybium/2011-352-004
Pollom R, Barreto R, Charvet P, Cuevas JM, Faria V, Herman K, Marcante F, Montealegre-Quijano S, Motta F, Paesch L, Rincon G. Rioraja agassizii. The IUCN Red List of Threatened Species. 2020: e.T63109A3118640. https://dx.doi.org/10.2305/IUCN.UK.2020-3.RLTS.T63109A3118640.en
Ringuelet RA, Aramburu RH. Peces Marinos de la Republica Argentina: Clave para el reconocimento de famílias y gêneros. Buenos Aires: Agro Publicacion Tecnica; 1960. Available from: http://www.ecopuerto.com/Bicentenario/informes/PecesMarinosReconocimiento.pdf
Roux C. Résultats scientifiques des campagnes de la “Calypso” au large dês cotes atlantiques de l´Amérique du Sud (1961–1962). Première partie (suite).30. Poisson chondrichthyens du plateau continental brésilien et du Rio de La Plata. Paris: Masson et Cie; 1977.
Ruschi A. Lista dos tubarões, raias e peixes de água doce e salgada do Estado do Espírito Santo e uma observação sobre a introdução do dourado no Rio Doce. Bol Mus Biol Mello Leitão. 1965; 25A:1–24.
Sadowsky V. Relação dos peixes cartilaginosos. In: Relatório sobre a segunda pesquisa oceanográfica e pesqueira do Atlântico sulentre Torres e Maldonado (lat. 29º S 35ºS). Rio Grande do Sul: GEDIP; 1973. p.483–88.
Siqueira AE. Aspectos do desenvolvimento reprodutivo de Rioraja agassizi (Müller & Henle, 1841) (Chondrichthyes, Rajiformes, Rajidae). [Master Dissertation]. Rio de Janeiro: Universidade do Estado do Rio de Janeiro; 1999.
Soares LSH, Vazzoler AEAM, Correia AR. Diel feeding chronology of the skate Raja agassizii (Muller & Henle) (Pisces, Elasmobranchii) on the continental shelf off Ubatuba, Southern Brazil. Rev Bras Zool. 1999; 16(1):201–12. https://doi.org/10.1590/S0101-81751999000100016
Soares KDA, Gomes UL, Santos HRS. Scapulocoracoid morphology in the skates of the tribe Riorajini (Elasmobranchii, Rajiformes, Arhynchobatidae). Zoomorphology. 2020; 139:493–500. https://doi.org/10.1007/s00435-020-00504-2
Vooren CM. Demersal elasmobranchs. In: Seelinger C, Castello JP, editors. Subtropical convergence environments: the coast and sea in the southwestern Atlantic. Berlin: Springer-Verlag; 1997. p.141–46.Whitley GP. Additions to the check-list of the fishes of New South Wales. No. 2. Aust Zool. 1929; 5(4):353–57.
Authors

Karla D. A. Soares1 ![]()
![]() , Renan A. Moreira1
, Renan A. Moreira1 ![]() and Ulisses L. Gomes2
and Ulisses L. Gomes2 ![]()
[1] Laboratório de Ictiologia, Departamento de Zoologia, Instituto de Biociências, Universidade de São Paulo, Rua do Matão, Trav. 14, 101, 05508-900 São Paulo, SP, Brazil. (KDAS) karlad.soares@yahoo.com.br (corresponding author); (RAM) moreirarandrade@gmail.com.
[2] Laboratório de Taxonomia de Elasmobrânquios, Departamento de Zoologia, Instituto de Biologia, Universidade do Estado do Rio de Janeiro, Rua São Francisco Xavier, 524, Maracanã, 20550-900 Rio de Janeiro, RJ, Brazil. (ULG) ulisseslg@netscape.net.
Authors Contribution
Karla D. A. Soares: Data curation, Formal analysis, Investigation, Methodology, Project administration, Visualization, Writing-original draft, Writing-review and editing.
Renan A. Moreira: Data curation, Formal analysis, Writing-original draft, Writing-review and editing.
Ulisses L. Gomes: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Writing-original draft, Writing-review and editing.
Ethical Statement
Not applicable.
Competing Interests
The authors declare no competing interests.
How to cite this article
Soares KDA, Moreira RA, Gomes UL. Redescription of the Rio skate Rioraja agassizii (Rajiformes: Arhynchobatidae) with notes on internal anatomy and intraspecific variation. Neotrop Ichthyol. 2021; 19(2):e210059. https://doi.org/10.1590/1982-0224-2021-0059
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2021 The Authors.
Diversity and Distributions Published by SBI
![]() Submitted February 26, 2021
Submitted February 26, 2021
![]() Accepted May 6, 2021 by Lisa Whitenack
Accepted May 6, 2021 by Lisa Whitenack
![]() Epub Jun 30, 2021
Epub Jun 30, 2021