![]() Oscar A. Shibatta1
Oscar A. Shibatta1 ![]() ,
, ![]() Lucas R. Jarduli1,2,
Lucas R. Jarduli1,2, ![]() Vitor P. Abrahão3 and
Vitor P. Abrahão3 and ![]() Lenice Souza-Shibatta1
Lenice Souza-Shibatta1
PDF: EN XML: EN | Supplementary: S1 S2 S3 S4 S5 | Cite this article
Abstract
Lophiosilurus is a monotypic genus represented by L. alexandri, a species endemic to the São Francisco river basin, Brazil. In previous phylogenetic analyses, the genus has been recovered as the sister group of Cephalosilurus. However, few species of Cephalosilurus or few characters were included in those studies. Thus, the current study aims to test the monophyletic hypothesis of the genera Lophiosilurus and Cephalosilurus with a more comprehensive phylogenetic analysis, including all Cephalosilurus species and a representative number of characters. Phylogenetic analyses of 18 terminal taxa (15 ingroups and three outgroups) were conducted based on a combined 75 character matrix, including 70 discrete morphological characters concerning osteology and neuroanatomy, four continuous characters, and the geometric morphometry of the head. The monophyly of the family Pseudopimelodidae was highly supported, and Cephalosilurus is synonymized with Lophiosilurus. The recovered phylogeny of the genus was (L. albomarginatus (L. nigricaudus (L. apurensis (L. fowleri, L. alexandri)))).
Keywords: Biodiversity, Cephalosilurus, Pacamã, Systematics, Taxonomy.
Lophiosilurus é um gênero monotípico representado por L. alexandri, uma espécie endêmica da bacia do rio São Francisco, Brasil. Em análises filogenéticas anteriores, o gênero foi recuperado como grupo irmão de Cephalosilurus. No entanto, poucas espécies de Cephalosilurus ou poucos caracteres foram incluídos nesses estudos. Assim, este estudo tem como objetivo testar a hipótese de monofilia dos gêneros Lophiosilurus e Cephalosilurus, com uma análise filogenética mais abrangente, incluindo todas as espécies de Cephalosilurus e um número representativo de caracteres. As análises filogenéticas de 18 táxons terminais (15 do grupo interno e três grupos externos) foram realizadas com base em uma matriz combinada de 75 caracteres, incluindo 70 caracteres morfológicos discretos de osteologia e neuroanatomia, quatro caracteres contínuos e um de morfometria geométrica da cabeça. A monofilia da família Pseudopimelodidae foi altamente apoiada e Cephalosilurus foi sinonimizado como Lophiosilurus. A filogenia recuperada do gênero foi (L. albomarginatus (L. nigricaudus (L. apurensis (L. fowleri, L. alexandri))).
Palavras-chave: Biodiversidade, Cephalosilurus, Pacamã, Sistemática, Taxonomia.
Introduction
Lophiosilurus Steindachner, 1876 is one of the seven genera of Pseudopimelodidae Fernández-Yépez, Antón, 1966, a catfish family endemic to South America, which comprises 53 valid species (Fricke et al., 2021). Although this is a small family in species number, it has a wide geographic distribution, occurring in drainages west of the Andes in Colombia and Ecuador, the Maracaibo basin, coastal rivers of Guianas and eastern Brazil, and the Orinoco, Amazonas, Araguaia-Tocantins, São Francisco, and La Plata (Paraná, Paraguay, Uruguay) river basins (Shibatta, 2003a; Shibatta, van der Sleen, 2018). Other genera of the family are Pseudopimelodus Bleeker, 1858, Batrochoglanis Gill, 1858, Cephalosilurus Haseman, 1911, Microglanis Eigenmann, 1912, Cruciglanis Ortega-Lara & Lehmann, 2006, and Rhyacoglanis Shibatta & Vari, 2017.
Lophiosilurus is a monotypic genus, represented by L. alexandri Steindachner, 1876, a species endemic to Brazil’s São Francisco river basin. Its body morphology is the most differentiated within the family, especially concerning the broad and prognathous mouth, eye in dorsal position, low dorsal fin, small adipose fin, and depressed and wide-body (Assega et al., 2016). It has psammophilic behavior, burying itself and building nests in the sand. In addition, it feeds on fish, possesses a sit-and-wait strategy, and camouflage color from brown to greyish yellow. This fish has commercial value due to the consistency and flavor of its meat and has been the object of studies for cultivation in fish farming (e.g., Santos et al., 2013). In Brazil, it is popularly known as pacamão or pacamã, and internationally, among aquarists, as the Pacman catfish (Chang, 2013).
Phylogenetic analyses (i.e., Shibatta, 1998; Sullivan et al., 2013; Shibatta, Vari, 2017) have revealed Lophiosilurus as the sister group of Cephalosilurus. Cephalosilurus comprises C. fowleri Haseman, 1911 as type species, also endemic to the São Francisco river basin, C. albomarginatus (Eigenmann, 1912) from the Tukeit river basin, Guyana, C. nigricaudus (Mees, 1974) from the Sipaliwini river basin, Suriname, and C. apurensis (Mees, 1978) from the Apure river basin, Venezuela, and Colombia (Fig. 1). Cephalosilurus is currently diagnosed by its prognathous mouth, color pattern with large dark brown blotches or spots, and depressed body slightly higher than L. alexandri (Shibatta, 1998). Cephalosilurus fowleri is also marketed as pacamã by professional fishers in the São Francisco river basin. However, the biology of this and other species of Cephalosilurus is poorly known.

FIGURE 1 | Geographic distribution of Lophiosilurus species. Lophiosilurus apurensis, MZUEL 6492, 189.7 mm SL, Apure river basin, Venezuela; L. nigricaudus, INPA 21632, 58.1 mm SL, Sipaliwini river basin, Suriname; L. albomarginatus, ROM 61336, 88.4 mm SL, Tukeit river basin, Guyana; L. alexandri, MZUEL 5377, 157.8 mm SL, São Francisco river basin, Brazil; L. fowleri, FMNH 54254, holotype, 301.4 mm SL, São Francisco river basin, Brazil.
In the last 20 years, knowledge of the systematics of Pseudopimelodidae has advanced considerably. Descriptions of new species and genera and new phylogenetic hypotheses have improved our understanding of the family (e.g.,Shibatta, 1998; Shibatta, Benine, 2005; Shibatta, Pavanelli, 2005; Ortega-Lara, Lehmann, 2006; Alcaraz et al., 2008; Jarduli, Shibatta, 2013; Shibatta, 2014; Shibatta, Vari, 2017; Shibatta, 2019). Morphological and molecular studies have established the monophyly of Pseudopimelodidae and its relationships within Pimelodoidea (e.g., Lundberg et al., 1991a; Shibatta, 2003a; Diogo et al., 2004; Ortega-Lara, Lehmann, 2006; Birindelli, Shibatta, 2011; Sullivan et al., 2013; Shibatta, Vari, 2017; Abrahão et al., 2018). Although the monophyly of Pseudopimelodidae is strongly supported, the relationships among its members are only partially resolved. Although different studies have supported the close relationship between Cephalosilurus and Lophiosilurus (e.g., Shibatta, 2003b; Ortega-Lara, Lehmann, 2006; Birindelli, Shibatta, 2011; Sullivan et al., 2013; Shibatta, Vari, 2017; Rangel-Medrano et al., 2020; Silva et al., 2021), they did not include all species in the clade (Fig. 2).
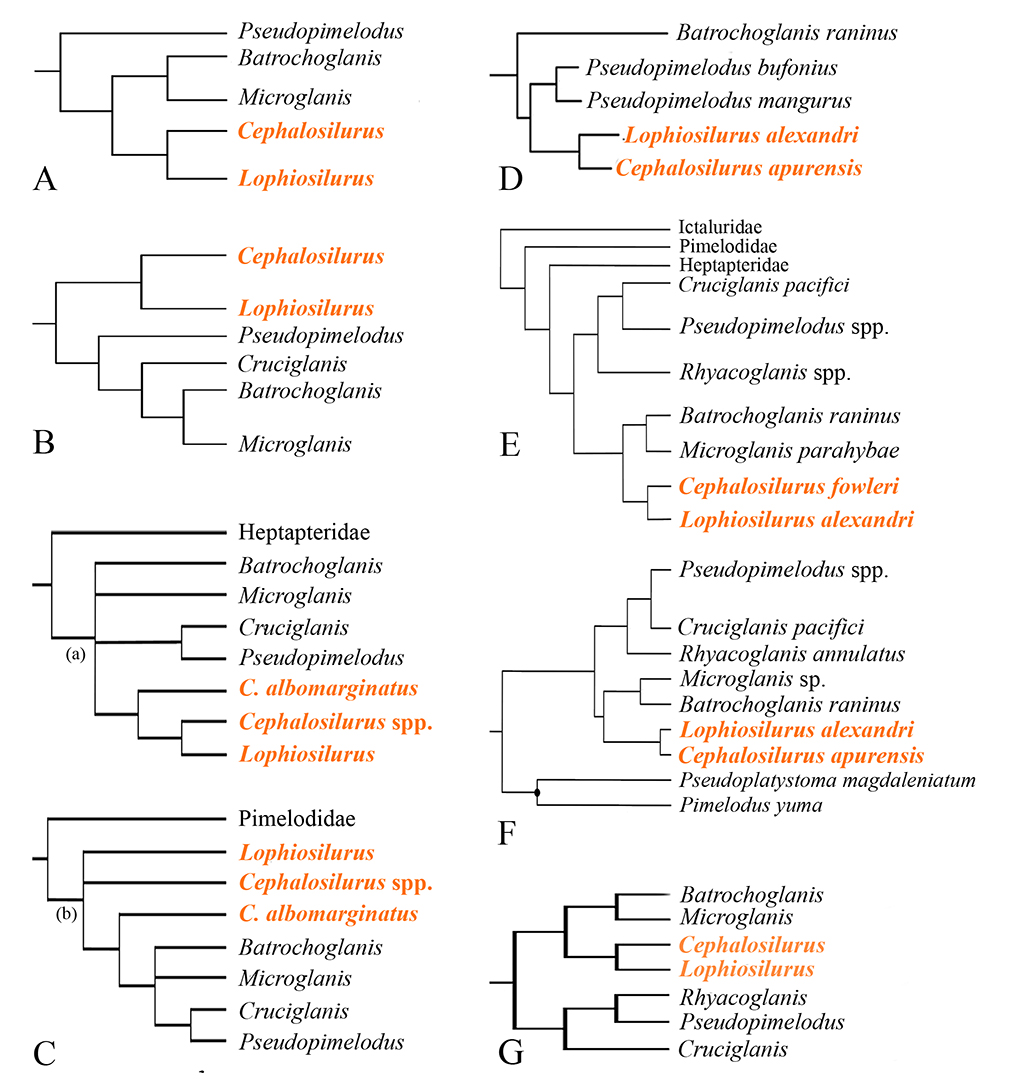
FIGURE 2 | Previous hypothesis of Pseudopimelodidae phylogeny: A. Shibatta (2003b); B. Ortega-Lara, Lehmann (2006); C. Birindelli, Shibatta (2011); D. Sullivan et al. (2013); E. Shibatta, Vari (2017); F. Rangel-Medrano et al. (2020); G. Silva et al. (2021).
The depressed body is the most interesting feature shared by Lophiosilurus and Cephalosilurus that differentiate these genera from remaining pseudopimelodids. Body shape is often only indirectly included in phylogenetic analyses; however, new methods developed in the last two decades allow body shape to be incorporated with discrete and continuous morphological characters (Goloboff et al., 2006; Catalano, Goloboff, 2018). Thus, studies combining different kinds of characters can improve the resolution and support in clades differentiable by body shape, refining our understanding of the evolution of studied groups (González-José et al., 2008; Catalano et al., 2015; Solis-Zurita et al., 2019).
In this contribution, we present a phylogenetic analysis, including all species of Cephalosilurus and Lophiosilurus alexandri, integrating discrete and continuous morphological characters derived from osteology, neuroanatomy, gas bladder morphology, morphometrics, counts, and head shape data. This study aims to test the hypothesis of Cephalosilurus monophyly and propose a new diagnosis for the group.
Material and methods
Characters and taxa. The phylogenetic matrix included 75 morphological characters, 41 of them from Shibatta, Vari (2017), two from Ortega-Lara, Lehmann (2006), ten from Abrahão et al. (2018), and 22 new characters (17 discrete, four continuous, and one from geometric morphometry of head shape landmark configuration). The discrete characters were treated as non-additive, whereas continuous characters (i.e., coordinates of a landmark in space and measurements and counts after standardization) were analyzed in TNT as additive characters, using Farris optimization (Farris, 1970), with the implementation described in Goloboff et al. (2006) as applied in Ferrer et al. (2014). The continuous characters and counts were standardized (0‒1; the maximum possible variation between any two taxa), with the implementation described in Goloboff et al. (2006). In this context, transformation costs are defined as the numerical difference between states. No discretization step (e.g., gap coding methods) is required.
The coding of character number four from Shibatta, Vari (2017) was changed for Cruciglanis pacifici, according to the original description (Ortega-Lara, Lehmann, 2006) and re-examination of the species. The analysis of the neurocranium of Cruciglanis pacifici is based on fig. 4a of Ortega-Lara, Lehmann (2006). Terminal taxa included twelve pseudopimelodid species analyzed by Shibatta, Vari (2017) plus three species of Cephalosilurus (C. albomarginatus, C. apurensis, and C. nigricaudus). Ictalurus punctatus (Rafinesque, 1818) (Ictaluridae), Rhamdia quelen (Quoy, Gaimard, 1824) (Heptapteridae), and Steindachneridion scriptum (Miranda Ribeiro, 1918) (Pimelodidae) were the outgroup taxa, with the former as the root of analysis. The results obtained herein indicate that Cephalosilurus is a junior synonym of Lophiosilurus. Following this, all species of Cephalosilurus will henceforth be treated as Lophiosilurus. Nomenclature of bones follows Arratia (2003a,b) and Birindelli (2014).
Examined material. The following specimens were used in this study (alc = alcohol; c&s = cleared and stained; gm = geometric morphometry): Batrochoglanis raninus (Valenciennes, 1840): MZUSP 23407 (2 c&s: not measured); INPA 1979 (1 alc/gm: 108.0 mm SL); INPA 7343 (1 alc/gm: 65.8 mm SL); INPA 8057 (1 alc/gm: 67.8 mm SL); INPA 19863 (2 alc/gm: 102.2–107.0 mm SL); INPA 34130 (3 alc/gm: 46.2–54.6 mm SL); LIRP 9435 (1 alc/gm: 51.8 mm SL); MZUEL 6035 (1 alc/gm: 77.6 mm SL). MZUSP 23407 (2 alc/gm: 50.7–74.4 mm SL); Batrochoglanis villosus (Eigenmann, 1912): ANSP 135903 (1 alc: 91.3 mm SL); MZUSP 7356 (2 c&s: not measured). Cruciglanis pacifici INCIVA (IMCN) 113 (1 alc/gm: 91.6 mm SL). Ictalurus punctatus: MZUEL 13834 (5 dry skel: not measured); MZUEL 6672 (10 alc/gm: 158.76–257.29 mm SL). Lophiosilurus albomarginatus (Eigenmann, 1912): ROM 61336 (17 alc/10 gm: 29.0–88.4 mm SL); FMNH 53221 (holotype: alc/gm: 75 mm SL); FMNH 53572 (paratype: alc/gm: 68.8 mm SL); FMNH 53222 (8 alc/gm: 21.5–58.5 mm SL); ROM 61482 (2 alc/gm: 50.5–78.8 mm SL). Lophiosilurus alexandri: MZUEL 16469 (1 dry skel: not measured); MZUEL 5377 (3 alc/gm: 120.5–160.5 mm SL); MZUEL 13823 (1 alc/gm: 256.9 mm SL); MZUEL 16486 (1 alc/gm: 279.8 mm SL); MZUEL 19691 (2 alc/gm: 181.0–265.5 mm SL); MZUSP 96276 (1 alc/gm: 70.1 mm SL). Lophiosilurus apurensis: NRM 15995 (1 alc/gm: 201.9 mm SL); MZUEL 6492 (1 alc/gm: 186.0 mm SL); MZUEL 6493 (1 alc/gm: 188.4 mm SL); MZUSP 110996 (1 alc/gm: 180.3 mm SL); MZUSP 11099 (1 alc/gm: 200.2 mm SL). Lophiosilurus fowleri (Hasemann, 1911): ANSP 172158 (2 alc: 39.2–199.5 mm SL); FMNH 54254 (holotype: alc/gm: 301.4 mm SL); MCP 16675 (2 alc/gm: 44.9–128.0 mm SL); MCP 14094 (1 alc/gm: 328.8 mm SL); MCP 14126 (1 alc/gm: 262.7 mm SL); MZUSP 38097 (1 alc/gm: 244.6 mm SL); MZUSP 24647 (1 alc/gm: 277.0 mm SL). Lophiosilurus nigricaudus (Mees, 1974): INPA 21632 (2 alc/gm: 55.4–57.5 mm SL). Microglanis parahybae Steindachner, 1880:MNRJ 15989 (2 c&s: not measured; 5 alc/gm: 30.3–34.2 mm SL); MNRJ 16047 (5 alc.: 29.4–35.8 mm SL; 5 alc/gm: 28.6–38.9 mm SL). Pseudopimelodus bufonius (Valenciennes, 1840): MZUEL 5744 (20 alc/gm: 88.28–260.49 mm SL); MZUEL 07715 (1 alc/gm: 85.77 mm SL); INPA 8058 (1 alc/gm: 68.8 mm SL). Pseudopimelodus mangurus (Valenciennes, 1835): MZUEL 17690 (8 alc.: 112.0–190.4 mm SL); MZUEL 829 (1 alc/gm: 140.8 mm SL); MZUEL 1073 (1 alc/gm: 185.2 mm SL); MZUEL 3681 (1 alc/gm: 97.3 mm SL); MZUEL 6618 (1 alc/gm: 127 mm SL); MZUEL 1477 (2 alc/gm: 87.55–111.7 mm SL); MZUEL 1777 (1 alc/gm: 144, 53 mm SL); MZUEL 38278 (1 alc/gm: 83.55 mm SL); MZUEL 5741 (2 alc/gm: 159.9–208.2 mm SL); MCP 10336 (1 alc/gm: 129.0 mm SL); MCP 12685 (1 alc/gm: 146.1 mm SL); MCP 13087 (1 alc/gm: 173.7 mm SL). Rhamdia quelen: MZUEL 14631 (3 dry skel.: not measured); MZUEL 3845 (15 alc/gm: 104.6–193.3 mm). Rhyacoglanis annulatus Shibatta & Vari, 2017: ANSP 160625 (1 alc/gm: 42.5 mm SL); ANSP 192597 (1 alc/gm: 27.0 mm SL). Rhyacoglanis epiblepsis Shibatta & Vari, 2017:AMNH 40127 (25 paratypes alc/gm: 41.0–54.2 mm SL). Rhyacoglanis paranensis Shibatta & Vari, 2017:MZUEL 6034 (10 alc/gm: 44.6–38.7 mm SL); MZUEL 14121 (1 alc/gm: 89.3 mm SL); MZUEL 12159 (12 alc/gm: 33.7–27.6 mm SL). Rhyacoglanis pulcher Shibatta & Vari, 2017:BMNH 1880.12.8.105–107 (3 syntypes alc/gm: 58.5–68.5 mm SL). Rhyacoglanis seminiger Shibatta & Vari, 2017: LIRP 8042 (9 paratypes alc/gm: 48.3–74.8 mm SL); MZUEL 14123 (2 alc/gm: 60.4–64.8 mm SL); MZUSP 82085 (3 alc/gm: 44.4–70.4 mm SL). Steindachneridion scriptum: MZUEL 866 (1 c&s: 107.5 mm SL; 1 alc/gm: 602.2 mm SL); MZUEL 1482 (1 alc/gm: 163.3 mm SL); MZUEL 16238 (10 alc/gm: 101–125.9 mm SL). For additional material see Shibatta, Vari (2017) and Abrahão et al. (2018). Institutional abbreviations follow Sabaj (2019).
Geometric morphometry. Digital images of the head in the dorsal view of each specimen were taken with a 12.1 Mpixels High-Sensitivity CMOS sensor camera. Fourteen homologous landmarks (Fig. 3) were plotted in two dimensions using TPSDig 2.21 (Rohlf, 2015). The procedures performed to obtain the landmark data follow Catalano et al. (2015). First, all specimens of each species were superimposed using a General Procrustes Analysis (GPA; Gower, 1975; Rohlf, Slice, 1990) to obtain the consensus configurations representing each species shape. The consensus configurations derived from this step represent the shape of each species. Next, the TPSRelw 1.65 program (Rohlf, 2016) was used to obtain the GPA and consensus settings. The consensus configurations for each species were used to define multiple superimpositions using a new GPA. Finally, the multiple superimpositions were incorporated into a matrix of combined characters, generating a combined data set (see Tab. S1), as recommended by Catalano, Goloboff (2018).

FIGURE 3 | Landmark position on Pseudopimelodus mangurus, MZUEL 1073, 185.2 mm SL. 0 = Tip of snout; 1 = Posterior margin of the left anterior nostril; 2= Posterior margin of the right anterior nostril; 3 = Insertion of left maxillary barbel; 4 = Insertion of right maxillary barbel; 5 = Posterior margin of the left posterior nostril; 6 = Posterior margin of the right posterior nostril; 7 = Medial margin of left eye; 8 = Medial margin of right eye; 9 = Most lateral point of the head in the left; 10 = Most lateral point of the head in the right; 11 = Upper end of the left operculum; 12 = Upper end of the right operculum; 13 = Insertion of dorsal fin spine.
Phylogenetic analysis. The combined matrix of discrete, continuous, and landmark data (see Tab. S1) was used to conduct the phylogenetic search under maximum parsimony in TNT program version 1.5 (Goloboff, Catalano, 2016; Catalano, Goloboff, 2018). The phylogenetic analysis was performed following the approach proposed by Catalano et al. (2010) to analyze landmark data. This approach maximizes the degree to which the similarity in landmark position in different taxa can be accounted for by common ancestry (Catalano, Goloboff, 2012). The tree score was given by the sum of the landmark displacements along the tree (Catalano et al., 2015). The tree score and ancestral shapes were established using the algorithms described in Goloboff, Catalano (2011) to optimize landmark data on a tree. This approach sets the optimal ancestral position of each landmark through a heuristic approximation, in which a grid is placed over the space occupied by all the observed locations for a given landmark. Each grid cell is considered a possible state (i.e., position) for the inner nodes. A cost matrix between states is constructed by calculating the distances between the centers of each of the cells. Once each of the observed positions has been assigned to the corresponding cell, the optimal positions are established using a cost matrix algorithm (Sankoff, Rousseau, 1975; Goloboff, 1998) to find the values for the ancestral nodes that minimize ancestor/descendant differences (Catalano et al., 2010). In this study, the landmark optimization was run with the following settings: 8×8 grid of cells; two levels of nested grids; observed landmark positions included as states and a final iterative improvement in the positions. The theoretical basis and algorithms for landmark analysis in a parsimony context are described in Catalano et al. (2010), Goloboff, Catalano (2011), Catalano, Goloboff (2012), and Goloboff, Catalano (2016).
The search strategy consisted of 5000 Random Addition Sequences (RAS = Wagner trees) followed by TBR (Tree Bisection and Reconnection algorithm) and holding 100 trees per replication. Branches supporting values were calculated using symmetric resampling (change probability set at 33%), with 5000 replications in TNT. Differential costs do not distort symmetric resampling, and the probability of increasing the weight of a character equals the probability of decreasing it (Goloboff et al., 2003; Goloboff et al., 2008). The retention index and the consistency index of the geometric morphometry character were computed according to Ospina-Garcés, Luna (2017).
Results
Continuous characters description. The values of continuous characters 1 to 4 of each species are shown in Tab. S2.
1. Gill-rakers on first branchial arch; counts: minimum 2; maximum 27 (CI 0.84; RI 0.87). In Ictalurus punctatus, Steindachneridion scriptum, and Rhamdia quelen, the number of gill-rakers varies from 11 to 18 (median 15). In Cruciglanis, Pseudopimelodus, and Rhyacoglanis, the number is lower, ranging from 2 to 10 (median 6), as well as in Batrochoglanis and Microglanis from 6 to 11 (median 11). In Lophiosilurus species, the gill-rakers on the first branchial arch range from 9 to 27 (median 13.5), with the impressive highest number in Lophiosilurus apurensis with 27. Although most catfish species present less than 15 gill rakers on the first branchial arch, some Siluridae and Doradoidea present higher numbers (Birindelli, 2014). In Pseudopimelodidae, lower numbers of gill-rakers are shared by Cruciglanis, Pseudopimelodus, and Rhyacoglanis and are proposed as a synapomorphy for this group.
2. Dorsal-fin spine; length: minimum 5% of SL; maximum 21.3% of SL (CI 0.50; RI 0.10). In Ictalurus punctatus, Rhamdia quelen, and Steindachneridion scriptum, the ossified portion of the dorsal-fin spine is elongated, with size ranging from 8.2 to 16.2% of SL. The dorsal-fin spine of Pseudopimelodidae is usually longer, varying between 8.7 to 21.3% of SL. However, in Lophiosilurus alexandri, the dorsal-fin spine is extremely short, with the dorsal-fin spine length varying from 5 to 5.9% of SL.
3. Body depth relative to body width; distance between pectoral girdles: minimum 33.2%; maximum 116.6% (CI 0.45; RI 0.43). The body of Rhamdia quelen (101.4%) and Ictalurus punctatus (111.6‒116.6%) is the deepest among the examined material. The body is slightly shallower in Steindachneridion scriptum, Cruciglanis pacifici, Microglanis parahybae, Pseudopimelodus, and Rhyacoglanis (ranging from 61% to 92.9%). Lophiosilurus albomarginatus, L. apurensis, L. fowleri, and Batrochoglanis raninus exhibit body depth proportions from 58.6 to 67.7%. Lophiosilurus alexandri and L. nigricauda are the most depressed, with body widths of 33.2‒47.8% and 48.7%, respectively. Assega et al. (2016) observed negative allometry in the body depth of L. alexandri throughout its development and discussed the phylogenetic meaning of body depth. According to those authors, a relatively deep body is plesiomorphic, and a depressed body is apomorphic.
4. Adipose-fin base; length: minimum 4.6% of SL; maximum 43.4% of SL (CI 0.54; RI 0.04). The most extended adipose-fin base length is observed in Rhamdia quelen (37‒43.5%), and the shortest is observed in Lophiosilurus alexandri (4.6‒7.44%). Most Pseudopimelodidae, Ictalurus punctatus, and Steindachneridion scriptum have intermediate values. A large adipose fin base is present in Dyplomystes nahuelbutaensis since small juveniles (Lundberg et al., 2004), corroborating the plesiomorphic condition of character in Siluriformes.
Discrete characters description.
Skin and dermal structures. Characters 5 to 8 herein are described as 1 to 4 in Shibatta, Vari (2017).
5. Unculiferous tubercles; (0) absent; (1) present (Shibatta, Vari, 2017: fig. 14) (CI 1.00; RI 1.00).
6. Unculiferous epidermal structures; degree of development: (0) little developed; (1) well developed (CI 0.33; RI 0.60).
7. Skin covering pectoral-fin spine; thickness: (0) thin; (1) thick (CI 0.33; RI 0.71).
8. Axillary pore: (0) present; (1) absent (CI 0.50; RI 0.50).

FIGURE 4 | Dorsal view of left maxillary bone; anterior up: A.Steindachneridion scriptum, MZUEL 19302; B.Rhamdia quelen, MZUEL 14631; C.Lophiosilurus apurensis, MBUCV-V-15379. Scale bars = 1 mm.
Pigmentation. Characters 9 to 17 herein are described as 5 to 13 in Shibatta, Vari (2017).
9. Light blotch on the cheek: (0) absent; (1) present (CI 1.00; RI 1.00).
10. Dark brown mask well defined on the head:(0) absent; (1) present (CI 1.00; RI 1.00).
11. Dark band on predorsal region: (0) absent; (1) present (CI 0.33; RI 0.60).
12. Fusion of dark predorsal and subdorsal bands: (0) absent (1) partial, (2) complete (CI 1.00; RI 1.00).
13. Dark bands on the trunk and caudal peduncle: (0) absent throughout ontogeny; (1) present at some point during ontogeny (CI 1.00; RI 1.00).
14. Fusion of dark subdorsal and subadipose bands: (0) absent; (1) present (CI 0.33; RI 0.00).
15. Small scattered spots on the lateral surface of the body: (0) limited in number, widely dispersed; (1) numerous, closely dispersed (CI 0.33; RI 0.00).
16. Dark caudal-fin stripe: (0) absent; (1) present (CI 0.50; RI 0.80).
17. Caudal fin; pigmentation: (0) middle of the dark caudal-fin stripe without convergence with dark caudal-peduncle band; (1) middle stripe convergent with caudal-peduncle band; (2) entire fin dark (CI 1.00; RI 1.00).
18. Caudal fin dark-spot; configuration (char. 61 from Ortega-Lara, Lehman, 2006): (0) without dark spot; (1) with a dark spot at its base and fused with the peduncular spot, covering the anterior three-quarters of its length, and distal rim hyaline. (CI 0.33; RI ‒). Among species analyzed, state 1 is shared by Cruciglanis pacifici (Ortega-Lara, Lehmann, 2006: fig. 5), Lophiosilurus albomarginatus, and L. nigricaudus.
Jaws. Characters 19 to 20 herein are described as 13 to 14 in Shibatta, Vari (2017).
19. Premaxilla; bifurcated dorsolateral process:(0) absent; (1) present (autapomorphy).
20. Premaxillary tooth plate; form: (0) not posterolaterally extended; (1) posterolaterally extended (autapomorphy).
21. Posterior portion of maxillary bone; shape: (0) pointed; (1) irregular; (2) rounded (Fig. 4) (CI 1.00; RI 1.00). In Ictalurus punctatus and Steindachneridion scriptum, the posterior extremity of maxillary bone is pointed (state 0), and in Rhamdia quelen, it is irregular (state 1). In almost all species of Pseudopimelodidae, the maxillary bone can be posteriorly pointed (state 0). However, in Lophiosilurus albomarginatus, L. alexandri, L. apurensis, and L. fowleri, the maxillary bone is posteriorly semi-circular (state 2).
Head and neurocranium. Characters 22 to 25 herein are described as 15 to 18 in Shibatta, Vari (2017).
22. Anterior nostril; location: (0) distant from the margin of mouth; (1) near the margin of mouth (CI 1.00; RI 1.00).
23. Vomer: (0) present; (1) absent (Ortega-Lara, Lehmann, 2006: fig. 3) (CI 1.00; RI 1.00).
24. Lateral ethmoid-autopalatine joint; length: (0) abridged, autopalatine only touching the lateral ethmoid (Shibatta, 2019: fig. 2); (1) elongated, about 1/6 of autopalatine articulated with lateral ethmoid (autapomorphy).
25. Autopalatine; shape: (0) depth similar anteriorly and posteriorly; (1) depth greater posteriorly; spoon-shaped (Shibatta, 2019: fig. 3) (CI 1.00; RI 1.00).
26. Medial constriction on mesethmoid: (0) present; (1) absent (Fig. 5) (CI 1.00; RI 1.00). In Ictalurus punctatus, Rhamdia quelen, and Steindachneridion scriptum, the mesethmoid has a constriction, widening posteriorly (state 0). This plesiomorphic condition is also observed in Lophiosilurus albomarginatus, Batrochoglanis raninus, Microglanis parahybae, Pseudopimelodus mangurus, Rhyacoglanis paranensis, R. epiblepsis, and Cruciglanis pacifici. However, in L. alexandri, L. apurensis, and L. fowleri, constriction does not occur, and the mesethmoid is nearly trapezoidal, wider anteriorly than posteriorly (state 2).
27. Anterior frontal fontanel; shape: (0) as a groove (1) ellipsoid (Fig. 5) (CI 0.50; RI 0.50). In Ictalurus punctatus, Rhamdia quelen, and Steindachneridion scriptum, the anterior frontal fontanel is narrow and long as a groove (state 0) as in many Pseudopimelodidae. However, in Batrochoglanis raninus, Microglanis parahybae, and Lophiosilurus albomarginatus, it is wide with an ellipsoid shape (state 1).
28. Orbitosphenoid flap-like projection: (0) absent; (1) present (CI 0.50; RI ‒). In Ictalurus punctatus and Rhamdia quelen, the orbitosphenoid does not has a ventral flap-like projection. Nevertheless, it is present in Steindachneridion scriptum and all species of Pseudopimelodidae.
29. Orbitosphenoid flap-like projection; anterior width: (0) narrower than parasphenoid width; (1) similar to parasphenoid width; (2) broader than parasphenoid width (CI 0.66; RI 0.66). In Lophiosilurus alexandri, L. fowleri, and Batrochoglanis raninus, the orbitosphenoid is narrower than parasphenoid (state 0). The orbitosphenoid and parasphenoid widths are similar in Steindachneridion scriptum and Pseudopimelodus mangurus (state 1). In Microglanis parahybae and Rhyacoglanis epiblepsis,and R. paranaensis, the width is more extensive (state 2). This character is inapplicable in Ictalurus punctatus and Rhamdia quelen since the orbitosphenoid does not have a lateral flap-like projection in the ventral region of the neurocranium lateral to the parasphenoid.
30. Frontal; shape: (0) trapezoidal with wide anterior; (1) trapezoidal with broad posterior; (2) rectangular (Fig. 5) (CI 0.67; RI 0.67). In Siluriformes, the shape of the frontal bone is quite variable. In Ictalurus punctatus, the frontal bone has a greater width in the anterior region, at the lateral ethmoid level, than in the posterior region at sphenotic level (state 0). In Rhamdia quelen, Steindachneridion scriptum, Batrochoglanis raninus, Microglanis parahybae, Lophiosilurus albomarginatus, Rhyacoglanis paranensis, and R. epiblepsis, the posterior region is wider than the anterior one (state 1). The width of the anterior frontal area in Pseudopimelodus mangurus, Lophiosilurus alexandri, L. fowleri, and L. apurensis is similar to that of the posterior region (state 2).
31. Anterior frontal fontanel; configuration (char. 59 from Ortega-Lara, Lehman, 2006): (0) not reaching posteriorly to the level of the infraorbital sensory canal opening of the sphenotic bone; (1) reaching that level. (CI 0.50; RI 0.67). In Ictalurus punctatus, Rhamdia quelen, Steindachneridion scriptum, and Cruciglanis pacifici, the anterior frontal fontanel reaches the level of the infraorbital canal opening of the sphenotic bone (state 0). In other analyzed species of Pseudopimelodidae, the anterior frontal fontanel is shorter (state 1) (Fig. 5; Ortega-Lara, Lehmann, 2006: fig. 4).
32. Parietal-supraoccipital; shape: (0) trapezoidal; (1) square (Fig. 5) (CI 0.25; RI 0.40). In Ictalurus punctatus and Rhamdia quelen, the parietal-supraoccipital is trapezoidal and wider posteriorly (state 0). Species of Lophiosilurus and Pseudopimelodus mangurus exhibit this same condition. A quadrangular form without posterior expansion occurs in Steindachneridion scriptum, Batrochoglanis raninus, Microglanis parahybae, Cruciglanis pacifici, and Rhyacoglanis epiblepsis (state 1).
33. Infraorbitals; shape: (0) gradually expanding posteriorly; (1) wide uniformly (Shibatta, 2019: fig. 2) (CI 0.50; RI 0.67). In Ictalurus punctatus, Rhamdia quelen, Steindachneridion scriptum, and Lophiosilurus alexandri, the infra-orbital bones gradually expand posteriorly (state 0). The width is consistent from second to fifth (state 1) in all other pseudopimelodids.
34. Ectopterygoid: (0) present; (1) absent (CI 1.00; RI ‒). The ectopterygoid is present and homologous across most Siluriformes groups, and this condition is observed in Steindachneridion scriptum and all species of Pseudopimelodidae examined herein (state 0). However, in Rhamdia quelen, this bone is absent (state 1).
35. Ectopterygoid posterior prolongation; medial shape: (0) straight; (1) concave; (2) convex (Fig. 6) (CI 0.60, RI 0.33). In Steindachneridion scriptum, the medial region of posterior prolongation of the ectopterygoid is straight (state 0). This character is inapplicable in Ictalurus punctatus and Rhamdia quelen by the absence of ectopterygoid. Also, in Lophiosilurus albomarginatus and L. apurensis, the ectopterygoid is present, but the prolongation is absent and coded as inapplicable. In Batrochoglanis raninus, Lophiosilurus alexandri, L. fowleri, Microglanis parahybae, Pseudopimelodus mangurus, Rhyacoglanis paranensis, and R. epiblepsis, the prolongation is concave (state 1). In Cruciglanis pacifici, it is convex (state 2).
36. Anterior portion of entopterygoid; shape: (0) approximately straight; (1) narrowing anteriorly to rounded tip (Fig. 6) (CI 1.00, RI 1.00). The entopterygoid has the anterior region almost straight and broad as the posterior one in Steindachneridion scriptum and Rhamdia quelen (state 0). In Pseudopimelodidae, there is a tip in the anterior region (state 1).
37. Posterior portion of entopterygoid; shape: (0) almost straight, without posterior projection; (1) with a posterior projection; (2) sharp tip (Fig. 6) (CI 0.67, RI 0.67). In Steindachneridion scriptum, Rhamdia quelen, Batrochoglanis raninus, Microglanis parahybae, Pseudopimelodus mangurus, Rhyacoglanis paranensis, and R. epiblepsis, the posterior region of the entopterygoid has a relatively uniform width along its length and a truncated posterior portion without projection (state 0). This structure ends in a small projection in Cruciglanis pacifici, Lophiosilurus albomarginatus, and L. apurensis (stage 1). In Lophiosilurus alexandri and L. fowleri, the entopterygoid has a prolonged sharp tip in the posterior region (state 2).

FIGURE 5 | Neurocranium in dorsal view: A.Lophiosilurus alexandri, MZUEL 5542; B.L. fowleri, MZUEL 12095; C.L. apurensis, MBUCV-V-15379; D.L. albomarginatus, ROM 61336; E.Batrochoglanis raninus, MZUSP 23407; F.Microglanis parahybae, MNRJ 15989; G. Pseudopimelodus mangurus, MZUEL 12937; H.Rhyacoglanis paranensis, MZUEL 6034. Bones in yellow and cartilage in bluish grey. Scale bars = 5 mm.
Hyoid and gill arches. Characters 38 to 42 herein are described as 19 to 23 in Shibatta, Vari (2017).
38. Dorsal hypohyal: (0) present; (1) absent (Shibatta, 2019: fig. 6) (CI 1.00; RI 1.00).
39. Second basibranchial; shape: (0) without lateral processes; (1) with lateral processes resulting in a cruciform shape (Ortega-Lara, Lehmann, 2006: fig. 2) (autapomorphy).
40. Metapterygoid; shape: (0) longer than wide; (1) as long as wide (Lundberg et al. 1991a: fig. 9) (CI 1.00; RI 1.00).
41. Fenestra in quadrate: (0) absent; (1) present (Shibatta, 2019: fig. 5) (CI 1.00; RI 1.00).
42. Gill rakers on first branchial arch; distribution: (0) along the anterior margin of ceratobranchial; (1) restricted to the posterior region of anterior margin of ceratobranchial (Shibatta, 2019: fig. 7) (CI 0.33; RI 0.60).
43. Gill rakers on first branchial arch; shape: (0) conical and elongated, without bifurcation; (1) bifurcated or even branched (Fig. 7) (CI 1.00; RI 1.00). In most species of Pseudopimelodidae, in Ictalurus punctatus, Rhamdia quelen, and Steindachneridion scriptum, the gill rakers are elongated and not bifurcated (state 0). Bifurcated gill rakers are found only in Lophiosilurus albomarginatus, L. alexandri, L. apurensis, and L. fowleri within Pseudopimelodidae (state 1).

FIGURE 6 | Ventral view of left ectopterygoid and entopterygoid bones; anterior up: A.Batrochoglanis raninus, MZUSP 23407; B.Lophiosilurus albomarginatus, ROM 61336; C.Lophiosilurus alexandri, MZUEL 5542.
Appendicular and axial skeleton. Characters 44 to 49 and 51 to 55 herein are described as 24 to 34 in Shibatta, Vari (2017).
44. Mesocoracoid arch; shape: (0) elongate (Shibatta, 2019: fig. 8); (1) triangular (CI 1.00; RI 1.00).
45. Posterior cleithral process; length: (0) elongate, more than half continuing posteriorly to the margin of opercular membrane; (1) short, broadly covered by an opercular membrane (CI 0.50; RI 0.80).
46. Branched pectoral-fin rays; number: (0) 8 or more; (1) 7; (2) 6; (3) 5 (CI 0.75; RI 0.80).
47. Serrae along anterior and posterior margins of pectoral-fin spine; height: (0) shorter anteriorly; (1) all of the same approximate length (CI 0.40; RI 0.50).
48. Serrae along the anterior margin of pectoral-fin spine; distribution: (0) across entire margin; (1) restricted to base (CI 0.50; RI 0.00).
49. Tip of pectoral-fin spine; shape: (0) pointed; (1) bifurcated (CI 0.33; RI 0.67).
50. Anterior process of parapophysis of the fourth vertebra; shape: (0) straight, not curved; (1) ventrally curved; (2) laterally swelled (Fig. 8) (CI 1.00; RI 1.00). In Ictalurus punctatus, the anterior transverse process of the fourth vertebra is almost entirely straight, as is Steindachneridion scriptum and Rhamdia quelen (state 0). This vertebra is ventrally curved in Pseudopimelodus mangurus and Rhyacoglanis paranensis, forming a folding structure (state 1). In Lophiosilurus, Batrochoglanis raninus, and Microglanis parahybae, only the lateral tip is swelled (state 2).
51. Fifth and sixth centra; joint: (0) not interdigitating; (1) deeply interdigitating (Lundberg et al., 1991b: fig. 12) (autapomorphy).
52. Vertebrae; number: (0) 46 or more; (1) 36‒45; (2) 30‒35 (CI 0.50; RI 0.71).
53. Dorsal-fin spine; serration: (0) smooth anteriorly; (1) serrated anteriorly (CI 0.33; RI 0.71).
54. Caudal fin; shape: (0) two pointed lobes; (1) two rounded lobes; (2) two subtle rounded lobes (emarginated); (3) no distinct lobes (rounded) (CI 0.60; RI 0.71).
55. Caudal-fin lobes; length: (0) lobes approximately equal; (1) ventral lobe longer; (2) dorsal lobe longer (CI 0.40; RI 0.67).
56. Dorsal lobe of the caudal skeleton; composition: (0) hypurals 3, 4, and 5 free; (1) hypural 5 free, hypurals 3 and 4 fused; (2) hypurals 3, 4, 5 fused (Fig. 9) (CI 0.67; RI –). In Ictalurus punctatus and Steindachneridion scriptum, the caudal bones have hypurals 3, 4, and 5 free (state 0). State 1 is observed in Rhamdia quelen, Pseudopimelodus mangurus, Batrochoglanis raninus, Microglanis parahybae, Lophiosilurus apurensis,and L. fowleri. State 2 occurs in L. alexandri.
57. Ventral lobe of caudal skeleton; composition: (0) parhypural and hypural 1 free; (1) parhypural and hypural 1 fused (Fig. 9) (CI 1.00; RI 1.00). In Ictalurus punctatus, Rhamdia quelen, and Steindachneridion scriptum, all elements of ventral caudal bones are free (state 0). Conversely, state 1 is observed in all examined species of Pseudopimelodidae.

FIGURE 7 | Shape of gill rakers on the first ceratobranchial; lateral view of left selected portion; anterior to left: A.Steindachneridion scriptum, MZUEL 19302; B.Lophiosilurus alexandri, MZUEL 5542. Scale bars = 5 mm.

FIGURE 8 | Ventral view of complex vertebrae; anterior up: A.Ictalurus punctatus, MZUEL 13834; B.Steindachneridion scriptum, MZUEL, 19302; C.Pseudopimelodus mangurus, MZUEL 17286; D.Lophiosilurus alexandri, MZUEL 5542. Scale bar = 5 mm.

FIGURE 9 | Lateral view of caudal bones; anterior to left: A.Steindachneridion scriptum, MZUEL 19302; B.Rhamdia quelen, MZUEL 11664; C. Pseudopimelodus mangurus, MZUEL 17286; D.Lophiosilurus alexandri, MZUEL 13288. Scale bars = 5 mm.
Gas bladder. Characters 58 to 61 herein are described as 35 to 38 in Shibatta, Vari (2017).
58. Gas bladder; shape: (0) heart-shaped; (1) dumbbell-shaped (Birindelli, Shibatta, 2011: fig. 1) (CI 1.00; RI 1.00).
59. Constrictor muscle of gas bladder; presence: (0) absent; (1) present (CI 0.50; RI 0.67)
60. Lateral trabeculae on internal T-shaped gas bladder septum; presence: (0) absent; (1) present (CI 0.50; RI 0.80).
61. Pseudotympanum opening; size: (0) large, height about twice the diameter of the eye; (1) small, height approximately equal to the diameter of the eye (Birindelli, Shibatta, 2011: fig. 2) (CI 0.50; RI 0.83).
Laterosensory system and maxillary barbel. Characters 62 to 64 herein are described as 39 to 41 in Shibatta, Vari (2017).
62. Lateral line; length: (0) complete; (1) incomplete but long; extending beyond vertical through adipose fin; (2) incomplete and short; falling short of vertical through adipose fin (Ruiz, Shibatta, 2010: fig. 3) (CI 0.40; RI 0.00).
63. Cephalic lateral line; arrangement: (0) unbranched (Shibatta, 2019: fig. 14); (1) branched (Lundberg et al., 1991b: fig. 11) (autapomorphy).
64. Maxillary barbel; length: (0) long; extending beyond vertical through the dorsal-fin origin; (1) short; falling short of vertical through the dorsal-fin origin (CI 1.00; RI 1.00).
Gross brain morphology. The following characters (65 to 74) were obtained from Abrahão et al. (2018; corresponding numbers in parenthesis) and renumbered here; a detailed discussion on each character can be obtained from that paper. In addition, Abrahão et al. (2018) generalized character states for family or genus after examining several of their representatives. The same was followed in this work.
65. Intumescence on lobus facialis, position (char. 1): (0) oriented along the entire lateral margin of each lobe; (1) located along the anterolateral portion; (2) anteriorly placed (Fig. 10) (CI 100; RI 100). In Ictaluridae and Heptapteridae, the lobus facialis appears to have two regions longitudinally subdivided into two halves (state 0). Pimelodidae, Pseudopimelodus, Microglanis, Batrochoglanis, Cruciglanis, and Rhyacoglanis the lobus facialis anterolaterally, occupying half the width of each lobe in the posterior portion and extending to the anteromedial part (state 1). In Lophiosilurus, the intumescence on lobus facialis is anteriorly placed (state 2).
66. Posterior portion of the lateral line lobe; configuration (char. 2): (0) reaching the half-length of the lobus facialis; (1) reaching the anterior part of the lobus vagi (Fig. 10) (CI 100; RI 100). In Ictaluridae and Heptapteridae, the prolongation of the posterior portion of the lateral-line lobe ends abruptly and reaches only half the length of the lateral margin of lobus facialis (state 0). In Pimelodidae and Pseudopimelodidae, the posterior portion of the lateral-line lobe has a tail-shaped prolongation that extends beyond the division between lobus facialis and lobus vagi (state 1).
67. Saccus vasculosus; relative size (char. 3): (0) smaller than the pituitary gland; (1) same size or larger than the pituitary gland (CI 1.00; RI 1.00). In Ictaluridae and Heptapteridae, saccus vasculosus is smaller than the pituitary gland. On the other hand, the saccus vasculosus is more conspicuous in Pimelodidae and Pseudopimelodidae, with the same size or larger than the pituitary gland (state 1).
68. Conspicuous bulge on the anterior portion of lobus vagi (char. 4): (0) present; (1) absent (Fig. 10) (CI 1.00; RI 1.00). In Ictaluridae, Heptapteridae, and Pimelodidae, there is a prominent bulge on the anterior portion of the lobus vagi (state 0). In all examined Pseudopimelodidae, such bulge is less conspicuous or absent (state 1).
69. Cerebellum; general shape (char. 5): (0) oval-shaped; (1) somewhat triangular (Fig. 10). (CI 1.00; RI 1.00). In Ictaluridae, Heptapteridae, and Pimelodidae, the cerebellum has an oval shape with the posterior region rounded (state 0). The same condition is observed in Pseudopimelodus, Cruciglanis, and Rhyacoglanis. In Batrochoglanis, Microglanis, Cephalosilurus, and Lophiosilurus, the cerebellum is somewhat triangular with the posterior margin straight, and an anterior margin rounded or at an acute angle (state 1).
70. Anterior portion of the cerebellum; prolongation (char. 6): (0) extending beyond the half-length of the telencephalon; (1) extends until the posterior portion of telencephalon; (2) extends as far as boundary between mesencephalon and telencephalon (also exhibiting habenula in the dorsal portion of the diencephalon) (Fig. 10) (CI 1.00; RI 1.00). In Ictaluridae, Heptapteridae, and Pimelodidae, the cerebellum is longer, with its anterior portion prolonged beyond the half-length of the telencephalon (state 0). In Pseudopimelodus, Cruciglanis, and Rhyacoglanis, the cerebellum has intermediate size, with the anterior portion reaching until the telencephalon’s posterior portion (state 1). In Batrochoglanis, Microglanis, and Lophiosilurus, the cerebellum’s anterior portion is prolonged as far as the boundary between mesencephalon and telencephalon but does not meet the latter and exhibiting habenula in the dorsal portion of the diencephalon (state 2).
71. Lobus facialis; shape (char. 7): (0) elongated; (1) reduced (Fig. 10) (CI 1.00; RI 1.00). In Ictaluridae, Heptapteridae, and Pimelodidae, the lobus facialis is elongated, comprising more than half the length of the lateral-line lobe (state 0). It is the same in several Pseudopimelodidae species, but in Lophiosilurus, the lobus facialis is reduced, comprising less than half the length of the lateral-line lobe (state 1).
72. Cerebellum; shape (char. 8): (0) elongated sagittal, length greater than its width; (1) wide coronal, width greater than its length (CI 1.00; RI 1.00). The cerebellum is elongated in all species examined herein, but in Batrochoglanis and Microglanis, the cerebellum is broader as state 1 (Fig. 10).
73. Cerebellum; relative volume (char. 9): (0) more voluminous than telencephalon; (1) less voluminous than telencephalon (Fig. 10) (CI 1.00; RI 1.00). In all examined species, the cerebellum is the broadest subdivision of the brain (state 0). However, in Batrochoglanis and Microglanis, the cerebellum is less voluminous than the telencephalon (state 1).
74. Crest on the dorsal surface of the cerebellum (char. 10): (0) absent; (1) present (Fig. 10) (CI 1.00; RI 1.00). There is no crest along the cerebellum’s dorsal surface (state 0) of all examined species, except in Rhyacoglanis, where lateral depressions form the crest in the cerebellum’s parasagittal plane (state 1).

FIGURE 10 | Dorsal view of brain; anterior up: A.Rhamdia quelen, MZUEL 6036, 222.4 mm SL; B.Rhyacoglanis paranensis, MZUEL 6034, 39.2 mm SL; C.Cruciglanis pacifici, MCP non-catalogued, 142.4 mm SL; D.Lophiosilurus alexandri, MZUEL 5377, 58.7 mm SL.
Geometric morphometry character.One character was recovered from geometric morphometry analysis:
75. Shape configuration of the head. Fourteen landmarks of the head of Pseudopimelodidae and outgroups were analyzed (Fig. 3). The set of landmarks from the head forms a configuration that represents one character in the phylogenetic analysis, and the contribution of this configuration is equivalent to that of one discrete character. Each change of position in homologous landmarks represents a different state of this character. The landmarks under parsimony allow visualization of their displacement along the branches (Fig. 11; see Figs. S3 and S4) (CI 0.92; RI 0.83).

FIGURE 11 | Inferred phylogeny and landmark mapping (numbers) on the head of Lophiosilurus species. The arrow indicates the ancestral configuration. Enlarged image of Lophiosilurus alexandri in the box (grey lines = ancestral shape; red lines = reconstructed shape, blue lines = landmark position changing to the corresponding node in the ancestor).
Phylogenetic analysis. The combined morphological data parsimony analysis recovered a single most parsimonious tree (Fig. 12), CI = 0.63, and RI = 0.76, with the best score hitting 5000 times out of 5000 resamplings. Pseudopimelodidae was recovered as a monophyletic group and supported by 13 characters. Within Pseudopimelodidae, two lineages were recovered, one composed by (Rhyacoglanis (Cruciglanis, Pseudopimelodus)), with five characters and support of 63%, and the other by (Lophiosilurus (Microglanis, Batrochoglanis)), with nine characters and support of 76%. The characters supporting all the branches are presented in Fig. 12, and the change in character states is in Tab. S5.

FIGURE 12 | Resampled tree (CI = 63; RI = 76; length (best score) = 148.87) of Pseudopimelodidae derived from combined 75 characters matrix data set, with synapomorphies above and support values (symmetric resampling) under the branches. Pseudopimelodidae subfamilies names according to Silva et al. (2021).
Although the wideness of the head can be confirmed with the naked eye, the phylogenetic significance and subtle changes are visualized more easily by geometric morphometry analysis. The change in this species is evidenced by comparing the landmark displacements to the ancestral shape, obtained by applying spatial optimization on the landmarks of each configuration of the head shape. The widening of head shape, easily observed by the lateral displacement of landmarks 9 and 10, shows different states in the family Pseudopimelodidae, reaching the most significant displacement in Lophiosilurus alexandri (Fig. 11). Other states of this character (head shape configuration) in the Lophiosilurus clade are very subtle to point out, such as the posterior displacement of anterior nostrils (landmarks 1 and 2) and medial displacement of the eyes (landmarks 7 and 8). In Lophiosilurus apurensis, the sister species of L. alexandri and L. fowleri, the eyes are more laterally displaced (landmarks 7 and 8), and the predorsal region is shortened (anterior displacement of landmark 13). The morphology differences in the head of the species based on landmarks are presented in Figs. S3 and S4.
Following the phylogenetic analysis, the morphological characterization of the expanded Lophiosilurus genus is:
Lophiosilurus Steindachner, 1876
Lophiosilurus alexandri Steindachner, 1876:154 [106], Pl. 15 (original description).
Pseudopimelodus agassizii Steindachner, 1880:61 (unnecessary name for replacement of Lophiosilurus alexandri Steindachner, 1876).
Cephalosilurus Haseman, 1911:317, Pl. (description of Cephalosilurus fowleri, the type of the genus).
Diagnosis. The following characters support Lophiosilurus: 1) reduction of body depth relative to pectoral girdle width (ch. 3: 65.2–76.9%>63–65%); 2) parieto-supraoccipital trapezoidal, posteriorly expanded (ch. 51: 1>0); 3) posterior portion of entopterygoid with a posterior projection (ch. 56: 0>1); 4) posterior portion of maxillary bone semi-circular (ch. 57: 0>2); 5) gill rakers on first branchial arch bifurcated or even branched (ch. 58: 0>1); 6) intumescence on lobus facialis anteriorly placed (ch. 63: 1>2); 7) lobus facialis reduced (ch. 69: 0>1); 8) snout shortening, increase in the anterior and posterior nostrils distance, interorbital distance, head width, and shortening of the predorsal region, as observed in the displacement of the landmarks 0, 1, 2, 5, 6, 8, 9, 11, and 13 (ch. 75). Other characters of the genus are: small to large body size, between 116 and 490 mm SL; body depressed; large, angled head at the edges of the mouth; usually prognathous; anterior nostrils located on the upper lips; lateral margin of the premaxillary dentigerous plate projected posteriorly; pectoral-fin spine covered by a thin skin; pectoral fin 6 to 7 branched rays; axillary pore present; pelvic fin starting at vertical through the base of the fourth or of the penultimate branched ray of dorsal fin; caudal fin rounded or with the upper lobe longer; lateral line complete; variable color pattern, with dark subdorsal and subadipose dark brown blotches, irregularly shaped or with only a few spots scattered across the trunk; the color of caudal fin ranging from light brown without dark spots to completely dark brown or black.
Remarks. Lophiosilurus alexandri presents the most depressed head and several autapomorphies that kept it a separate genus among Pseudopimelodidae. However, according to the relationship of Lophiosilurus alexandri, as shown in Fig. 9, Cephalosilurus is a synonym of Lophiosilurus. According to the International Code of Zoological Nomenclature (ICZN, 1999; Art. 23), Lophiosilurus Steindachner, 1876, prioritizes Cephalosilurus Haseman, 1911. Therefore, all species of Cephalosilurus must be rearranged accordingly (Tab. 1): Lophiosilurus albomarginatus (Eigenmann, 1912), Lophiosilurus apurensis (Mees, 1978), Lophiosilurus fowleri (Haseman, 1911), and Lophiosilurus nigricaudus (Mees, 1974) new combination.
TABLE 1 | Key taxonomic publications with changes in the namesof Lophiosilurus species. † undescribed fossil species identified by Lundberg et al. (2010).
Nominal species | Mees (1974) | Shibatta
(2003b) | Current contribution |
Cephalosilurus
fowleri Haseman, 1911 | Pseudopimelodus
fowleri | Cephalosilurus
fowleri | Lophiosilurus
fowleri (Haseman, 1911) |
Cf. Cephalosilurus sp.† | ‒ | ‒ | Lophiosilurus sp. |
Lophiosilurus
alexandri Steindachner, 1876 | ‒ | Lophsiosilurus
alexandri | Lophiosilurus
alexandri Steindachner, 1876 |
Pseudopimelodus
albomarginatus Eigenmann, 1912 | Pseudopimelodus
albomarginatus | Cephalosilurus
albomarginatus | Lophiosilurus
albomarginatus (Eigenmann, 1912) |
Pseudopimelodus
apurensis Mees, 1978 | ‒ | Cephalosilurus
apurensis | Lophiosilurus
apurensis (Mees, 1978) |
Pseudopimelodus
nigricauda Mees, 1974 | Pseudopimelodus
nigricauda | Cephalosilurus
nigricaudus | Lophiosilurus
nigricaudus (Mees, 1974) |
The range of distribution of extant species of Lophiosilurus includes the São Francisco river basin, Brazil (L. alexandri and L. fowleri), Orinoco river basin, Venezuela and Colombia (L. apurensis), coastal rivers of Guyana (L. albomarginatus), and Suriname (L. nigricaudus) (Fig. 1). Lundberg et al. (2010) identified some dorsal-fin spine fossils from Amazon Neogene as belonging to a species of Pseudopimelodidae. According to the authors, the spines are “robust, with a distinctively short shaft and broad base”. Some are from the current Acre river basin in Peru, and others are from the Cuenca basin, Ecuador. The formation where they occur is from the middle-late Miocene. The authors further note that the spines were previously identified as Pseudopimelodus (Lundberg, 1998) but could belong to Cephalosilurus. The authors’ spine photo (Lundberg et al., 2010, p. 282) is from a relatively large specimen (we estimated the spine’s length from the lateral point of greatest width to the apical extremity = 26 mm). The largest width of the spine is nearly 50% of that length. This proportion is not found in any living species of Pseudopimelodidae (L. alexandri (43.4% (n = 1); L. fowleri = 42.1% (n = 1); L. apurensis = 38.4% (n = 1); L. albomarginatus = 36.4‒38.7 (n = 2); Pseudopimelodus mangurus (36.4‒37.2% (n = 2)), but the shape of the spine connects this fossil to L. fowleri and L. alexandri. Therefore, due to its differences in morphometric proportions, it could be an undescribed species of Lophiosilurus. This new species of Lophiosilurus from the fossil records expands the distribution range of the genus.
Discussion
Pseudopimelodidae phylogeny. The monophyly of Pseudopimelodidae was highly supported (100%), corroborating previous hypotheses (Lundberg et al., 1991a; Shibatta, 1998; Shibatta, 2003b; Ortega-Lara, Lehmann, 2006; Birindelli, Shibatta 2011; Sullivan et al., 2013; Shibatta, Vari, 2017; Rangel-Medrano et al., 2020; Fig. 2). However, intergeneric relationships were quite variable in all these analyses. Lundberg et al. (1991a) and Shibatta (1998; 2003b) recovered Pseudopimelodus as the sister group of other genera (Fig. 2A). Ortega-Lara, Lemann (2006) had the Lophiosilurus clade as the sister of all other genera (Fig. 2B). Sullivan et al. (2013) supported Batrochoglanis raninus as the sister group of all other pseudopimelodids (Fig. 2D). The hypotheses published by Shibatta, Vari (2017), and Silva et al. (2021) were quite similar but differed in the relationships among Cruciglanis, Pseudopimelodus, and Rhyacoglanis (Figs. 2E, and G). This study corroborates the phylogenetic hypothesis of Pseudopimelodidae genera presented by Shibatta, Vari (2017).
Although they recovered different relationships among genera, all those studies supported a close relationship between Cephalosilurus and Lophiosilurus. Birindelli, Shibatta (2011) was the first to include almost all species of Lophiosilurus in the phylogenetic analysis (Fig. 2C) and observed that L. apurensis, L. fowleri, L. nigricaudus, and L. alexandri were unique among pseudopimelodids by possessing the constrictor muscle of the gas bladder. The absence of the constrictor muscle in L. albomarginatus was the main character to point out the polyphyly of the Cephalosilurus. However, according to our present analysis, the absence of the gas bladder muscle is a plesiomorphy.
This contribution points to the evolution of two clades within Pseudopimelodidae, each with strong support (77 and 61%). A different relationship within Pseudopimelodidae was found in a previous study by Sullivan et al. (2013; Fig. 2D). The authors analyzed five species of four genera of Pseudopimelodidae in a study on the phylogeny of the Pimelodoidea superfamily, using rag1 and rag2 nuclear and 12S and 16S rDNA mitochondrial genes. The hypothesis of Batrochoglanis raninus as sister to the clade ((Pseudopimelodus mangurus, P. bufonius), (Lophiosilurus alexandri, Cephalosilurus apurensis)) was strongly supported both in parsimony and Bayesian analysis. However, the incomplete taxon sampling (Microglanis, Rhyacoglanis, and Cruciglanis were missing) may have impaired the consistency of these relationships. More recently, Rangel-Medrano et al. (2020) examined the mitochondrial cox1 (or COI) and nuclear rag2 genes, including Cruciglanis, Microglanis, and Rhyacoglanis in the analysis, and presented a phylogenetic tree that corroborates the two lineages evidenced by Shibatta, Vari (2017). According to the dated phylogeny of those authors, these lineages may have diverged in the upper Eocene, just under 40 mya. Both lineages were recently recovered by Silva et al. (2021) using nuclear loci of ultraconserved elements analysis. Silva et al. (2021) recognized one of the clades as subfamily Pseudopimelodinae and described the other as subfamily Batrochoglaninae.
Geometric morphometry and evolution of head shape in Pseudopimelodidae. In this study, the geometric morphometric character had low homoplasy levels and can be interpreted as phylogenetically informative. The results of the present study clearly show that landmark data can be an important source of evidence for phylogenetic analysis. The analysis of morphological data also provides a potential source of alternative evidence that can be used to complement other data. Phylogenetic morphological studies generate a vast amount of biological knowledge presented in a systematized and ordered manner, representing relevant data for systematists (Catalano et al., 2015).
The head shape configuration had high values of consistency and retention indices. Although morphometry is widely explored in taxonomy, it has only recently been used in phylogenetic analyses (Adams et al., 2004; Gonzalez-José et al., 2008; Klingenberg, Gidaszewski, 2010; Catalano, Torres, 2017; Ospina-Garcés, Luna, 2017). The mapping of different states (changes in the landmark positions) with geometric morphometry (GM) along the branches provided a new evidence source. In addition, this is the first time a landmark data configuration has been combined with other kinds of characters to study Neotropical freshwater fish systematics. Solis-Zurita et al. (2019) recommend that morphometric characters be combined with different characters to infer homology and phylogenetic groups. Several authors have noted the advantages of including GM in phylogenetic inferences (David, Laurin, 1996; Larson, 2005; Gonzalez-José et al., 2008; Catalano et al., 2015; Catalano, Torres, 2017; Ospina-Garcés, Luna, 2017).
The phylogenetic analysis using geometric morphometry helped support the Pimelodoidea clade, an essential contribution to the phylogeny proposed by Shibatta, Vari (2017), who found no synapomorphy for that clade. Using the geometric morphometric character resulted in a wholly solved phylogeny in the Rhyacoglanis clade. A small phylogenetic change was observed in Rhyacoglanis paranensis, compared to the hypothesis proposed by Shibatta, Vari (2017). In that study, R. paranensis was found to be the sister group of all other congeners, while in the current hypothesis, the species is the sister of R. epiblepsis and R. annulatus. However, studies on the Rhyacoglanis species from the Amazon basin are lacking, and the possible inclusion of new species may alter the configuration of currently known phylogenetic relationships.
A global analysis of landmark positions helped reveal subtle changes which are challenging to see by direct observation of specimens. Furthermore, it was not necessary to create discrete categories with arbitrary limits to demonstrate the variation in the shape of the heads of the studied species. This new data source enables valuable information on the species’ shape (Adams et al., 2004; Klingenberg, Gidaszewski, 2010). The shape is one of the most important and easily measured phenotype elements and expresses the interaction of many, if not most, genes (Ollier et al., 2006; Covain et al., 2008).
All Cephalosilurus species inclusion allowed testing the genus monophyly and their reallocation into Lophiosilurus. A better understanding of the Lophiosilurus species morphology allowed us to recognize a new fossil species representing an extinct species that occurred in the Amazon basin, filling an essential geographical gap in the genus’s distribution. Using continuous characters instead of discretizing them allowed us to better understand the group’s evolution without creating subjective categories. The congruence between the phylogenetic relationships of the Pseudopimelodidae genera obtained with the exclusive use of morphological characters and those obtained by other authors through molecular characters shows the information power of both types of data.
Acknowledgments
We thank José Luis O. Birindelli for valuable suggestions to improve the manuscript. To the II International Symposium’s scientific committee on Phylogeny and Classification of Neotropical Fishes for their invitation to present part of this study at the event. We also thank Scott Shaefer (AMNH), John Lundberg, Mark Sabaj (ANSP), Lucia R. Py Daniel (INPA), Flavio Bockmann (LIRP), Mary Burridge (ROM), Marcelo de Britto (MNRJ), Mario de Pinna, Alex Datovo (MZUSP), and Richard Vari (USNM), for the loan of specimens. Programa de Pós-Graduação em Ciências Biológicas da Universidade Estadual de Londrina and Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for supporting LRJ’s thesis. Santiago Catalano and Sandra Ospina-Garcés assisted us in the use of TNT for landmark data analysis. This study was partially financed by a grant from the All Catfish Species Inventory (NSF DEB‒0315963) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; proc. 200280/2014‒7). CNPq research productivity grant to OAS (303685/2018‒2). VPA was supported by CAPES (grant# 88887.318624/2019‒00) and FAPESP (proc. 2014/11397‒1; 2015/26804‒4; 2016/19075‒9; 2017/17957‒7). To Maria Luisa Sarmento Soares and Brian Sidlauskas for the manuscript revision.
References
Abrahão VP, Pupo FM, Shibatta OA. Comparative brain gross morphology of the Neotropical catfish family Pseudopimelodidae (Osteichthyes, Ostariophysi, Siluriformes), with phylogenetic implications. Zool J Linn Soc. 2018; 184(3):750–72. https://doi.org/10.1093/zoolinnean/zly011
Adams DC, Rohlf FJ, Slice DE. Geometric morphometrics: ten years of progress following the “revolution”. Ital J Zool. 2004; 71(1):5–16. https://doi.org/10.1080/11250000409356545
Alcaraz HSV, Graça WJ, Shibatta OA. Microglanis carlae, a new species of bumblebee catfish (Siluriformes: Pseudopimelodidae) from the río Paraguay basin in Paraguay. Neotrop Ichthyol. 2008; 6(3):425–32. https://doi.org/10.1590/S1679-62252008000300016
Arratia G. Catfish head skeleton – an overview. In: Arratia G, Kapoor BG, Chardon M, Diogo R, editors. Catfishes. Enfield, England: Science Publishers; 2003a. p.3–46.
Arratia G. The siluriform postcranial skeleton – an overview. In: Arratia G, Kapoor BG, Chardon M, Diogo R, editors. Catfishes. Enfield, England: Science Publishers; 2003b. p.121–57.
Assega FM, Birindelli JLO, Bialetzki A, Shibatta OA. External morphology of Lophiosilurus alexandri Steindachner, 1876 during early stages of development, and its implications for the evolution of Pseudopimelodidae (Siluriformes). PloS ONE. 2016; 11:e0153123. https://doi.org/10.1371/journal.pone.0153123
Birindelli JLO. Phylogenetic relationships of the South American Doradoidea (Ostariophysi: Siluriformes). Neotrop Ichthyol. 2014; 12(3):451–564. http://dx.doi.org/10.1590/1982-0224-20120027
Birindelli JLO, Shibatta OA. Morphology of the gas bladder in bumblebee catfishes (Siluriformes, Pseudopimelodidae). J Morphol. 2011; 272(7):890–96. https://doi.org/10.1002/jmor.10961
Catalano SA, Ercoli M, Prevosti F. The more, the better: The use of multiple landmark configurations to solve the phylogenetic relationships in Musteloids. Syst Biol. 2015; 64(2):294–306. https://doi.org/10.1093/sysbio/syu107
Catalano SA, Goloboff PA, Giannini NP. Phylogenetic morphometrics (I): the use of landmark data in a phylogenetic framework. Cladistics. 2010; 26(5):539–49. https://doi.org/10.1111/j.1096-0031.2010.00302.x
Catalano SA, Goloboff PA. Simultaneously mapping and superimposing landmark configurations with parsimony as optimality criterion. Syst Biol. 2012; 61(3):392–400. https://doi.org/10.1093/sysbio/syr119
Catalano AS, Goloboff PA. A guide for the analysis of continuous and landmark characters in TNT (Tree Analysis using New Technologies). Technical Report; 2018. https://doi.org/10.13140/RG.2.2.23797.27360
Catalano SA, Torres A. Phylogenetic inference based on landmark data in 41 empirical datasets. Zool Scr. 2017; 46(1):1–11. https://doi.org/10.1111/zsc.12186
Chang I. Breeding success with the Pac-man catfish, Lophiosilurus alexandri. Amazonas. 2013; 2:74–78.
Covain RS, Dray S, Fisch-Muller S, Montoya-Burgos JI. Assessing phylogenetic dependence of morphological traits using co-inertia prior to investigate character evolution in Loricariinae catfishes. Mol Phylogenet Evol. 2008; 46(3):986–1002. https://doi.org/10.1016/j.ympev.2007.12.015
David B, Laurin B. Morphometrics and cladistics: measuring phylogeny in the sea urchin Echinocardium. Evolution. 1996; 50(1):348–59. https://doi.org/10.1111/j.1558-5646.1996.tb04498.x
Diogo R, Chardon M, Vanderwalle P. Osteology and myology of the cephalic region and pectoral girdle of Batrochoglanis raninus, with a discussion on the synapomorphies and phylogenetic relationship of the Pseudopimelodinae and Pimelodidae (Teleostei: Siluriformes). Anim Biol. 2004; 54(3):262–80. http://hdl.handle.net/2268/246296
Farris J. Methods for computing Wagner trees. Syst Biol. 1970; 19(1):83–92. https://doi.org/10.1093/sysbio/19.1.83
Ferrer J, Wingert JM, Malabarba LR. Description of a new species and phylogenetic analysis of the subtribe Cynopoecilina, including continuous characters without discretization (Cyprinodontiformes: Rivulidae). Zool J Linn Soc. 2014; 172(4):846–66. https://doi.org/10.1111/zoj.12190
Fricke R, Eschmeyer WN, Van der Laan R, editors. Eschmeyer’s catalog of fishes: genera, species, references. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Accessed 04 Nov 2021.
Goloboff PA. Tree searches under Sankoff parsimony. Cladistics. 1998; 14(3):229–37. https://doi.org/10.1006/clad.1998.0068
Goloboff PA, Catalano SA. Phylogenetic morphometrics (II): algorithms for landmark optimization. Cladistics. 2011; 27(1):42–51. https://doi.org/10.1111/j.1096-0031.2010.00318.x
Goloboff PA, Catalano SA. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics. 2016; 32(3):221–38. https://doi.org/10.1111/cla.12160
Goloboff PA, Farris JS, Källersjö M, Oxelman B, Szumik CA. Improvements to resampling measures of group support. Cladistics. 2003; 19(4):324–32. https://doi.org/10.1111/j.1096-0031.2003.tb00376.x
Goloboff PA, Farris JS, Nixon KC. TNT, a free program for phylogenetic analysis. Cladistics. 2008; 24(5):774–86. https://doi.org/10.1111/j.1096-0031.2008.00217.x
Goloboff PA, Mattoni CI, Quinteros AS. Continuous characters analyzed as such. Cladistics. 2006; 22(6):589–601. https://doi.org/10.1111/j.1096-0031.2006.00122.x
González-José R, Escapa I, Neves WA, Cúneo R, Pucciarelli HM. Cladistic analysis of continuous modularized traits provides phylogenetic signals in Homo evolution. Nature. 2008; 453:775. https://doi.org/10.1038/nature06891
Gower JC. Generalized Procrustes analysis. Psychometrika. 1975; 40:33–51. https://doi.org/10.1007/BF02291478
Haseman JD. Descriptions of some new species of fishes and miscellaneous notes on others obtained during the expedition of the Carnegie Museum to central South America. Ann Carnegie Mus. 1911; 7:315–28, pls. 46–52.
International Code of Zoological Nomenclature (ICZN). The International Trust for Zoological Nomenclature, London; 1999. Available from: https://www.iczn.org/the-code/the-international-code-of-zoological-nomenclature/the-code-online.
Jarduli LR, Shibatta OA. Description of a new species of Microglanis (Siluriformes: Pseudopimelodidae) from the Amazon basin, Amazonas State, Brazil. Neotrop Ichthyol. 2013; 11(3):507–12. https://doi.org/10.1590/S1679-62252013000300004
Klingenberg P, Gidaszewski NA. Testing and quantifying phylogenetic signals and homoplasy in morphometric data. Syst Biol. 2010; 59(3):245–61. https://doi.org/10.1093/sysbio/syp106
Larson PM. Ontogeny, phylogeny, and morphology in anuran larvae: morphometric analysis of cranial development and evolution in Rana tadpoles (Anura: Ranidae). J Morphol. 2005; 264(1):34–52. https://doi.org/10.1002/jmor.10313
Lundberg JG, Berra TM, Friel JP. First description of small juveniles of the primitive catfish Diplomystes (Siluriformes: Diplomystidae). Ichthyol Explor Freshw. 2004; 15(1):71–82. Available from: https://mansfield.osu.edu/assets/mansfield/tberra/pdf/Diplomystes.pdf
Lundberg JG, Bornbusch AH, Mago-Leccia F. Gladioglanis conquistador n. sp. from Ecuador with diagnoses of the subfamilies Rhamdiinae Bleeker and Pseudopimelodinae n. subf. (Siluriformes: Pimelodidae). Copeia. 1991a; 1991(1):190–209. https://doi.org/10.2307/1446263
Lundberg JG, Mago-Leccia F, Nass P. Exallodontus aguanai, a new genus and species of Pimelodidae (Pisces: Siluriformes) from deep river channels of South America and delimitation of the subfamily Pimelodinae. Proc Biol Soc Wash. 1991b; 104(4):840–69.
Lundberg JG, Sabaj MH, Dahdul WM, Aguilera OA. The Amazonian Neogene fish fauna. In: Hoorn C, Wesselingh FP, editors. Amazônia: Landscape and species evolution: A look into de past. Hoboken, NY: Blackwell Publishing Ltd.; 2010. p.281–301.
Mees GF. The Auchenipteridae and Pimelodidae of Suriname (Pisces, Nematognathi). Zoolog Verh (Leiden). 1974; 132:1–256. Available from: https://repository.naturalis.nl/pub/317592
Ollier S, Couteron P, Chessel D. Orthonormal transform to decompose the variance of a life-history trait across a phylogenetic tree. Biometrics. 2006; 62(2):471–77. https://doi.org/10.1111/j.1541-0420.2005.00497.x
Ortega-Lara A, Lehmann PA. Cruciglanis, a new genus of Pseudopimelodid catfish (Ostariophysi: Siluriformes) with description of a new species from the Colombian Pacific coast. Neotrop Ichthyol. 2006; 4(2):147–56. https://doi.org/10.1590/S1679-62252006000200002
Ospina-Garcés SM, Luna ED. Phylogenetic analysis of landmark data and the morphological evolution of cranial shape and diets in species of Myotis (Chiroptera: Vespertilionidae). Zoomorphology. 2017; 136:251–65. https://doi.org/10.1007/s00435-017-0345-z
Rangel-Medrano J, Ortega-Lara A, Márquez EJ. Ancient genetic divergence in bumblebee catfish genus Pseudopimelodus (Pseudopimelodidae: Siluriformes) from northwestern South America. PeerJ. 2020; 8:e9028. http://doi.org/10.7717/peerj.9028
Rohlf FJ. tpsDig v.2.21. Distributed by the author, Department of Ecology and Evolution. New York: State University of New York, Stony Brook; 2015.
Rohlf FJ. tpsRelw v.1.65. Distributed by the author, Department of Ecology and Evolution. New York: State University of New York, Stony Brook; 2016.
Rohlf FJ, Slice D. Extensions of the Procrustes method for the optimal superimposition of landmarks. Syst Zool. 1990; 39(1):40–59. https://doi.org/10.2307/2992207
Ruiz WBG, Shibatta OAS. A new species of Microglanis (Siluriformes, Pseudopimelodidae) from lower rio Tocantins basin, Pará, Brazil, with description of superficial neuromasts and pores of lateral line system. Zootaxa. 2010; 2632:53–66. https://doi.org/10.11646/zootaxa.2632.1.3
Sabaj MH. Standard symbolic codes for institutional resource collections in herpetology and ichthyology: an online reference. Version 7.1; 2019. Available from: https://asih.org/standard-symbolic-codes/about-symbolic-codes
Sankoff D, Rousseau P. Locating the vertices of a Steiner tree in an arbitrary space. Math Program. 1975; 9:240–46. https://doi.org/10.1007/BF01681346
Solis-Zurita C, De Luna E, González D. Phylogenetic relationships in the Sceloporus variabilis (Squamata: Phrynosomatidae) complex based on three molecular markers, continuous characters and geometric morphometric data. Zool Scr. 2019; 48(4):419–39. https://doi.org/10.1111/zsc.12349
Santos HB, Sampaio EV, Arantes FP, Sato Y. Induced spawning and reproductive variables of the catfish Lophiosilurus alexandri Steindachner, 1876 (Siluriformes: Pseudopimelodidae). Neotrop Ichthyol. 2013; 11(3):607–14. https://doi.org/10.1590/S1679-62252013000300014
Shibatta OA. Sistemática e evolução da família Pseudopimelodidae (Ostariophysi, Siluriformes), com a revisão taxonômica do gênero Pseudopimelodus.[PhD Thesis]. São Carlos: Universidade Federal de São Carlos; 1998.
Shibatta OA. Phylogeny and classification of ‘Pimelodidae’. In: Arratia G, Kapoor BG, Chardon M, Diogo R, editors. Catfishes. Enfield, England: Science Publishers; 2003a. p.385–400.
Shibatta OA. Family Pseudopimelodidae (Bumblebee catfishes, dwarf marbled catfishes). In: Reis RE, Kullander SO, Ferraris Jr CJ, organizers. Check List of the Freshwater Fishes of South and Central America Porto Alegre: Edipucrs; 2003b. p.401–405.
Shibatta AO, Benine RC. A new species of Microglanis (Siluriformes: Pseudopimelodidae) from upper rio Paraná basin, Brazil. Neotrop Ichthyol. 2005; 3(4):579–85. https://doi.org/10.1590/S1679-62252005000400015
Shibatta OA, Pavanelli CS. Description of a new Batrochoglanis species (Siluriformes, Pseudopimelodidae) from the rio Paraguai basin, State of Mato Grosso, Brazil. Zootaxa. 2005; 1092(1):21–30. https://doi.org/10.11646/zootaxa.1092.1.2
Shibatta OA. A new species of Microglanis (Siluriformes: Pseudopimelodidae) from the upper rio Tocantins basin, Goiás State, Central Brazil. Neotrop Ichthyol. 2014; 12(1):81–87. https://doi.org/10.1590/S1679-62252014000100008
Shibatta OA. New species of bumblebee catfish of the genus Batrochoglanis Gill, 1858 (Siluriformes: Pseudopimelodidae) from the Aripuanã river basin, Mato Grosso, Brazil.Zootaxa, 2019; 4674(2):243–63. https://doi.org/10.11646/zootaxa.4674.2.6
Shibatta OA, van der Sleen P. Family Pseudopimelodidae-bumblebee catfishes, dwarf-marbled catfishes. In: van der Sleen P, Albert J, editors. Field Guide to the fishes of the Amazon, Orinoco & Guianas New Jersey: Princeton University Press; 2018. p.308–10.
Shibatta OA, Vari RP. A new genus of Neotropical rheophilic catfishes, with four new species (Teleostei: Siluriformes: Pseudopimelodidae). Neotrop Ichthyol. 2017; 15(2):e160132. https://doi.org/10.1590/1982-0224-20160132
Silva GSC, Melo B, Roxo FF, Ochoa LE, Shibatta OA, Sabaj MH, Oliveira C. Phylogenomics of the bumblebee catfishes (Siluriformes: Pseudopimelodidae) using ultraconserved elements. Jour Zool Syst Evol Res. Forthcoming 2021. http://doi.org/10.1111/jzs.12513
Steindachner F. Ichthyologische Beiträge (V). [Subtitles i-v.]. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften. Math Naturwiss Kl. 1876; 74:49–240, pls. 1–15.
Steindachner F. Zur Fisch-Fauna des Cauca und der Flüsse bei Guayaquil. Denkschriften der Kaiserlichen Akademie der Wissenschaften in Wien, Math Naturwiss Kl. 1880; 42:55–104, pls. 1–9.
Sullivan JP, Muriel-Cunha J, Lundberg JG. Phylogenetic relationships and molecular dating of the major groups of catfishes of the Neotropical superfamily Pimelodoidea (Teleostei, Siluriformes). P Acad Nat Sci Phila. 2013; 162:89–110. https://www.jstor.org/stable/42751950
Authors

![]() Oscar A. Shibatta1
Oscar A. Shibatta1 ![]() ,
, ![]() Lucas R. Jarduli1,2,
Lucas R. Jarduli1,2, ![]() Vitor P. Abrahão3 and
Vitor P. Abrahão3 and ![]() Lenice Souza-Shibatta1
Lenice Souza-Shibatta1
[1] Programa de Pós-Graduação em Ciências Biológicas, Departamento de Biologia Animal e Vegetal, Centro de Ciências Biológicas, Universidade Estadual de Londrina, 86057-970 Londrina, PR, Brazil. (OAS) shibatta@uel.br (corresponding author), (LSS) lenicesouza@hotmail.com.
[2] Centro Universitário das Faculdades Integradas de Ourinhos, 19909-100 Ourinhos, SP, Brazil. (LRJ) lucasjarduli@gmail.com.
[3] Programa de Pós-Graduação em Biodiversidade e Evolução, Instituto de Biologia, Universidade Federal da Bahia, Rua Barão de Geremoabo, 668, Ondina, 40170-115 Salvador, BA, Brazil. (VPA) vitorabrahao32@gmail.com.
Authors Contribution
Oscar Akio Shibatta: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Lucas Ribeiro Jarduli: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing-original draft, Writing-review and editing.
Vitor Pimenta Abrahão: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing-original draft, Writing-review and editing.
Lenice Souza-Shibatta: Conceptualization, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing-original draft, Writing-review and editing.
Ethical Statement
Specimens examined in this study were available in collections.
Competing Interests
The authors declare no competing interest
How to cite this article
Shibatta OA, Jarduli LR, Abrahão VP, Souza-Shibatta L. Phylogeny of the Neotropical Pacman catfish genus Lophiosilurus (Siluriformes: Pseudopimelodidae). Neotrop Ichthyol. 2021; 19(4):e210040. https://doi.org/10.1590/1982-0224-2021-0040
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2021 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted August 5, 2021 by Gloria Arratia
Accepted August 5, 2021 by Gloria Arratia
![]() Submitted February 1, 2021
Submitted February 1, 2021
![]() Epub December 10, 2021
Epub December 10, 2021