![]() Leandro Ranucci1
Leandro Ranucci1 ![]() ,
, ![]() Carlos A. Fernandes1,2,
Carlos A. Fernandes1,2, ![]() Luciana A. Borin-Carvalho1,
Luciana A. Borin-Carvalho1, ![]() Isabel C. Martins-Santos1 and
Isabel C. Martins-Santos1 and ![]() Ana L. B. de Portela-Castro1,2
Ana L. B. de Portela-Castro1,2
PDF: EN XML: EN | Cite this article
Abstract
Moenkhausia is a highly specious genus among the Characidae, composed of 96 valid species. Only twelve species have a known karyotype. Thus, here are presented the first cytogenetic data of two allopatric populations of Moenkhausia bonita and one of M. forestii, both belonging to the upper Paraná River basin (PR) with discussion on the evolutionary and cytotaxonomic aspects of the genus. The two species presented 2n = 50 chromosomes but different karyotype formulas and occurrence of 1–2 B chromosomes. These elements are small metacentrics in M. bonita and small acrocentrics in M. forestii. In both species, B chromosomes were euchromatic. Ag-NOR sites were found in pair 3 (metacentric), coinciding with fluorescent in situ hybridization (FISH) by the 18S rDNA probe in both species. However, the species differed in terms of the number and position of 5S rDNA sites. Heterochromatic blocks, mapped in M. bonita showed the least amount of heterochromatin in the terminal and pericentromeric regions, while the M. forestii karyotype revealed a greater amount of interstitial heterochromatic blocks. The karyotype distinctions between the two species, including the morphology of B chromosomes, may contribute as a reference in the taxonomic studies in this group.
Keywords: 5S rDNA, 18S rDNA, C-banding, Fish cytogenetics, Microchromosomes.
Moenkhausia é um gênero altamente especioso dentre os Characidae, composto por 96 espécies válidas, mas apenas doze espécies têm seus cariótipos conhecidos. Portanto, são apresentados aqui os primeiros dados citogenéticos de duas populações alopátricas de Moenkhausia bonita e uma de M. forestii, ambas pertencentes à bacia do alto rio Paraná (PR), com uma ampla discussão sobre os aspectos evolutivos e citotaxonômicos do gênero. As duas espécies apresentaram 2n = 50 cromossomos, mas diferentes fórmulas cariotípicas e ocorrência de 1–2 cromossomos B. Esses elementos são pequenos metacêntricos em M. bonita e acrocêntricos pequenos em M. forestii. Em ambas as espécies, os cromossomos B apresentaram-se eucromáticos. Sítios Ag-NOR foram encontrados no par 3 (metacêntrico), coincidindo com a hibridização fluorescente in situ (FISH) pela sonda 18S rDNA em ambas as espécies. No entanto, as espécies diferiram em termos de número e posição dos sítios de 5S rDNA. Blocos heterocromáticos mapeados em M. bonita revelaram pequena quantidade de heterocromatina nas regiões terminal e pericentromérica, enquanto o cariótipo de M. forestii revelou uma maior quantidade de blocos heterocromáticos intersticiais. As distinções cariotípicas entre as duas espécies, incluindo a morfologia dos cromossomos B, podem contribuir como uma referência em estudos taxonômicos neste grupo.
Palavras-chave: Banda-C, Citogenética de peixes, DNAr 5S, DNAr 18S, Microcromossomos.
Introduction
Moenkhausia Eigenmann, 1903 is one of the most specious genera of the Characidae family and allocated in the Stethaprioninae subfamily (Mirande, 2010), which is widely distributed in several rivers in the Neotropical region. This genus includes small species of variable sizes (approximately 10 cm of total length), commonly known as “lambaris” or “tetras”, and many species are appreciated in aquariums. Moenkhausia currently comprise 96 valid nominal species (Fricke et al., 2021) with a wide variety of forms and pigmentation patterns (Benine et al., 2007).
Phylogenetic relationships are still not well defined in this genus, considering that its taxonomy is based only on a combination of morphological characteristics (number of multicuspid teeth in the internal pre-maxillary series, caudal fin partially covered by scales, and complete lateral line), which, according to Lima et al. (2007), are not unique to the genus and, consequently, result in many doubts about the monophyletic nature of the group. Studies using molecular DNA markers have contributed to the development of hypotheses of phylogenetic relationships in Moenkhausia, suggesting that this is probably a polyphyletic group (Mirande, 2010; Oliveira et al., 2011; Mariguela et al., 2013). In addition, molecular data from COI gene sequences have contributed to the distinction between very similar species morphologically, for example, M. bonita Benine, Castro & Sabino, 2004 and H. marginatus (Benine et al., 2004), which revealed a genetic distance of 7.2% (Mota et al., 2018).
Cytogenetic studies in Moenkhausia are still scarce, presently involving the analysis of twelve species (Tab. 1). Furthermore, the chromosome data registered in species of Moenkhausia have mainly been based on karyotypic characteristics, such as diploid number (2n), total number of arms (NF), karyotype formulas, and distribution of ribosomal genes, evidenced principally by silver nitrate stain (Ag-NOR). Data on the location of ribosomal sites through fluorescent in situ hybridization (FISH) using 18S and/or 5S rDNA probes were recorded only for some species, namely, M. intermedia Eigenmann, 1908, M. sanctaefilomenae (Steindachner, 1907), M. forestii Benine, Mariguela & Oliveira, 2009, M. oligolepis (Günther, 1864), M. cosmops Lima, Britski & Machado, 2007, M. nigromarginata Costa, 1994, Moenkhausia sp. and including the populations of M. forestii and M. bonita analyzed in the present study (Tab. 1).
TABLE 1 | Review of cytogenetic data in Moenkhausia. Abbreviations: m = metacentric; sb = submetacentric; st = subtelocentric; a = acrocentric; FN = fundamental number; cyt = cytotype; B = B chromosome; 5S = 5S rDNA, 18S = 18S rDNA; NOR = AgNOR; Ref = reference; BC = C band; H = heterochromatin; E = euchromatin; P = partially heterochromoatic; * = technique not performed. AM = Amazonas; MG = Minas Gerais; SP = São Paulo; MT = Mato Grosso; MS = Mato Grosso do Sul; PR = Paraná. 1– Portela-Castro et al. (1988); 2– Dantas et al. (2007); 3– Foresti et al. (1989); 4– Alberdi, Fenocchio (1997); 5– Arefjev (1990); 6– Miyazawa (1997); 7– Santos (1999); 8– Hashimoto et al. (2012); 9– Portela-Castro et al. (2001); 10– Scudeler et al. (2015); 11– Utsunomia et al. (2016); 12– Present study; 13– Portela-Castro, Júlio Junior (2002); 14– Fernandes, Alves (2017); 15– Nascimento et al. (2020). Column 18S corresponds to the number of chromosome pairs with 18S rDNA.
Species | Collection site | 2n | Karyotype formula | FN | AgNOR/Par | 18S/Pair | 5S/Pair | B/BC | Ref. |
M.
intermedia | Lake of Mato, Mogi Guaçu River, SP | 50 | 50m/sm | 100 | 6(m/sm) | * | * | 0-1; * | 1 |
M.
intermedia | Paraná River, Paraná River basin, PR | 50 | 16m+34sm | 100 | * | * | * | 0 | 13 |
M.
intermedia | Mogi Guaçu, SP | 50 | 16m+34sm | 100 | 2(m) | 2(m) | * | 0 | 2 |
M.
sanctaefilomenae | Batalha River, Tietê River basin, SP | 50 | 6m+16sm+28st | 100 | 6(m); B | * | * | 2-3; H-E. | 8 |
M.
sanctaefilomenae | Capivara River Tietê River basin, SP | 50 | 48m/sm+2st/a | 98 | 19(sm/m) | * | * | 0-8; H-E. | 3 |
M.
sanctaefilomenae | Aquapey River (Argentina) | 50 | 48m/sm+2st | 100 | * | * | * | 0-3; * | 4 |
M.
sanctaefilomenae | Paraná River, Paraná River basin, PR | 50 | 12m+36sm+2st | 100 | * | * | * | 0-2; P. | 9 |
M.
sanctaefilomenae | Capivara stream tributary, Tietê River basin, SP | 50 | 12m+36sm+2st | 100 | sm | 3 pairs (m), 2 pairs (sm), 2 -B | * | 0-6; H-E. | 2 |
M.
sanctaefilomenae | Araquá stream, tributaries of Tietê River, SP | 50 | 6m+44sm | 100 | * | 7,12,15,17(sm);
1-B | * | 0-4; * | 10 |
M.
sanctaefilomenae | Olaria stream, tributaries of Tietê River, SP | 50 | 6m+44sm | 100 | * | 4,7,10,15(sm);
1-B | * | 0-4; * | 10 |
M.
sanctaefilomenae | Mané Teixeira stream, tributaries of Tietê
River, SP | 50 | 6m+44sm | 100 | * | 7,10,14,15,17(sm) | * | 0-2* | 10 |
M.
sanctaefilomenae | Guaçu stream, Paraná River basin, MS | 50 | 8m+36sm+6st | 100 | 24(st) | * | * | 0-8; P. | 14 |
M.
sanctaefilomenae | Batalha River, Tietê River basin, SP | 50 | 6m+16sm+28st | 100 | 6(m); B. | 6(m); 1-B | 1,2(m); 5,7(sm) | 0-6; H-E. | 11 |
M.
sanctaefilomenae | Novo River, Paranapanema River basin, SP | 50 | 6m+16sm+28st | 100 | 6(m). | 6(m) | 1,2(m); 5,6,7 (sm);
17,18(st) | 0 | 11 |
M.
costae | São Francisco River, MG | 50 | 50m/sm | 100 | 6(m/sm) | * | * | 0 | 1 |
M.
pittieri | not registered | 49 | 4m+39sm/st+6a | 92 | * | * | * | 0 | 5 |
M.
pittieri | not registered | 50 | 4m+40sm/st+6a | 94 | * | * | * | 0 | 5 |
M.
dichroura | Cuiabá River, MT | 50 | 32m+14sm+4st | 100 | * | * | * | 0 | 6 |
M.
gracilima (cyt a) | Amazonas River, AM | 50 | 14m+26sm+6st+4a | 96 | * | * | * | 0 | 7 |
M.
gracilima (cyt b) | Amazonas River, AM | 48 | 4m+24sm+12st+8a | 88 | * | * | * | 0 | 7 |
Moenkhausia
sp. | Taquaral stream, Barra do Garças region, MT | 50 | 16m+34sm | 100 | sm | sm | * | 0 | 2 |
M.
bonita | Índios River, Paraná River basin, PR | 50 | 14m+24sm+8st+4a | 96 | 3(m) | 3(m). | 8,14(sm);
20,22(st) | 0-1; E. | 12 |
M.
bonita | Paraná River, Paraná River basin, PR | 50 | 14m+24sm+8st+4a | 96 | 3(m) | 3(m) | 9(sm); 23 (st) | 0-2; E. | 12 |
M.
forestii | Paraná River, Paraná River basin, PR | 50 | 10m+28sm+10st+2a | 98 | 3(m) | 3(m) | 1(m) | 0-2; E. | 12 |
M.
forestii | Sapo stream, Tangará da Serra, MT | 50 | 10m+32sm+8st | 100 | 22(st) | 22(st) | 1,2(m);6,8,10 (sm) | 0-3; H-E-P | 15 |
M.
oligolepis | Corredeira stream, Denise, MT | 50 | 12m+32sm+6st | 100 | 23(st) | 23(st) | 17 chromosomes | 0-4; H-E-P | 15 |
M.
oligolepis | Sangue River, Campo Novo do Parecis, MT | 50 | 12m+32sm+6st | 100 | 22(sm) | 22(sm) | 4 chromosomes | 0-3, H-E-P | 15 |
M.
oligolepis | Nameless stream, Xapuri, AC | 50 | 10m+26sm+14st | 100 | 17(sm) | 7 chromosomes | 21 chromosomes | 0-2; P | 15 |
M.
cosmops | Verde River, Campo Novo do Parecis, MT | 50 | 14m+30sm+6st | 100 | 19(sm) | 19(sm) | 1,2(m) | 0 | 15 |
M.
nigromaginata | Verde River, Campo Novo do Parecis, MT | 50 | 14m+32sm+4a | 96 | 16(sm) | 16(sm) | 19(sm);24,25(a) | 0 | 15 |
M.
nigromarginata | Membeca River, Campo Novo do Parecis, MT | 50 | 14m+32sm+4a | 96 | 16(sm) | 16(sm) | 24,25(a) | 0 | 15 |
Moenkhausia
sp. | Membeca River, Campo Novo do Parecis, MT | 50 | 10m+32sm+8st | 100 | 22(sm) | 22sm | 1(m);6(sm) | 0 | 15 |
The occurrence of B chromosomes in the species of Moenkhausia was initially documented in specimens of M. intermedia (Portela et al., 1988) and M. sanctaefilomenae (Foresti et al., 1989). B microchromosomes were described in M. forestii while individuals of three populations of M. oligolepis presented three morphologically distinct types of B chromosomes (Nascimento et al., 2020). The B chromosomes of M. sanctaefilomenae, characterized as microchromosomes due to their small size, are highly variable in number. They occur in cells with 1 to 8 of these elements, whose differences are found intra and inter-individually. These elements also vary in the amount of heterochromatin and can be euchromatic, partially euchromatic, or completely heterochromatic (Foresti et al., 1989). For some populations of M. sanctaefilomenae, these extra chromosomes have some specific characteristics, such as restricted to males (Portela-Castro et al., 2000) or presence of some B chromosome carriers of ribosomal genes (Hashimoto et al., 2012, Scudeler et al., 2015), arousing the interest of cytogenetic studies in different populations of this species with a focus on its origin and their evolution, as well as using more resolutive methodologies, such as microdissection of these elements and other chromosomal markers (Hashimoto et al., 2012; Scudeler et al., 2015; Utsunomia et al., 2016).
Although B chromosomes, also referred to as supernumerary or accessory chromosomes, occur in most eukaryotic genomes, the understanding of their origin, function, maintenance, and evolution in carrier species is still a great challenge, and the molecular mechanisms involved in their transmission and maintenance in populations are still unclear (Camacho et al., 2000; Houben et al., 2014; 2019). However, some modern technologies, such as data sequencing of DNA, have demonstrate that they are rich in repetitive elements and originated from A chromosomes (Utsunomia et al., 2016; Clark et al., 2017; Coan, Martins, 2018; Dahr et al., 2019). According to Ahmad, Martins (2019), B chromosomes are enriched with genes for many significant biological functions, such as those related to the cell cycle and chromosome structure, being able to favor genomic rearrangements and influence the nuclear environment, affecting the function of other chromatin regions. These findings provide new insight into studies on chromosome B and evolutionary implications bearing species.
Moenkhausia bonita and M. forestii are species described for the Paraguay River basin, in the state of Mato Grosso do Sul, whose typical locations are the Baía Bonita River (a tributary of the Miranda River, Bonito city) and Sepotuba River (Cáceres city), respectively. However, specimens corresponding to these species have been found in the upper Paraná River floodplain, in the Porto Rico region, or small tributaries of this basin (Benine et al., 2009; Mota et al., 2018).
To obtain more information about the chromosomal diversity in Moenkhausia, this study established the first cytogenetic description of M. bonita and M. forestii, as well as to contribute to records of B chromosomes present in these populations collected in the upper Paraná River basin, Brazil. We also compiled the cytogenetic data available for the genus to provide a more systematic overview of the karyotypic structure and its evolutionary aspects.
Material and methods
Thirty-four individuals of Moenkhausia were collected from populations in the upper Paraná River basin: 12 (three males, one female and eight sex indeterminate) individuals of M. bonita from Índios River, Cianorte, Brazil (23º43’50.2”S 52º44’12.2”W); 11 (four males and seven females) individuals of M. bonita and 11 (three females and eight sex indeterminate) individuals of M. forestii from the Paraná River, Porto Rico, Brazil (22º45’55.6”S 53º15’25.9”W).
Voucher specimens were deposited in the fish collection of the Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (NUPELIA), Universidade Estadual de Maringá, Maringá, Paraná, Brazil, as Moenkhausia bonita from Índios River (NUP 22691), Moenkhausia bonita from Paraná River (NUP 22692), and Moenkhausia forestii (NUP 22693).
The experiments followed the ethical conduct, and before euthanasia, the fish were anesthetized by an overdose of clove oil (Griffiths, 2000). Metaphase chromosomes were obtained from anterior kidney cells using the air-drying technique (Bertollo et al., 1978). C-positive heterochromatin (C-bands) was visualized by the procedure of Sumner (1972) with modifications suggested by Lui et al. (2012). NORs were detected employing silver nitrate staining (Ag-NORs), according to Howell, Black (1980).
The chromosomes were classified as metacentric (m), submetacentric (sm), subtelocentric (st), and acrocentric (a) according to Levan et al. (1964). The fundamental number (FN) was calculated according to the chromosomal arm number (chromosomes m, sm, and st are considered to contain two arms – p (smaller arm)and q (long arm)arms – and those with one arm – only a q arm.
Physical mapping of the 5S and 18S rDNA was carried out by FISH according to Pinkel et al. (1986) and modifications suggested by Margarido, Moreira-Filho (2008), using DNA probes obtained from the genomes of Megaleporinus elongatus (Valenciennes, 1850) (Martins, Galetti, 1999) and Prochilodus argenteus Spix & Agassiz, 1829 (Hatanaka, Galetti, 2004), respectively. The probes were labelled through nick translation with digoxigenin-11-dUTP (5S rDNA) and biotin-16-dUTP (18S rDNA) (Roche®). Detection and amplification of the hybridization signal was carried out using avidin-FITC and anti-avidin biotin (Sigma-Aldrich) for the probe of 18S rDNA, and anti-digoxigenin rhodamine (Roche®) for the probe of 5S rDNA. Slides were counterstained with DAPI (4’6-Diamine-2’-phenylindole dihydrochloride; 50 μg ml−1) and analyzed in an epifluorescence microscope (Olympus BX51). The images were captured using the software DP controller (Media Cybernetics), and image composition was carried out with Adobe Photoshop CS6.
Results
Moenkhausia bonita – Índios and Paraná Rivers (upper Paraná River basin). Both populations had the same diploid number of 2n = 50, karyotype formula consisting of 14m + 24sm + 8st + 4a, and the fundamental number, NF = 96. In addition, male and female specimens from both populations showed cells with variations of 0–2 B chromosomes (Figs. 1A,B). B chromosomes are small metacentrics and in M. bonita from the Índios River have size equivalent to pair 5 (Fig. 1A), while in M. bonita from the Paraná River are the smallest of the standard complement (Fig. 1B). The frequencies of the B chromosomes for each population of M. bonita revealed an intra- and inter-individual numerical variability (Tab. 2). The B chromosomes were detected in 65.87% and 43.29% of metaphases in specimens from Índios and Paraná rivers, respectively. Note that cells with 1 B chromosome were more frequent in individuals of both populations in relation to the number of cells bearing B chromosomes (Tab. 2).

FIGURE 1 | Karyotypes A and D Moenkhausia bonita from the Índios River, B and E M. bonita from the Paraná River, C and F M. forestii from the Paraná River. Karyotypes A, B, and C stained with Giemsa, D, E, and F C-banded. In evidence the AgNOR-bearing chromosomes (box) and B chromosomes. Scales bar = 10 µm.
Ag-NOR sites were located on a single chromosome pair in karyotypes of all individuals in both populations. These sites were located at the terminal position on the p arm of pair 3 in population of Índios River (Fig. 1A, in box) and for individuals of the population from the Paraná River (Fig. 1B). In this last population, only one Ag-NOR chromosome was active. C-banding showed few marked chromosomes; only a large heterochromatic block in the p arm of pair 3 (Ag-NOR) occurred in the specimens from the Índios River (Fig. 1C). In the specimens of the population of M. bonita from the Paraná River, heterochromatin was detected on the pericentromeric region of the pairs 2, 9, 13, and 16 and a large heterochromatic block in the p arm of pair 3 (Fig. 1E). Additionally, similar to euchromatin in the A chromosomes, B chromosomes were faintly stained with C-banding, suggesting that they were euchromatic in both populations (Figs. 1D,E). Double FISH with 18S and 5S rDNA probes are shown in Fig. 2. Moreover, 18S rDNA-FISH (green signs) confirmed the Ag-NOR pair (No 3) in both populations and 5S rDNA-FISH (red signs) showed that minor rDNA clusters occur in pericentromeric regions on pairs 8, 14, 20 and 22 in the specimens from the Índios River (Fig. 2A). In the population from the Paraná River, these sites occurred in a terminal position on the p arm of pairs 9 and 23 (Fig. 2B).
Moenkhausia forestii – Paraná River (upper Paraná River basin). The diploid number was 50 chromosomes (10m + 28sm + 10st + 2a, FN = 98). In addition to the basic karyotype, the analyzed specimens presented a variation from zero to two B chromosomes in the somatic cells (Fig. 1C). These elements are smaller than any chromosome of the normal A complement. B chromosomes were detected in 36.53% of metaphases, but cells with 1 B chromosome were more frequent (34.70%, Tab. 2).
TABLE 2 | Frequency of B chromosomes in metaphases of Moenkhausia bonita 1 from Índios River (Paraná, Brazil), M. bonita 2 and M. forestii from Paraná River (Paraná, Brazil). Simbols: ♀ female; ♂ male; V unidentified sex.
Individuals M. bonita 1 Sex | Number of | Total metaphases | Individuals M. bonita 2 Sex | Number of | Total metaphases | Individuals M. forestii Sex | Number of | Total metaphases | ||||||
0 | 1 | 2 | 0 | 1 | 2 | 0 | 1 | 2 | ||||||
1 ♂ | 1 | 1 | 0 | 2 | 1 ♂ | 6 | 0 | 0 | 6 | 1 V | 1 | 3 | 0 | 4 |
2 V | 5 | 2 | 0 | 7 | 2 ♀ | 3 | 2 | 0 | 5 | 2 ♀ | 4 | 6 | 0 | 10 |
3 ♂ | 0 | 1 | 0 | 1 | 3 ♀ | 174 | 4 | 0 | 178 | 3 V | 2 | 4 | 0 | 6 |
4 V | 8 | 4 | 0 | 12 | 4 ♂ | 42 | 81 | 7 | 130 | 4 ♀ | 11 | 3 | 0 | 14 |
5 V | 5 | 14 | 7 | 26 | 5 ♀ | 7 | 31 | 5 | 43 | 5 V | 5 | 5 | 0 | 10 |
6 ♂ | 6 | 13 | 0 | 19 | 6 ♀ | 2 | 20 | 2 | 24 | 6 ♀ | 46 | 4 | 0 | 50 |
7 V | 4 | 11 | 1 | 16 | 7 ♀ | 0 | 5 | 9 | 14 | 7 V | 8 | 21 | 2 | 31 |
8 ♀ | 10 | 26 | 0 | 36 | 8 ♂ | 2 | 5 | 0 | 7 | 8 V | 46 | 10 | 1 | 57 |
9 V | 1 | 1 | 0 | 2 | 9 ♂ | 1 | 3 | 6 | 10 | 9 V | 3 | 0 | 0 | 3 |
10 V | 0 | 1 | 0 | 1 | 10 ♀ | 0 | 2 | 0 | 2 | 10 V | 1 | 2 | 0 | 3 |
11 V | 3 | 0 | 0 | 3 | 11 ♀ | 8 | 5 | 0 | 13 | 11 V | 12 | 18 | 1 | 31 |
12 V | 0 | 1 | 0 | 1 | ||||||||||
Total % | 43 34.13 | 75 59.52 | 8 6.35 | 126 | Total % | 245 56.71 | 158 36.58 | 29 6.71 | 432 | Total % | 139 63.47 | 76 34.70 | 4 1.83 | 219 |
Ag-NOR sites were located on a single chromosome pair in karyotypes of all individuals. These sites were located at the terminal position on the p arm of pair 3 (Fig. 1C, in box). Heterochromatin was detected in the centromeric/pericentromeric region, highlighting interstitial blocks in the long arms of most chromosomes, including Ag-NOR pair (No 3, Fig. 1F). Additionally, B chromosomes were faintly stained with C-banding, suggesting that they were euchromatic (Fig. 1F).
Double FISH with 18S and 5S rDNA are shown in Fig. 2C. Ag-NORs sites were confirmed by 18S rDNA-FISH (Fig. 2C). Additionally, 5S rDNA-FISH showed that minor rDNA clusters occurred in the pericentromeric region of pair 1 (Fig. 2C).
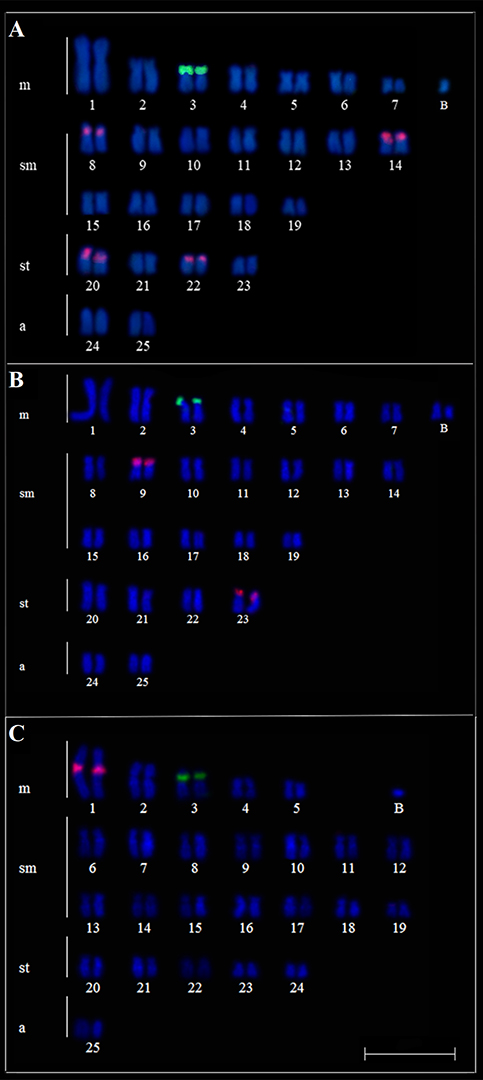
FIGURE 2 | Karyotypes A.Moenkhausia bonita from the Índios River, B.M. bonita from the Paraná River, and C.M. forestii from the Paraná River after double-FISH with 18S rDNA (green) and 5S rDNA (red) probes. In evidence B chromosomes (B, C). Scales bar = 10 µm.
Discussion
Our results showed that individuals from both populations of M. bonita had the same 2n = 50, FN = 96, and identical karyotypes composed of 14m + 24sm + 8st + 4a chromosomes, as well as other chromosomal characteristics revealed by Ag-NOR and 18S-FISH, differing in the C-banding pattern and number of 5S rDNA sites. Moreover, the individuals of M. forestii differed from that of M. bonita in their karyotypes (10m + 28sm + 10st + 2a). FN values (98), C-banding pattern, and number of 5S rDNA sites. Additionally, the karyotype structure of M. forestii proved to be distinct, not only from M. bonita, but from all other species of Moenkhausia, as can be seen in Tab. 1.
Cytogenetic studies in Moenkhausia revealed few variations concerning the diploid number, with 2n = 50 the predominant value (Tab. 1). In contrast, 2n = 48 was registered in M. gracilima Eigenmann, 1908, designed as a specimen of cytotype b (Arai, 2011) and a divergent value for M. pittieri Eigenmann, 1920 with 2n = 49 according to Arefjev (1990). These values, in principle, suggest the occurrence of chromosomal rearrangements of type fusions, reducing the diploid number from 50 to 48, for example. However, more karyotypes and resolute studies in Moenkhausia are needed that could prove this hypothesis.
Although species of Moenkhausia have a relatively conservative karyotype macrostructure, the occurrence of B chromosomes constitutes a differential macrostructure for this genus, which has been observed in other small characids. Portela et al. (1988) recorded cells with 1 microchromosome B in M. intermedia collected from the Lake of Mato (located near the Mogi-Guaçu River, SP), and later, Foresti et al. (1989) showed a variation of 1 to 8 B chromosomes in a M. sanctaefilomenae population of the Tietê River (upper Paraná River basin). B chromosomes have also been recorded in other populations of M. sanctaefilomenae with similar numerical variation (Dantas et al., 2007; Hashimoto et al., 2012; Utsunomia et al., 2016). However, for the population of M. sanctaefilomenae collected in the Aquapey River (Corrientes, Argentina) Alberdi, Fenocchio (1997) observed 1 to 3 extra chromosomes, while Portela-Castro et al. (2000) registered 1 to 2 B chromosomes that were restricted to male individuals in the population of the Paraná River. B chromosomes have also been recorded in M. forestii and three populations of M. oligolepis (Nascimento et al., 2020).The evidence of small B chromosomes in M. bonita contributes as a new reference in this group, totalizing five species (M. intermedia, M. sanctaefilomenae, M. oligolepis, M. forestii, and M. bonita) with B chromosomes in Moenkhausia.
The frequencies of cells with and without carriers of 1–2 B chromosomes (Tab. 2), in both species, demonstrate an inter and intra-individual variability of these elements, suggesting mitotic instability, probably due to their non-Mendelian behavior that may be related to chromosomal non-disjunction during meiosis, leading to uneven segregation of genetic material between germ cells (Da Rosa et al., 2014).
Utsunomia et al. (2016) reported that M. sanctaefilomenae harbours highly invasive B chromosomes, which are present in all populations analyzed to date in the Parana and Tietê Rivers. Therefore, the description of new occurrences of B chromosomes, as presented here demonstrate that these elements probably have an important evolutionary role in the genus.
B chromosomes of M. bonita and M. forestii after C-banding presented themselves as euchromatic, a pattern similar to that observed in some B chromosomes of M. sanctaefilomenae (Foresti et al., 1989; Hashimoto et al., 2012; Fernandes, Alves, 2017), indicating a difference in the molecular composition of these elements, at the interspecific and even intraspecific level (M. sanctaefilomenae, for example). Analysis of the B chromosomes contents of different populations of M. sanctaefilomenae,employing FISH mapping, chromosome painting and DNA sequencing demonstrated several types of repetitive DNA sequences (18S rDNA, H3 histone genes, and two satellite DNAs) present in heterochromatic and euchromatic B chromosomes (two B variants), suggesting their origin, which is presumably independent from the same A chromosome (autosome no. 6), but suggesting that the heterochromatic B chromosome shows signs of being more recent than euchromatic B chromosomes (Utsunomia et al., 2016). The B chromosomes contents in M. forestii and of different populations of M. oligolepis, also demonstrated several types of repetitive DNA sequences (Nascimento et al., 2020). The chromosomal mapping of the 18S rDNA sites using the FISH technique revealed signals in the B chromosomes of M. forestii, while clusters of the H1 histone and U2 snDNA genes were found in the B chromosomes of M. forestii and M. oligolepis (Nascimento et al., 2020). The fact that the B chromosomes of M. bonita and M. forestii are euchromatic does not imply that they do not have repetitive sequences in their composition, since no 18S rDNA and 5S rDNA signals were detected on these chromosomes. Thus, this heterogeneity in the molecular composition of B chromosomes in Moenkhausia species suggests an independent evolution for each species.
The amount and distribution of heterochromatin are variable among Moenkhausia species. The population of M. bonita from the Índios River showed little heterochromatin in its karyotype structure, highlighting only the nucleolar pair. A similar situation was observed in specimens of M. intermedia from the Paraná River (Portela-Castro, Júlio Júnior, 2002). In addition, the population of M. bonita from the Paraná River presented few banded chromosomes (4 pairs), indicating that the small amount of heterochromatin may be a characteristic of this species. Conversely, M. forestii presented a distribution of pericentromeric and interstitial heterochromatic blocks in the long arms of several chromosomes, a pattern similar to that reported for M. sanctaefilomenae, M. intermedia, Moenkhausia sp. (Dantas et al., 2007), and for populations of M. sanctaefilomenae analyzed by Portela-Castro, Júlio-Junior (2002) and Hashimoto et al. (2012). Thus, the heterochromatin distribution pattern proved to be an efficient cytotaxonomic marker in the distinction of M. bonita and M. forestii.
Simple NOR, observed in M. bonita and M. forestii, is the most frequent pattern evidenced for the species of Moenkhausia, except for M. sanctaefilomenae, in which populations with simple and multiple NOR were observed (Tab. 1). In addition, the M. bonita and M. forestii populations shared the same pair of nucleolar, reinforcing the conservative character for these genes. In contrast, few Moenkhausia species have sites of 5S rDNA, covering only populations of M. sanctaefilomenae, M. oligolepis, M. forestii, M. cosmops, M. nigromarginata, and Moenkhausia sp. (Tab. 1) and the species of the present study, M. bonita and M. forestii. In all species analyzed, 5S rDNA sites were found on chromosomes distinct from the nucleolar pair; however, these sites were distributed on more than one chromosome pair, in interstitial and subterminal positions (Tab. 1). The populations of M. bonita from Índios and Paraná rivers differed in terms of the number of 5S rDNA sites, as well as the position, revealing polymorphism in these ribosomal sites. A similar situation was observed regarding the distribution of 5S rDNA sites between two populations of M. sanctaefilomenae from different locations, among other repetitive sequences (Utsunomia et al., 2016). Studies with other small characids on the distribution of the 5S rDNA genes, such as in Astyanax Baird & Girard, 1854, also revealed numerical variability, being detected from 2 to 6 5S rDNA sites in several species of this genus, which are still variable in position (Kavalco et al., 2016; Piscor et al., 2017; Gavazzoni et al., 2018; Teixeira et al., 2018). The presence of multiple 5S sites associated with heterochromatic regions and transposable elements have been identified in some fish species, such as in Ancistrus, suggesting that their dispersion is mediated by these elements (Prizon et al., 2017, 2018). This could occur in the dispersion of these sites in M. bonita, providing the difference in the number of sites between the two populations analyzed here.
In conclusion, karyotype differences between the species of M. bonita (two populations) and M. forestii, including the morphology of the B chromosomes, demonstrate an inter and intraspecific karyotype variability in the chromosomal microstructure of the species only evidenced by repetitive sequence markers (such as mapping of ribosomal genes of 18S and 5S rDNA). Additionally, the quantity and distribution of constitutive heterochromatin between M. bonita and M. forestii constitutes another distinguishing feature between them. The data presented reinforce the taxonomy, corroborating the existence of two species, M. bonita and M. forestii. These results reflect effective mechanisms in karyotype evolution for this group. In addition, new occurrences of small B chromosomes in Moenkhausia, as shown in this study, totals five species carrying these extra elements so far. It has been suggested that these elements were not recently occurring in this genus and evolved independently for each species.
Acknowledgments
This study was financed by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico). The authors thank Dr. Weferson J. da Graça (NUPELIA/UEM) for the taxonomic identification of specimens. We are grateful to the Ministério do Meio Ambiente/Instituto Chico Mendes de Conservação da Biodiversidade (MMA/ICMBio; license number 58187–2) for authorizing the collection of the biological material.
References
Ahmad SF, Martins C. The moderns view of B chromosomes under the impact of high scale omics analyses. Cells. 2019; 8(2):156. https://doi.org/10.3390/cells8020156
Alberdi AJ, Fenocchio A. Karyotypes of five Tetragonopterinae species (Pisces, Characidae) from Argentina. Cytologia. 1997; 62(2):171–76. http://doi.org/10.1508/cytologia.62.171
Arai R. Fish karyotypes: a check list. Japan: Springer; 2011.
Arefjev VA. Problems of karyotypic in the family Characidae (Pisces, Characiformes) with the description of somatic karyotypes for six species of tetras. Caryologia. 1990; 43(3–4):305–19. https://doi.org/10.1080/00087114.1990.10797009
Benine RC, Castro RMC, Sabino J. Moenkhausia bonita: a new small characin fish from the rio Paraguay basin, southwestern Brazil (Characiformes: Characidae). Copeia. 2004; (1):68–73. https://doi.org/10.1643/CI-03-008R1
Benine RC, Castro RMC, Santos ACA. A new Moenkhausia Eigenmann, 1903 (Ostariophysi: Characiformes) from Chapada Diamantina, rio Paraguaçu basin, Bahia, Northeastern Brazil. Neotrop Ichthyol. 2007; 5(3):259–62. http://dx.doi.org/10.1590/S1679-62252007000300004
Benine RC, Mariguela TC, Oliveira C. New species of Moenkhausia Eigenmann, 1903 (Characiformes: Characidae) with comments on the Moenkhausia oligolepis species complex. Neotrop Ichthyol. 2009; 7(2):161–68. https://doi.org/10.1590/S1679-62252009000200005
Bertollo LAC, Takahashi CS, Moreira-Filho O. Cytotaxonomic considerations on Hoplias lacerdae (Pisces, Erythrinidae). Rev Bras Genet. 1978; 1:103–20.
Camacho JPM, Sharbel TF, Beukeboom LW. B-chromosome evolution. Philos Trans R Soc London. 2000; 355(1394):163–78. http://doi.org/10.1098/rstb.2000.0556
Clark FE, Conte MA, Ferreira-Bravo IA, Poletto AB, Martins C, Kocher TD. Dynamic sequence evolution of a sex-associated B chromosome in Lake Malawi Cichlid Fish. J Hered. 2017; 108(1):53–62. https://doi.org/10.1093/jhered/esw059
Coan RLB, Martins C. Landscape of transposable elements focusing on the B chromosome of the ciclid fish Astatotilapia latifasciata. Genes. 2018; 9(6):269. https://doi.org/10.3390/genes9060269
Dahr MK, Kour J, Kaul S. Origin, behaviour, and transmission of B chromosome with special reference to Plantago lagopus. Genes. 2019; 10(2):152. https://doi.org/10.3390/genes10020152
Dantas ESO, Vicari MR, Souza IL, Moreira-Filho O, Bertollo LAC, Artoni RF. Cytotaxonomy and karyotype evolution in Moenkhausia Eigenmann, 1903 (Teleostei, Characidae). The Nucleus. 2007; 50(3):505–38.
Fernandes CA, Alves DS. Occurrence of multiple euchromatic B Microchromosomes in Moenkhausia sanctaefilomenae (Pisces, Characidae) from the upper Paraná River basin, Brazil. Cytologia. 2017; 82(5):545–50. https://doi.org/10.1508/cytologia.82.547
Foresti F, Almeida-Toledo LF, Toledo-Filho AS. Supernumerary chromosome, C-banding pattern characterization and multiple nucleolus organizer regions in Moenkhausia sanctafilomenae (Pisces, Characidae). Genetica. 1989; 79:107–14. https://doi.org/10.1007/BF00057927
Fricke R, Eschmeyer WN, Van der Laan R, editors. Eschmeyer’s catalog of fishes: genera, species, references [Internet]. São Francisco: California Academy of Sciences; 2021. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Gavazzoni M, Paiz LM, Oliveira CAM, Pavanelli CS, Graça WJ, Margarido VP. Morphologically cryptic species of the Astyanax bimaculatus ‘‘caudal peduncle spot’’ subgroup diagnosed through cytogenetic characters. Zebrafish. 2018; 15(4):382–88. https://doi.org/10.1089/zeb.2018.1574
Griffiths S. The use of clove oil as an anaesthetic and method for sampling intertidal rockpool fishes. J Fish Biol. 2000; 57(6):1453–64. https://doi.org/10.1111/j.1095-8649.2000.tb02224.x
Hashimoto DT, Voltolin TA, Paes ADNVA, Foresti F, Bortolozzi J, Porto-Foresti F. Cytogenetic analysis of B chromosomes in one population of the fish Moenkhausia sanctaefilomenae (Steindachner, 1907) (Teleostei, Characiformes). Comp Cytogenet. 2012; 6(2):141–51. https://doi.org/10.3897/CompCytogen.v6i2.1769
Hatanaka T, Galetti Jr., PM. Mapping of the 18S and 5S ribosomal RNA genes in the fish Prochilodus argenteus Agassiz, 1829 (Characiformes, Prochilodontidae). Genetica. 2004; 122(3):239–44. https://doi.org/10.1007/s10709-004-2039-y
Houben A, Banei-Moghaddam AM, Klemme S, Timmis JN. Evolution and biology of supernumerary B chromosomes. Cel Mol Life Sci. 2014; 71:467–78. https://doi.org/10.1007/s00018-013-1437-7
Houben A, Jones N, Martins C, Trifonov V. Evolution, composition and regulation of supernumerary B Chromosomes. Genes. 2019; 10(2):161. http://doi.org/10.3390/genes10020161
Howell WM, Black DA. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia. 1980; 36:1014–15. https://doi.org/10.1007/BF01953855
Kavalco KF, Pazza R, Brandão KO, Garcia C, Bertollo LAC, Almeida-Toledo LF. Chromosomal diversification higher than molecular variation in Astyanax aff. fasciatus (Teleostei, Characidae). Zebrafish. 2016; 13(4):345–43. https://doi.org/10.1089/zeb.2016.1272
Levan A, Fredga K, Sandberg AA. Nomenclature for centromeric position on chromosomes. Hereditas. 1964; 52(2):201–20. https://doi.org/10.1111/j.1601-5223.1964.tb01953.x
Lima FCT, Britski HA, Machado FA. A new Moenkhausia (Characiformes: Characidae) from central Brazil, with comments on the area relationship between the upper rio Tapajós and upper rio Paraguai systems. Aqua. 2007; 13(2):45–54.
Lui RL, Blanco DR, Moreira Filho O, Margarido VP. Propidium iodide for making heterochromatin more evident in the C-banding technique. Biotech Histochem. 2012; 87(7):433–38. http://doi.org/10.3109/10520295.2012.696700
Margarido VP, Moreira-Filho OM. Karyotypic differentiation through chromosome fusion and number reduction in Imparfinis hollandi (Ostariophysi, Heptapteridae). Gen Mol Biol. 2008; 31(1):235–38. https://doi.org/10.1590/S1415-47572008000200012
Mariguela TC, Benine RC, Abe KT, Avelino GS, Oliveira C. Molecular phylogeny of Moenkhausia (Characidae) inferred from mitochondrial and nuclear DNA evidence. J Zool Syst Evol Res. 2013; 51(4):327–32. https://doi.org/10.1111/jzs.12025
Martins C, Galetti Júnior PM. Chromosomal localization of 5S rDNA genes in Leporinus fish (Anastomidae, Characiformes). Chromosome Res. 1999; 7:363–67. https://doi.org/10.1023/A:1009216030316
Mirande JM. Phylogeny of the family Characidae (Teleostei: Characiformes): from characters to taxonomy. Neotrop Ichthyol. 2010; 8(3):385–568. http://dx.doi.org/10.1590/S1679-62252010000300001
Miyazawa CS. Citogenética de caracídeos da bacia do rio Paraguai. Análises citotaxonômicas – evolutivas e considerações biogeográficas. [PhD Thesis]. São Carlos: Universidade Federal de São Carlos; 1997.
Mota TFM, Fabrin TMC, Deprás GC, Gasques LS, Oliveira AV, Pavanelli CS, Prioli SMAP, Prioli AJ. Molecular characterization of Moenkhausia (Pisces: Characiformes) populations with different lateral line developmental levels. An Acad Bras Cien. 2018; 90(3):2815–25. http://dx.doi.org/10.1590/0001-3765201820170493
Nascimento CN, Troy WP, Pansonato-Alves JC, Carvalho ML, Oliveira C, Foresti, F. Molecular cytogenetic analyses reveal extensive chromosomal rearrangements and novel B chromosomes in Moenkhausia (Teleostei, Characidae). Genet Mol Biol. 2020; 43(4):1–09. http://dx.doi.org/10.1590/1678-4685-GMB-2020-0027
Oliveira C, Avelino GS, Abe KT, Mariguela TC, Benine RC, Ortí G, Vari RP, Castro RMC. Phylogenetic relationships within the speciose family Characidae (Teleostei: Ostariophysi: Characiformes) based on multilocus analysis and extensive ingroup sampling. BMC Ecol Evol. 2011; 11:275. https://doi.org/10.1186/1471-2148-11-275
Pinkel D, Straume T, Gray JW. Cytogenetic analysis using quantitative, high-sensitivity, fluorescence in situ hybridization. Proc Natl Acad Sci USA. 1986; 83(9):2934–38. http://doi.org/10.1073/pnas.83.9.2934
Piscor D, Centofante L, Parise-Maltempi PP. Distinct classical and molecular cytogenetics of Astyanax marionae and A. fasciatus (Characiformes: Characidae): a comparative study of the organization of heterochromatin and repetitive genes. J Genet. 2017; 96:665–71. https://doi.org/10.1007/s12041-017-0813-8
Portela ALB, Galetti Júnior PM, Bertollo LAC. Considerations on the chromosome evolution of Tetragonopterinae (Pisces, Characidae). Rev Bras Gen. 1988; 11:307–16.
Portela-Castro ALB, Júlio Junior HF. Karyotype relationships among species of subfamily Tetragonopterinae (Pisces, Characidae): citotaxonomy and evolution aspects. Cytologia. 2002; 67(3):329–36. https://doi.org/10.1508/cytologia.67.329
Portela-Castro ALB, Júlio Junior HF, Nishiyama PB. New occurrence of microchromosomes B in Moenkhausia sanctaefilomenae (Pisces, Characidae) from the Paraná River of Brazil: analysis of the synaptonemal complex. Genetica. 2000; 110:277–83. https://doi.org/10.1023/A:1012742717240
Prizon AC, Bruschi DP, Borin-Carvalho LA, Cius A, Barbosa LM, Ruiz HB, Ruiz HB, Zawadzki CH, Fenocchio AS, Portela-Castro ALB. Hidden diversity in the populations of the armored catfish Ancistrus Kner, 1854 (Loricariidae, Hypostominae) from the Paraná River basin revealed by molecular and cytogenetic data. Front Genet. 2017; 8:185. https://doi.org/10.3389/fgene.2017.00185
Prizon AC, Bruschi DP, Gazolla CB, Borin-Carvalho LA, Portela-Castro ALBP. Chromosome spreading of the retrotransposable Rex-3 element and microsatellite repeats in karyotypes of the Ancistrus populations. Zebrafish. 2018; 15:504–14. https://doi.org/10.1089/zeb.2018.1620
Da Rosa R, Giuliano L, Dias AL. Meiotic studies in Teleosts: an approach for studying the behavior of chromosomes and its application. In: Carone S, editor. Teleosts: evolutionary development, diversity and behavioral ecology. New York: Nova Science Publishers; 2014. p.73–96.
Santos AF. Estudos citogenéticos em peixes dos gêneros Bryconops e Moenkhausia (Teleostei, Characidae, Tetragonopterinae). [Master Dissertation]. São Carlos: Universidade Federal de São Carlos/Fundação Universidade do Amazonas; 1999.
Scudeler PES, Diniz D, Wasko AP, Oliveira C, Foresti F. Whole chromosome painting of B chromosomes of the red-eye tetra Moenkhausia sanctaefilomenae (Teleostei, Characidae). Comp Cytogenet. 2015; 9(4):661–69. https://doi.org/10.3897/CompCytogen.v9i4.5460
Sumner AT. A simple technique for demostratings centromeric heterochromatin. Exp Cell Res. 1972; 75(1):304–06. http://doi.org/10.1016/0014-4827(72)90558-7
Teixeira TKSS, Venere PC, Ferreira DC, Mariotto S, Castro JP, Artoni RF, Centofante L. Comparative cytogenetics of Astyanax (Teleostei: Characidae) from the upper Paraguay basin. Neotrop Ichthyol. 2018; 16(1):e170092. http://dx.doi.org/10.1590/1982-0224-20170092
Utsunomia R, Silva DMZA, Ruiz-Ruano FJ, Araya-Jaime C, Pansonato-Alves JC, Scacchetti PC, Hashimoto DT, Oliveira C, Trifonov VA, Porto-Foresti F, Camacho JPM, Foresti F. Uncovering the ancestry of B chromosomes in Moenkhausia sanctaefilomenae (Teleostei, Characidae). PLoS ONE. 2016; 11(3):e0150573. https://doi.org/10.1371/journal.pone.0150573
Authors

![]() Leandro Ranucci1
Leandro Ranucci1 ![]() ,
, ![]() Carlos A. Fernandes1,2,
Carlos A. Fernandes1,2, ![]() Luciana A. Borin-Carvalho1,
Luciana A. Borin-Carvalho1, ![]() Isabel C. Martins-Santos1 and
Isabel C. Martins-Santos1 and ![]() Ana L. B. de Portela-Castro1,2
Ana L. B. de Portela-Castro1,2
[1] Departamento de Biotecnologia, Genética e Biologia Celular, Centro de Ciências Biológicas, Universidade Estadual de Maringá (UEM), Av. Colombo, 5790, 87020-900 Maringá, PR, Brazil. (LR) leandroranucci@gmail.com (corresponding author), (CAF) fxande@gmail.com; (LABC) labcarvalho@uem.br, (ICMS) icmdsantos@hotmail.com, (ALBPC) albpcastro@uem.br.
[2] Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (NUPELIA), Universidade Estadual de Maringá (UEM), Av. Colombo, 5790, 87020-900, Maringá, PR, Brazil.
Authors Contribution
Leandro Ranucci: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Software, Visualization, Writing-original draft, Writing-review and editing.
Carlos Alexandre Fernandes: Conceptualization, Data curation, Supervision, Writing-review and editing.
Luciana Andréia Borin-Carvalho: Methodology, Project administration, Supervision, Visualization, Writing-review and editing.
Isabel Cristina Martins-Santos: Conceptualization, Formal analysis, Methodology, Project administration, Supervision, Validation, Visualization.
Ana Luiza de Brito Portela-Castro: Conceptualization, Data curation, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing-original draft, Writing-review and editing.
Ethical Statement
This study was carried out in strict accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals, approved by the Committee on Ethics of Animal Experiments of the Universidade Estadual de Maringá (License number: protocol 4607140416 – CEUA/UEM). Animals were captured with the permission of the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio; license number 58187–2).
Competing Interests
The authors declare no competing interests.
How to cite this article
Ranucci L, Fernandes CA, Borin-Carvalho LA, Martins-Santos IC, Portela-Castro ALB. Occurrence of euchromatic B chromosomes in natural populations of Moenkhausia bonita and M. forestii (Pisces: Characidae). 2021; 19(4):e210056. https://doi.org/10.1590/1982-0224-2021-0056
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2021 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted August 21, 2021 by Claudio Oliveira
Accepted August 21, 2021 by Claudio Oliveira
![]() Submitted February 22, 2021
Submitted February 22, 2021
![]() Epub November 30, 2021
Epub November 30, 2021