![]() André Lincoln B. Magalhães1
André Lincoln B. Magalhães1![]() ,
,![]() Luis Artur V. Bezerra2,
Luis Artur V. Bezerra2,![]() Vanessa S. Daga3,
Vanessa S. Daga3,![]() Fernando M. Pelicice4,
Fernando M. Pelicice4,![]() Jean R. S. Vitule3 and
Jean R. S. Vitule3 and![]() Marcelo F. G. Brito5
Marcelo F. G. Brito5
PDF: EN XML: EN | Supplementary: S1 S2 S3 | Cite this article
Abstract
This study evaluated fish beta diversity in six headwater creeks located in the area affected by the largest ornamental aquaculture center implemented in the Minas Gerais State, southeastern Brazil. We sampled fish assemblages in 2017 and 2018 to investigate changes in assemblage structure (species richness and beta diversity), comparing these data with the historic species pool. We recorded 60 fish species, of which 16 were native and 44 non-native with 19 translocated, and 25 exotic. The exotics Poecilia reticulata, Xiphophorus maculatus, X. variatus, Danio rerio, and Misgurnus anguillicaudatus were the most widely distributed in the headwater creeks. The Contamination Index showed that most creeks had high proportional contamination by exotic species (above 60%). Beta diversity increased from historical to contemporary periods in all creeks due to the introduction and differential colonization pressure of several non-native translocated and exotic species, indicating biotic differentiation. Temperature and number of ponds were the main preditors of change in beta diversity in the headwater creeks during the contemporary period. In summary, we observed that invaders have induced substantial changes to fish communities under influence of environmental filters. Our results support the hipothesis that aquaculture is a main driver of fish non-native fish introduction and native biodiversity loss in the Neotropics.
Keywords: Aquaculture,Aquarium trade,Beta diversity, Escapes, Invasive species.
Este estudo avaliou a diversidade beta de peixes em seis riachos de cabeceira localizados em uma área afetada pelo maior centro de aqüicultura ornamental do Brasil, localizado em Minas Gerais. Amostramos assembleias de peixes em 2017 e 2018 para investigar mudanças na estrutura (riqueza de espécies e diversidade beta), comparando esses dados com tendências históricas de composição de comunidades. Registramos 60 espécies de peixes, sendo 16 nativos e 44 não nativos: 19 translocados e 25 exóticos. Os exóticos Poecilia reticulata, Xiphophorus maculatus, X. variatus, Danio rerio e Misgurnus anguillicaudatus foram os mais distribuídos nos riachos. O Índice de Contaminação mostrou que a maioria dos riachos apresentou alta contaminação proporcional por espécies exóticas (acima de 60%). A diversidade beta aumentou do período histórico para o contemporâneo em todos os riachos devido à introdução e pressão de colonização de várias espécies não-nativas translocadas e exóticas, indicando diferenciação biótica. Temperatura da água e número de tanques de piscicultura foram os principais fatores de mudança na diversidade beta dos riachos no período contemporâneo. Os não-nativos induziram mudanças em nível de comunidades e sob influência de variáveis ambientais. Os resultados mostram que a aquicultura é um dos principais vetores da introdução de peixes não-nativos e perda de biodiversidade nativa nos Neotrópicos.
Palavras-chave: Aquicultura, Comércio de aquários, Diversidade beta, Escapes, Espécies invasoras.
Introduction
The aquarium fish industry is both a major economic activity and an important pathway for the introduction of non-native species around the world (Rixon et al., 2005; Ishikawa, Tachihara, 2014; Lockwood et al., 2019; Olden et al., 2020). Among the vertebrate groups internationally marketed, freshwater fish contribute with more than 5,000 species (Bush et al., 2014; Hulme, 2015; Islami et al., 2017). In the last twenty years, the global aquarium trade (i.e., freshwater and marine fish) generated about US$ 3 billion/year (FAO, 2000) and an equipment and literature industry that exceeded US$ 15 billion (Meyers, 2001). The global aquarium trade has grown steadily, followed closely by environmental consequences. The introduction of non-native organisms, either intentional (e.g., aquarium dumping) or accidental (e.g., escapes from fish farms), have been a direct consequence of this growing industry. It has caused, for example, the invasion of more than 100 non-native freshwater aquarium fish (NFAF, hereafter) worldwide (Mendoza et al., 2015; Magalhães et al., 2017; Lockwood et al., 2019).
The outgrowth of ornamental fish farms in Brazil has supplied more than 500,000 residential aquaria with 26 million fish, contributing to rank fish among the most traded pets in the country (Magalhães et al., 2017). Consequently, the Brazilian aquarium trade has become a major pathway for the introduction of NFAF species (Frehse et al., 2016; Magalhães et al., 2017, 2019; Lima Junior et al., 2018), especially via escapes from aquaculture ponds. However, unlike traditional aquaculture, where a few species are intensively cultivated, such as Nile tilapia Oreochromis niloticus (Linnaeus, 1758), and common carp Cyprinus carpio Linnaeus, 1758 (Brito et al., 2020), in ornamental aquaculture several different fish species from different continents (i.e., non-native exotic), and across ecoregions within a country (i.e., non-native translocated; Vitule et al., 2019) may be raised in a single or multiple localities, with the risk of causing multiple disturbances in the resident biota. In fact, studies have reported changes in natural communities associated with the growing invasion of aquarium fish, including the disruption of the native community structure, and biotic homogenization or differentiation (Magalhães, 2006; Magalhães et al., 2020a; Padial et al., 2020).
Non-native fish species have already significantly affected Neotropical fish diversity, particularly species distribution, and community structure (Vitule et al., 2019; Brito et al., 2020; Magalhães et al., 2020a). Non-native species promote multiple disturbances (e.g., predation, competition, hybridization, transmission of parasites and diseases), but the presence of invaders can generate either increases or reductions in local biodiversity, since non-native organisms can either add species to local communities, or cause the extirpation of native species (McKinney, 2008; Magalhães et al., 2020a). All these alterations can influence beta diversity patterns (i.e., β-diversity), which is the degree of community differentiation across space, time, or any other dimension (Whittaker, 1960). This is, the interplay between immigration and extirpation of species structuring freshwater fish communities and beta diversity (Marchetti et al., 2001; Olden, Poff, 2003; Clavero, García-Berthou, 2006). For example, reductions in beta diversity indicate biotic homogenization (Olden et al., 2008), while increases in beta diversity indicate biotic differentiation (Larson, Pool, 2020). However, the role of the ornamental industry in inducing biotic homogenization and differentiation in aquatic communities has not been exhaustively described (e.g., Olden et al., 2008; Magalhães et al., 2020a), especially in the Neotropics. In this sense, Brazilian headwater creeks may be adequate systems to investigate how the introduction of non-native species shape the community structure (Magalhães, Jacobi, 2017; Magalhães et al., 2020a), as these systems are small-sized and highly invaded. Such studies may reveal how anthropogenic, environmental, and spatial factors (i.e., escapes from ornamental fish farms) affect beta diversity patterns.
To better understand how ornamental fish farms affect fish diversity patterns in contiguous environments, we investigated changes in fish beta diversity in headwater creeks located in the area affected by the largest ornamental aquaculture center in Brazil (Muriaé, Minas Gerais State). We investigated the structure of fish communities, the prevalence of NFAF species (i.e., translocated/exotic), and the changes in beta diversity patterns of local assemblages. We compared beta diversity patterns between historical and contemporary periods to determine trends indicating biotic homogenization or differentiation. Moreover, for the contemporary period, possible drivers (anthropogenic, environmental, and spatial) of beta diversity were investigated. We expected an increase in beta diversity over time (biotic differentiation) caused by the constant, but unpredictable, propagule pressure of different NFAF species, as ornamental fish farms promote multiple invasions over time. We also expected that spatial and environmental filters such as geographical location and temperature should explain fish occurrence and beta diversity across creeks.
Material and methods
Characterization of study area. This study was conducted in the largest ornamental aquaculture center in South America, which was established in the late 1970s, in Minas Gerais State, southeastern Brazil. The Muriaé Ornamental Aquaculture Center comprises 13 municipalities, and about 350 fish farms with 4,500 NFAF ponds (Magalhães, Jacobi, 2013, 2017). This center fulfills more than 70% of the Brazilian demand for aquarium fish (Magalhães, Jacobi, 2017). Most ponds are fully drained eight times a year without any protection against fish escapes (e.g., screens or detention ponds containing native predators such as trahira Hoplias malabaricus (Bloch, 1794) to eliminate non-natives before they have a chance to escape) (ALBM, pers. obs.). The water is usually released directly into headwater creeks, creating a huge pressure above 35,000 propagules each year (Magalhães et al., 2020a). In the past decade, the variety of NFAF cultivated in fish farms has increased from 60 to more than 200 species. Among the newly cultured species in the region are Endler’s guppy Poecilia wingei Poeser, Kempkes & Isbrücker, 2005, Panuco swordtail Xiphophorus nigrensis Rosen, 1960 (ALBM, pers. obs.), flagtail catfish Dianema urostriatum (Miranda Ribeiro, 1912), the oscar Astronotus ocellatus (Agassiz, 1831), the bronze corydoras Corydoras aeneus (Gill, 1858), and the transgenic Glofish® Danio rerio (Hamilton, 1822) (Magalhães et al., 2019, 2020a,b).
Six headwater tributaries (i.e., creeks) of the Glória River sub-basin (drainage area: 1,045 km2) were considered in the study, all belonging to the Paraíba do Sul River basin (Freshwater Ecoregions of the World #329, sensu Abell et al. (2008)). This is one of the most impacted watersheds in Brazil, covering an area of about 55,300 km2 distributed over São Paulo (24%: 13,272 km2), Minas Gerais (37%: 20,461 km2), and Rio de Janeiro (39%: 21,567 km2) States (Magalhães, Jacobi, 2013, 2017). The studied creeks run across the Atlantic Forest biome, one of the most important biodiversity hotspots in the world (Mittermeier et al., 2004). Riverine banks lack riparian vegetation, and are mostly covered by African grasses Urochloa spp. The native ichthyofauna is composed of a few native and endemic species, such as the two-spot minnow Deuterodon janeiroensis (Eigenmann, 1908), the catfish Trichomycterus spp., the dusky millions fish Phalloceros cf. leptokeras Lucinda, 2008, and the suckermouth catfish Neoplecostomus microps (Steindachner, 1877) (Magalhães et al., 2020a). It also includes rare species with limited distribution for the region, such as lambari Psalidodon parahybae (Eigenmann, 1908), piaba Astyanax cf. scabripinnis (Jenyns, 1842), catfish Trichomycterus auroguttatus Costa, 1992, suckermouth catfish Rineloricaria nigricauda (Regan, 1904), and the swamp eel Synbranchus marmoratus Bloch, 1795 (ALBM, pers. obs.).
Fish sampling. Fish individuals were collected (SISBIO license number 61740) with rectangular hand sieves (95 × 25 cm, and 0.3-mm mesh) along the margins and channel bed every two months (January to December). Sampling took place in different years (2017 and 2018) in each of the six headwater creeks: Lopes (LO: 20o50’16.5”S 42o13’48.9”W), Queiroga (QU: 20o56’10.1”S 42o18’34.4”W), and Boa Sorte (BS: 21o02’15.1”S 42o21’08.3”W) were sampled in 2017; Rochedo (RO: 21º07’19.7”S 42º18’38.1”W), Varginha (VA: 20o54’33.3”S 42o23’05.8”W), and São Luís (SL: 20o49’10.4”S 42o19’20.9”W) were sampled in 2018 (Fig. 1). In each creek, two people independently operated the sieve for 50 times along 100 m-long transects during a 2 h period, in the daylight. Each collector was positioned at opposing edges of the transect and moved from the edge toward the center. All fish (i.e., native and non-native) were anesthetized and sacrificed by immersion in Eugenol (Griffiths, 2000), fixed in 10% formalin, and transferred to 70% ethylic alcohol. These procedures were in accordance with the ‘Ethical Principles in Animal Research Guidelines’ adopted by the Brazilian College of Animal Experimentation (COBEA). The fluorescent strains of transgenic Glofish® D. rerio were identified using a 385 nm ultraviolet flashlight Sunshine model ss-003 (Guangzhou Sunshine Eletronics Technology Co. Ltd) in the dark. Voucher specimens were deposited under their respective catalogue numbers in the Universidade Federal de Sergipe fish collection, municipality of São Cristóvão, Sergipe State, Brazil (CIUFS 2210 to 2330; see Tab. S1). The species names were checked in Fricke et al. (2020).

FIGURE 1 | Sampling sites in the area affected by the Muriaé Ornamental Aquaculture Center in Brazil. Municipalities: Muriaé, Miradouro, Vieiras, and São Francisco do Glória (Total area of 1,419 km2; IBGE, 2020). Headwater creeks: LO = Lopes; QU = Queiroga; BS = Boa Sorte; RO = Rochedo; VA = Varginha; SL = São Luís.
Collected individuals were classified as native or non-native, the last sub-divided into translocated, i.e., species introduced from another river basin or freshwater ecoregion within Brazil, or exotic, i.e., species introduced from other countries (Alves et al., 2007; Vitule et al., 2019). Non-native fish species were identified according to Axelrod (1997) and Axelrod, Scott (2005). In addition, a list of native extirpated species (i.e., native species present in the creeks before the fish farms began operation) was developed based on the reports and informal interviews with experienced fish farmers (mean age: 72.27 ± 3.66 , N = 11: four in the municipality of Muriaé, four in the municipality of Vieiras, two in the municipality of Miradouro, one in the municipality of São Francisco do Glória); and anglers (mean age: 73.69 ± 4.92, N = 13: four in the municipality of Muriaé, two in the municipality of Vieiras, two in the municipality of Miradouro, five in the municipality of São Francisco do Glória) (sensu Magalhães et al., 2002). The information provided by fish farmers and anglers was later checked by the first author — who has studied fish from the Paraíba do Sul River basin in Minas Gerais since 2001 — and confirmed by colored photos, and specific literature (Menezes et al., 2007; Vieira, Rodrigues, 2010; Honório, Martins, 2018; Brito, Buckup, 2019).
Drivers of beta diversity. Environmental data were collected every two months in 2017 and also in 2018 (i.e., January-February, March-April, May-June, July-August, September-October, November-December), namely water temperature, water level (i.e., maximum creek or stream water elevation relative to a vertical gauge), pH, and rainfall. The first three were obtained before fish sampling at each site. Water temperature was measured with a mercury-in-glass thermometer with a scale marked from 0 to 50 ºC; water level (in cm) was measured using a 2 m-pole marked every 10 cm; and pH, with an MinyMix AL638 digital pH meter. Rainfall data (in mm) were obtained from the database of the weather station A517, located in the municipality of Muriaé (INMET, 2020).
The candidate spatial drivers of changes in beta diversity in the headwater creeks were latitude, longitude, and elevation. Coordinates were obtained using a Garmin GPS 60CX. Elevation was later referred to the coordinates in the online application Google Earth® (source: Image ©Maxar Technologies, www.maxar.com). The number of ponds used to raise fish in fish farms surrounding each sampling locality was the anthropogenic driver. We determined the number of ponds through simple counting after visiting each fish farm. The information about the number of individuals released in each drainage (propagule size) was not available, so we restricted our analysis to the number of ponds in each locality: Lopes Creek (LO) = one fish farm, 120 ponds, Queiroga Creek (QU) = one fish farm, 76 ponds, Boa Sorte Creek (BS) = one fish farm, 94 ponds, Rochedo Creek (RO) = one fish farm, 132 ponds, Varginha Creek (VA) = one fish farm, 16 ponds, and São Luís Creek (SL) = one fish farm, 110 ponds.
Data analysis. Species richness was expressed as the number of native or non-native (translocated and exotic) species per sampled creek. Species occurrence (%) across creeks was calculated as:
Li/Lt×100;
where Li is the total number of creeks where the species i was found, and Lt is the total number of creeks.
The proportion of non-native species in each creek (i.e., level of contamination by non-native species) was obtained using the Contamination Index (CI) with minor modification (Alves et al., 2007), calculated as the proportion of non-native species:
CI = NN/(N+NN);
where NN is the number of non-native species (translocated and exotic), and N is the number of native species. The CI varied from 0 (purely native assemblages) to 1, in assemblages totally contaminated by non-native species.
We quantified the changes in beta diversity over time, from historical to the contemporary state. A historical dataset was constructed consisting of native extirpated and native extant species (i.e., the list of native extirpated species mentioned in the “fish sampling” section plus the native species recorded in all creeks over the entire sampling period). The contemporary dataset corresponded to native and non-native species (translocated and exotic) sampled in all creeks in 2017 and 2018. In each period (historical and contemporary), with creeks as sampling units, we measured beta diversity as the average distance to the centroid of presence-absence dissimilarities in the multivariate space (Anderson et al., 2006). A decrease in beta diversity over time indicates biotic homogenization, while an increase in beta diversity over time indicates biotic differentiation (e.g., Olden et al., 2008; Larson, Pool, 2020). PBeta diversity was compared between periods with a Wilcoxon test for paired samples. Data handling and statistical analyses were performed in R environment (R Development Core Team, 2020), using “tidyverse” (Wickham et al., 2019) and ‘‘vegan’’ (Oksanen et al., 2013) packages, respectively.
Model selection. We employed a modeling approach to investigate possible drivers of beta diversity in the contemporary period, for which a normal distribution of residuals (Legendre, De Cáceres, 2013) and symmetric dissimilarities (Anderson et al., 2006) are required assumptions. Therefore, transformations (“decostand” function) were performed in the community composition matrix. Our response variable matrix D was composed of Euclidean distances of log (x + 1) chord transformations of fish species presence-absence (columns) in each campaign (36 rows, 6 months × 6 fish farms) (Legendre, Borcard, 2018). Despite being symmetric, the resulting dissimilarity matrix D approached double-zero asymmetric Jaccard dissimilarities of binary data, as suggested for beta diversity studies (Legendre, De Cáceres, 2013). Jaccard dissimilarities (Dj) used previously to compare historical and contemporary periods were also employed as the response variable for comparison with D.
We partitioned the variation (“varpart” function) of D and Dj according to four matrices of covariates that were environmental (water temperature, water level, pH, and rainfall), spatial (latitude, longitude, and elevation), and anthropogenic (number of ponds), quantified in each campaign at the Muriaé Ornamental Aquaculture Center. The number of ponds and the geographical position of each fish farm were constant during the whole sampling period. We did not search for mantel correlations between the matrices of independent covariates because the variation partitioning procedure was robust against auto correlated co variation, which was also balanced by the calculation of the adjusted R2 (adj-R2, Peres-Neto et al.,2006).
To investigate the individual influence of individual covariates, we employed a selection procedure based on the calculation of the adj-R2 after permutations on response matrices D or Dj, in a Permutational Multivariate Analysis of Variance (PERMANOVA, “adonis” function; 999 permutations). Matrices D and Dj were explained by all possible interactions between uncorrelated independent covariates.
The use of autocorrelated variables was prevented with previous correlation tests (Spearman’s ρ) between pairs of explanatory covariates. Considering spatial variables, we removed elevation and longitude, which were correlated with latitude (Lat, ρ = 0.94; P < 0.01) and the number of ponds (Np, ρ = 0.67; P < 0.01), respectively. Among environmental variables, we removed rainfall and water level, which were correlated among themselves (ρ = 0.50; P < 0.01), with temperature (Temp, ρ = 0.64; P < 0.01), and with pH (ρ = 0.62; P < 0.01). These last two variables were also correlated (ρ = 0.47; P < 0.01). Therefore, only temperature was used as an environmental proxy in the model. Then, the final structure of the PERMANOVA was:
D(j) = Temp×Lat×Np;
in which the best predictors, or their combinations were selected according to the highest adj-R2.
Results
Our study recorded 60 fish species, of which 16 (26.66%) were native and 44 non-natives, of which 19 were translocated (31.67%) and 25 exotic (41.67%) (see Tab. S1). The native species richness in local assemblages varied from three to nine: São Luís and Varginha creeks were the richest (Fig. 2). Non-native translocated species ranged from one to ten, with the Boa Sorte Creek presenting the highest number. On the other hand, the richness of non-native exotic species was high (above native species richness) in all creeks, except for the Varginha Creek. The water temperature varied between 17.8oC in Varginha Creek to 27.2oC in Boa Sorte Creek. Water level fluctuated between 29.5 cm in Boa Sorte Creek to 90 cm in Varginha Creek. The pH remained acidic in Lopes (6.5), Queiroga (6.7), Boa Sorte (6.4), Rochedo (6.6), and Varginha (6.9) creeks, but was neutral in São Luís Creek. Rainfall varied between 105.7 mm in Varginha Creek, municipality of Miradouro, to 119.2 mm in Boa Sorte, and Rochedo creeks, municipality of Muriaé (see Tab. S2 and Fig. S3).
Among the 44 non-native fish species, the most widely distributed in the studied headwater creeks in terms of species occurrence (%) were: the guppy Poecilia reticulata Peters, 1859 (100%), southern platyfish Xiphophorus maculatus (Günther, 1866) (83.33%), variable platyfish X. variatus (Meek, 1904) (66.67%), D. rerio (66.67%), and the oriental weatherloach Misgurnus anguillicaudatus (Cantor, 1842) (66.67%) (Fig. 3).

FIGURE 2 | Richness of native (blue) and non-native species [translocated (yellow) and exotic (red)], in each headwater creek in the Muriaé Ornamental Aquaculture Center, Brazil. Headwater creeks: LO = Lopes; QU = Queiroga; BS = Boa Sorte; RO = Rochedo; VA = Varginha; SL = São Luís.

FIGURE 3 | The 10 most widespread non-native exotic fish in the studied headwater creeks located in the Muriaé Ornamental Aquaculture Center, Brazil. Only non-native species with at least 50% of occurrence were listed.
The CI considering non-native translocated species ranged from 0.20 to 0.63, with the Boa Sorte and Queiroga creeks presenting respectively the highest and lowest contaminations. The CI considering non-native exotic species ranged from 0.13 to 0.81, with the Rochedo and Lopes creeks presenting the highest values, and Varginha Creek the lowest (Fig. 4). Most creeks’ CI values were higher than 0.6, indicating that creeks were dominated by non-native exotic species.
The contemporary period was dominated by non-native species and presented greater distances to the centroid (i.e., beta diversity) in relation to the historical period (Wilcoxon test, N = 6, P = 0.03, Fig. 5). The Boa Sorte Creek presented the greatest difference (0.3) between periods, followed by Queiroga (0.23) and Varginha (0.23), Lopes (0.12), Rochedo (0.11), and São Luís (0.11). Beta diversity values increased from historical to contemporary periods in all creeks due to the extirpation of native species and the introduction of NFAF species, indicating both turnover and biotic differentiation. In relation to the possible pristine environment (historical period), creeks in the contemporary period experienced the introduction and differential colonization pressure of several NFAF species, influencing beta diversity mainly by the replacement of pristine (i.e., native) communities (Tab. S1).

FIGURE 4 | Contamination Index (CI) in each headwater creek in the Muriaé Ornamental Aquaculture Center, Brazil. Headwater creeks: LO = Lopes; QU = Queiroga; BS = Boa Sorte; RO = Rochedo; VA = Varginha; SL = São Luís.

FIGURE 5 | Distances to the centroid obtained from the two main Principal Coordinate Analysis – PCoA axis (see Anderson et al., 2006 for further details) of fish community Jaccard dissimilarities in six headwater creeks (LO = Lopes; QU = Queiroga; BS = Boa Sorte; RO = Rochedo; VA = Varginha; SL = São Luís) sampled in historical (only native species) and in contemporary (native + non-natives) periods, in the Muriaé Ornamental Aquaculture Center, Brazil.
The temporal and spatial differences in fish composition represented by D in the contemporary periodwere influenced mainly by the interaction between environmental and spatial variables (adj-R2 = 0.53; P = 0.001), and by the interaction between spatial variables and number of ponds (adj-R2 = 0.52; P = 0.001), with 64% of the variation explained by all explanatory covariates (Tab. 1; Fig. S3; VARPART). The unexplained variation was smaller when using D (adj-R2 = 0.36) than Dj (adj-R2= 0.43) as the response matrices. Therefore, the following results and discussion were restricted to D. When the community matrix was submitted to permutations in the contemporary period (Tab. 1; PERMANOVA), the fish composition was individually explained particularly by number of ponds (Np, adj-R2= 0.24; Pseudo-F = 18.65; P = 0.001), but also by Np associated with latitude (adj-R2= 0.04; Pseudo-F = 6.86; P = 0.001), and with temperature (adj-R2 = 0.01; Pseudo-F = 4.47; P = 0.001). In addition, the geographical position, as well as temperature, explained individually the D composition in 4% of the runs (adj-R2 = 0.04; Fig. 6).
TABLE 1 | Variation partitioning (VARPART) of the response matrix (D, fish composition, log-transformed chord Euclidean distances), and Permutational Multivariate Analysis of Variance (PERMANOVA) of D, according to anthropogenic (Np = number of ponds), environmental (Env), and spatial (Sp) variables measured in the Muriaé Ornamental Aquaculture Center, Brazil. Environmental variables include water temperature (Temp, °C), water level (cm), pH, and rainfall (mm). In the PERMANOVA, only uncorrelated variables were included in the model. Variables are ranked according to the adjusted R2 (Adj-R2) values. Degrees of freedom (Df); Sum of Squares (SS); F-values (VARPART), and Pseudo-F ratios (PERMANOVA) are shown.
| VARPART | Df | SS | F-value | R2 | Adj-R2 | P |
| All | 8 | 12.54 | 8.66 | 0.72 | 0.64 | 0.001 |
| Env + Sp | 7 | 10.93 | 6.73 | 0.63 | 0.53 | 0.001 |
| Sp + Np | 4 | 9.97 | 10.36 | 0.57 | 0.52 | 0.001 |
| Env + Np | 5 | 9.38 | 7 | 0.54 | 0.46 | 0.001 |
| Sp | 3 | 7.51 | 8.08 | 0.43 | 0.38 | 0.001 |
| Env | 4 | 7.5 | 5.85 | 0.43 | 0.36 | 0.001 |
| Np | 1 | 5.08 | 13.98 | 0.29 | 0.27 | 0.001 |
| PERMANOVA | Pseudo-F | |||||
| Np | 1 | 2.65 | 18.65 | 0.26 | 0.24 | 0.001 |
| Temp | 1 | 0.72 | 5.09 | 0.07 | 0.04 | 0.001 |
| Lat | 1 | 0.70 | 4.91 | 0.07 | 0.04 | 0.001 |
| Lat:Np | 1 | 0.97 | 6.86 | 0.10 | 0.04 | 0.001 |
| Np:Temp | 1 | 0.64 | 4.47 | 0.06 | 0.01 | 0.001 |
| Lat:Temp | 1 | 0.24 | 1.67 | 0.02 | -0.04 | 0.09 |
| Lat:Np:Temp | 1 | 0.26 | 1.84 | 0.03 | -0.07 | 0.067 |
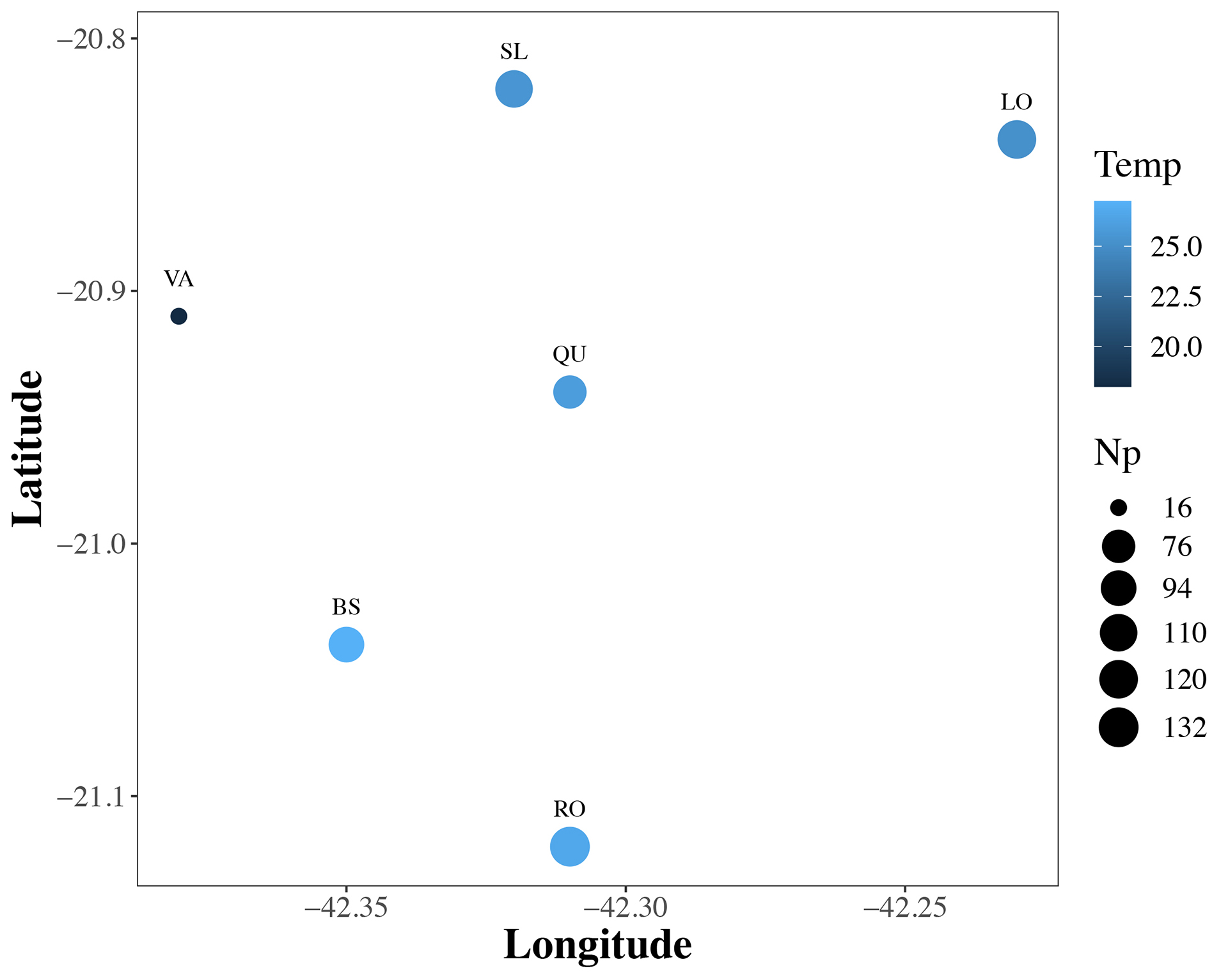
FIGURE 6 | Position of six headwater creeks (LO = Lopes; QU = Queiroga; BS = Boa Sorte; RO = Rochedo; VA = Varginha; SL = São Luís), scaled by temperature (Temp, blue gradient colours), and number of ponds (Np, circle size) used to raise fish species in the nearest fish farm (i.e., anthropogenic proxy of propagule pressure) in the Muriaé Ornamental Aquaculture Center, Brazil.
Discussion
We recorded significant changes in fish species composition of headwater creeks invaded by non-native species in the surroundings of the Muriaé Ornamental Aquaculture Center, Brazil. A massive fish invasion is underway and has induced patterns of biotic differentiation among fish assemblages (i.e., higher beta diversity), changing the original community structure (i.e., historical period). During the contemporary period, anthropogenic, environmental, and spatial filters limited the occurrence and turnover of some NFAF species, but the biotic differentiation was explained particularly by the number of ponds registered in each fish farm.
Non-native species have progressively invaded fish assemblages in the study area, a process occurring at the local level. A previous study in the region demonstrated that the environmental degradation and loss of regional biodiversity were associated with the predominance of NFAF species (Magalhães et al., 2020a). Here, we added that biotic differentiation among creeks was a consequence of the contamination of natural assemblages by a massive number of non-native species, co-occurring with a general decrease in abundance (both native and non-native individuals), during contemporary periods (Magalhães et al., 2020a), native species extinctions, and non-native turnover, therefore representing community heterogenization in the long-term. In the contemporary period, the conditions in the Varginha Creek, with lower water temperatures (< 20oC) and smaller propagule pressure (Np = 16), must have prevented the establishment of some non-native species, resulting in less contamination and variation in the local fish composition. However, these conditions were an exception in the studied area, as NFAF were predominant in other creeks. The other headwater creeks were located at intermediate elevations (454 m a.s.l. on average), with water temperatures above 20oC, exceeding 30oC in spring/summer. These creeks were impacted by human activities: the channels were silted and the banks lacked riparian vegetation in many stretches, likely creating favorable conditions for the establishment of NFAF species. Moreover, fish farms in these creeks held many production ponds (> 70, the main individual proxy explaining the fish community composition) and raised different NFAF species, creating a constant propagule pressure at a regional level that could be related to the distribution of fish species across creeks (Magalhães, Jacobi, 2008, 2013, 2017; Magalhães et al., 2002, 2019, 2020a).
In addition to the predominance of non-native species, the proportion of native species has decreased over time. We did not test cause-effect relationships on contrasting dynamics between native and non-native communities, but 23 new NFAF species (14 translocated and nine exotics) invaded the study area in the last few years, a high value when compared to non-native species reported in other creeks of the region (Magalhães et al., 2020a). In the last decade, Brazil has recorded numerous fish introductions, particularly NFAF, both translocated and exotic (Rocha et al., 2011; Vitule et al., 2012; Magalhães, Jacobi, 2013, 2017; Frehse et al., 2016; Magalhães et al., 2017, 2019, 2020a; Bezerra et al., 2019; Vitule et al., 2019). Our study confirms this trend and shows how ornamental aquaculture can affect fish assemblages in natural ecosystems, as fish escape from aquaculture ponds and invade contiguous areas.
We emphasize the high levels of contamination by non-native species (translocated and exotic) in each headwater creek, especially exotic species. The number of introductions peaked in the past fifteen years across the Minas Gerais State, when compared to the rest of Brazil (Melo et al., 2006; Alves et al., 2007; Vieira, Rodrigues, 2010; Magalhães, Jacobi, 2013, 2017; Magalhães et al., 2019). Many new records of non-native species have been documented in some watersheds, especially in the Paraíba do Sul River basin, where the Muriaé Ornamental Aquaculture Center is located. This ornamental fish-culture industry is responsible for producing about 10 million NFAF per year, resulting in a constant propagule pressure and high levels of contamination in local fish assemblages (Alves et al., 2007; Magalhães, Jacobi, 2013, 2017; Magalhães et al., 2019, 2020a). Our results revealed the predominance of non-native exotic species originating from countries of Central America, South America, Europe, Africa, and Asia in the contemporary period, which reached the creeks due to the draining of the NFAF ponds, with the water being discharged directly into headwater creeks (sensu Magalhães, Jacobi, 2008, 2013, 2017; Magalhães et al., 2019, 2020a). The Boa Sorte Creek was likely the most susceptible to the turnover of non-native exotic species, given the elevated distances to the centroid in the contemporary period (behind only of the Varginha Creek), and their increase in relation to the historical period.
Fish farmers are constantly searching for new non-native exotic species as market options, such as the orange chromide Pseudetroplus maculatus (Bloch, 1795), moonlight gourami Trichopodus microlepis (Günther, 1861), and the transgenic zebrafish D. rerio (Mendonça, Thomé, 2019; Magalhães et al., 2020b). Pristine environments (i.e., free from alterations or less disturbed) are becoming rare in the Paraíba do Sul River basin, as the watershed has been extensively degraded by multiple human impacts (e.g., impoundments, deforestation, siltation, non-native species). Currently, most lakes, reservoirs (e.g., Glória, Ormelo Junqueira Botelho, and Ilha dos Pombos reservoirs), streams/creeks (e.g., Boa Vista, Pinheiros, Santo Antônio, Quilombo creeks), and rivers (e.g., Una, Carangola, Glória, Muriaé, and Paraíba do Sul rivers) are invaded and disturbed by non-native fishes, which, in large spatial and temporal scales, are leading the ichthyofauna and their ecosystems to simplification (Araújo et al., 2001; Melo et al., 2006; Alves et al., 2007; Vieira, Rodrigues, 2010; Moraes et al., 2017; Honório, Martins, 2018; Brito, Buckup, 2019; Thomé et al., 2019; Magalhães et al., 2019, 2020a).
We recorded the introduction of numerous NFAF species, which probably played a role in the replacement of original assemblages. This strong shift in species composition, with the dominance of non-native fishes, may destabilize native communities and make them more susceptible to stochastic disturbances (Erös et al., 2020). Among the non-native exotic species recorded, the guppy P. reticulata, southern platyfish X. maculatus, variable platyfish X. variatus, zebrafish D. rerio (non-transgenic), and the oriental weatherloach M. anguillicaudatus were more widely distributed in the study area. Araújo et al. (2001), Melo et al. (2006), Magalhães et al. (2013, 2017), Moraes et al. (2017), and Thomé et al. (2019) reported that P. reticulata, X. maculatus, X. variatus, D. rerio (non-transgenic), and M. anguillicaudatus were introduced in various locations in the Paraíba do Sul River basin. In some of these locations, there are reports of ecological impacts (e.g., change in native fish community structure, biotic homogenization/differentiation) caused by these species (Araújo et al., 2001; Thomé et al., 2019; Magalhães et al., 2020a).
The Muriaé Ornamental Aquaculture Center has been a major source of NFAF species in different water bodies in the Paraíba do Sul River basin. This is probably one of the most invaded aquatic systems in the world, bringing together fishes from different continents and ecoregions. We demonstrate that the massive and continuous introduction events changed beta diversity patterns by inducing biotic differentiation among creek. As some of the NFAF reported here were also responsible for inducing biotic differentiation patterns in other creeks of the study area, such as M. anguillicaudatus, rosy barb Pethia conchonius (Hamilton, 1822), and the black molly P. sphenops Valenciennes, 1846 (Magalhães et al., 2020a), biotic differentiation at the local scale can be inexorable. Such an extreme case of fish invasion threatens native populations, which may experience strong decline, extirpation (i.e., local extinction) and range reduction, such as piaba A. cf. scabripinnis, suckermouth catfish N. microps, and catfish Trichomycterus spp., which can be considered rare or with limited distribution (Magalhães, Jacobi, 2008, 2013; Magalhães et al., 2019, 2020a).
In addition to changing composition patterns, the invasion of NFAF revealed possible negative ecological impacts and new introduction pathways. We observed hybridization between the native guppy Poecilia vivipara Bloch & Schneider, 1801 and the exotic X. variatus, and the infection of the native dogfish Oligosarcus hepsetus (Cuvier, 1829), and the cascarudo Callichthys callichthys (Linnaeus, 1758) by the exotic parasite anchor worm Lernaea cyprinacea (Linnaeus, 1758). Indication of hybridization between native and exotic poeciliids, and contamination by anchor worm in other native fish species (e.g., two-spot astyanax Astyanax bimaculatus (Linnaeus, 1758) = two-spot minnow D. janeiroensis) have been reported in the area since 2005 (Magalhães, 2006; Magalhães, Jacobi, 2017). Moreover, the “artificial” and aggressive transgenic D. rerio, detected for the first time in Latin America and probably in the Southern Hemisphere (ALBM, pers. obs.), can strongly compete for resources with native fishes (Cowx et al., 2010). The presence of the piaba Knodus moenkhausii (Eigenmann & Kennedy, 1903) points to a new introduction pathway: the live-food fish trade. This fish is farmed exclusively to feed large predatory fishes, such as the oscar A. ocellatus, and may constitute a new additional route for fish introductions in areas affected by ornamental aquaculture (Magalhães et al., 2019).
Our results confirm that escapes from aquaculture facilities are common, and could bring severe consequences to local fish populations including endemic, rare, and threatened species (Lima Junior et al., 2018; Magalhães et al., 2020a). Such scenario requires management strategies to reduce introduction rates and impacts, while fostering sustainable aquaculture and protecting native biodiversity. In this sense, a sustainable development plan should be guided by three critical principles (i.e., triple bottom line sensu Nobile et al., 2020): environmental protection, social development, and economic viability. First, aquaculture should value local fish diversity. In the present case, this could include farming several native fish from the Paraíba do Sul River basin with ornamental potential, such as the yellow tetra Hyphessobrycon bifasciatus Ellis, 1911, banded knifefish Gymnotus cf. carapo Linnaeus, 1758, sailfin catfish Hypostomus affinis (Steindachner, 1877), and the pearl cichlid Geophagus brasiliensis (Quoy & Gaimard, 1824) (Menezes, 2007; Vieira, Rodrigues, 2010). Non-natives should be avoided (e.g., Magalhães, Jacobi, 2013, 2017), and transgenic fish should be banned (e.g., Magalhães et al., 2020b). Second, social development must be fostered through education and training of people who work in ornamental fish farms (i.e., workers and managers). The implementation of a Best Management Practices (BMPs) program is an excellent way to improve management options, since fish escapes are associated mainly with operational failures (e.g., Magalhães, Jacobi, 2017; Nobile et al., 2020). Such a program could improve management practices during pond draining and harvest to avoid the release of fish species at random, or by establishing actions to recapture fish in the environment, considering that ornamental fish remain in the immediate vicinity soon after the introduction (Tuckett et al., 2017; Magalhães et al., 2019).Finally, economic aspects must be considered, since NFAF lost to the environment represent economic losses to the farmer. In this study, a total of 10,510 individuals of 44 non-native species escaped from ornamental fish farms between 2017 and 2018, totaling a financial loss of US$ 4,386 (US$ 2,193 per year). This value could be reversed in the purchase of commercial feed, medicines, and devices to prevent escapes (e.g., anti-escape nylon screens or construction of detention ponds) or by hiring specialized personnel such as Biologists, Veterinarians, Zootechnicians or Aquaculture Engineers. Thus, the cessation of propagule and colonization pressures could positively affect the environment and the regional economy (Magalhães et al., 2019, 2020a). In addition to the management strategies suggested above, we emphasize that, from a scientific and environmental point of view, this would be a great area for a controlled management of creeks with before-after-control-impact (BACI) designs (e.g., Methratta, 2020). We emphasize that these ecosystems could be an ideal model for the first work on the eradication and control of non-native fish in South America, which would be a historical landmark.
Our study reported significant changes in fish diversity in headwater creeks invaded by multiple ornamental fish, in the area affected by the Muriaé Ornamental Aquaculture Center. This is one of the most impressive case of fish invasion in the world, where non-native species have substantially replaced native and other non-native species. It is evident that anthropogenic, environmental, and spatial filters have played a role in the spread of NFAF, but a constant and high propagule pressure was the main individual factor associated with the high invasion rates and fish community assembling. Moreover, the invaded assemblages may be more susceptible to new invasions, since non-native populations can facilitate secondary introductions and colonization (Xiong et al., 2019). In fact, we confirmed that NFAF species could replace other NFAF locally. Future studies should devote attention to turnover or facilitation dynamics, which may be common in the studied area.
Acknowledgments
We thank the ornamental fish farmers who granted access to their properties, especially the farmer Jorge Farias for field assistance and contact with ancient fish farmers and anglers. Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio) provided collecting license. Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) provided research productivity grants provided for FMP (PQ Process Number 306869/2017–9), JRSV (PQ Process Numbers: 302367/2018–7 and 303776/2015–3) and MFGB (PQ Process Number 306851/2018–0). We also thank two anonymous reviewers for their valuable comments.
References
Abell R, Thieme ML, Revenga C, Bryer M, Kottelat M, Bogutskaya N et al. Freshwater ecoregions of the world: a new map of biogeographic units for freshwater biodiversity conservation. BioScience. 2008; 58(5):403–14. https://doi.org/10.1641/B580507
Alves CBM, Vieira F, Magalhães ALB, Brito MFG. Impacts of nonnative fish species in Minas Gerais, Brazil: present situation and prospects. In: Bert TM, editor. Ecological and genetic implications of aquaculture activities. Dordrecht: Springer Press; 2007. p.291–314.
Anderson MJ, Ellingsen KE, McArdle BH. Multivariate dispersion as a measure of beta diversity. Ecol Lett. 2006; 9(6):683–93. https://doi.org/10.1111/j.1461-0248.2006.00926.x
Araújo FG, Fichberg IP, Benjamin CT, Peixoto MG. Variações espaciais na assembléia de peixes no Rio Paraíba do Sul (Barra Mansa, Barra do Piraí), Rio de Janeiro, Brasil. Rev Bras Zool. 2001; 18(2):483–92. https://doi.org/10.1590/S0101-81752001000200019
Axelrod HR. Dr. Axelrod’s atlas of freshwater aquarium fishes.Neptune City: TFH Publications; 1997.
Axelrod GS, Scott BM. Encyclopedia of Exotic Tropical Fishes for Freshwater Aquariums. Neptune City: TFH Publications; 2005.
Bezerra LAV, Matheus OF, Daga VS, Occhi TVT, Faria L, Costa APL et al. A network meta-analysis of threats to South American fish biodiversity. Fish Fish. 2019; 20(4):620–39. https://doi.org/10.1111/faf.12365
Brito V, Buckup PA. The fish fauna of the upper Piraí drainage, a transposed mountain river system in southeastern, Brazil. Check List. 2019; 15(1):235–47. https://doi.org/10.15560/15.1.235
Brito MFG, Daga VS, Vitule JRS. Fisheries and biotic homogenization of freshwater fish in the Brazilian semiarid region. Hydrobiologia. 2020; 847:3877–95. https://doi.org/10.1007/s10750-020-04236-8
Bush ER, Baker SE, MacDonald DW. Global trade in exotic pets 2006–2012. Conserv Biol. 2014; 28(3):663–76. https://doi.org/10.1111/cobi.12240
Clavero M, García-Berthou E. Homogenization dynamics and introduction routes of invasive freshwater fish in the Iberian Peninsula. Ecol Appl. 2006; 16(6):2313–24. https://doi.org/10.1890/1051-0761(2006)016[2313:HDAIRO]2.0.CO;2
Cowx IG, Bolland JD, Nunn AD, Kerins G, Stein J, Blackburn J et al. Defining environmental risk assessment criteria for genetically modified fishes to be placed on the EU market. EFSA. 2010; 7(11):69E. https://doi.org/10.2903/sp.efsa.2010.EN-69
Erös T, Comte L, Filipe AF, Ruhi A, Tedesco PA, Brose U et al. Effects of nonnative species on the stability of riverine fish communities. Ecography.2020; 43(8):1156–66. https://doi.org/10.1111/ecog.04985
Food Agriculture Organization (FAO). The state of world fisheries and aquaculture [Internet]. Rome: Editorial Group FAO Information Division; 2000. Available from: http://www.fao.org/DOCREP/003/X8002E/X8002E00.htm
Frehse FA, Braga RR, Nocera GA, Vitule JRS. Non-native species and invasion biology in a megadiverse country: scientometric analysis and ecological interactions in Brazil. Biol Invasions. 2016; 18:3713–25. https://doi.org/10.1007/s10530-016-1260-9
Fricke R, Eschmeyer WN, Van der Laan R. Eschmeyer’s catalog of fishes: genera, species, references [Internet]. San Francisco: California Academy of Science; 2020. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Griffiths SP. The use of clove oil as an anaesthetic and method for sampling intertidal rockpool fishes. J Fish Biol. 2000; 57(6):1453–64. https://doi.org/10.1111/j.1095-8649.2000.tb02224.x
Honório JR, Martins IA. Ichthyofauna of the Una River in the Paraíba do Sul Paulista River Valley, Southeastern of Brazil. Biota Neotrop. 2018; 18(4):e20180528. https://doi.org/10.1590/10.1590/1676-0611-bn-2018-0528
Hulme PE. Invasion pathways at a crossroad: policy and research challenges for managing alien species introductions. J Appl Ecol. 2015; 52(6):1418–24. https://doi.org/10.1111/1365-2664.12470
Instituto Brasileiro de Geografia e Estatística (IBGE). Área territorial oficial [Inernet]. Instituto Brasileiro de Geografia e Estatística; 2020. Available from: https://www.ibge.gov.br
Instituto Nacional de Metereologia (INMET). Dados históricos [Internet]. Instituto Nacional de Metereologia; 2020. Available from: http://www.inmet.gov.br/portal/index.php?r=bdmep/bdmep
Ishikawa T, Tachihara K. Introduction history of nonnative freshwater fish in Okinawa-jima Island: ornamental aquarium fish pose the greatest risk for future invasions. Ichthyol Res. 2014; 61:17–26. https://doi.org/10.1007/s10228-013-0367-6
Islami MF, Sudrajat AO, Carman O. Induction of maturation and ovulation of Red Fin Shark fish Epalzeorhynchos frenatus in non-spawning season. Int J Fish Aquat Stud. 2017; 5(4):418–24.
Larson ER, Pool TK. Biological invasions drive biotic homogenization of North American crayfishes. Hydrobiologia. 2020; 847:3795–809. https://doi.org/10.1007/s10750-019-04164-2
Legendre P, Borcard D. Box-Cox-chord transformations for community composition data prior to beta diversity analysis. Ecography. 2018; 41(11):1820–24. https://doi.org/10.1111/ecog.03498
Legendre P, De Cáceres M. Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecol Lett. 2013; 16(8):951–63. https://doi.org/10.1111/ele.12141
Lima Junior DP, Magalhães ALB, Pelicice FM, Vitule JRS, Azevedo-Santos VM, Orsi ML et al. Aquaculture expansion in Brazilian freshwaters against the Aichi Biodiversity Targets. Ambio. 2018; 47(4):427–40. https://doi.org/10.1007/s13280-017-1001-z
Lockwood JL, Welbourne DJ, Romagosa CM, Cassey P, Mandrak NE, Strecker A et al. When pets become pests: the role of the exotic pet trade in producing invasive vertebrate animals. Front Ecol Environ. 2019; 17(6):323–30. https://doi.org/10.1002/fee.2059
Magalhães ALB. First record of lernaeosis in a native fish species from a natural environmental in Minas Gerais state, Brazil. Panam J Aquat Sci. 2006; 1(1):8–10.
Magalhães ALB, Amaral IB, Ratton TF, Brito MFG. Ornamental exotic fishes in the Glória reservoir and Boa Vista Stream, Paraíba do Sul river basin, state of Minas Gerais, southeastern Brazil. Comun Mus Ciênc Tecnol Sér Zool. 2002; 15(2):265–78.
Magalhães ALB, Brito MFG, Sarrouh B. An inconvenient routine: introduction, establishment and spread of new non-native fishes in the Paraíba do Sul River basin, state of Minas Gerais, Brazil. Neotropical Biol Conserv. 2019; 14(3):329–38. https://doi.org/10.3897/neotropical.14.e38058
Magalhães ALB, Daga VS, Bezerra LAV, Vitule JRS, Jacobi CM, Silva LGM. All the colors of the world: biotic homogenization-differentiation dynamics of freshwater fish communities on demand of the Brazilian aquarium trade. Hydrobiologia. 2020a; 847:3897–915. https://doi.org/10.1007/s10750-020-04307-w
Magalhães ALB, Lima-Junior DP, Brito MFG, Vitule JRS, Azevedo-Santos VM, Pelicice FM et al. Peixe ilegal ainda à venda no Brasil: o exemplo do “carismático” não-nativo paulistinha transgênico (Danio rerio) e os riscos de sua provável introdução. Bol Soc Bras Ictiol. 2020b; 132:39–52. Available from: https://www.sbi.bio.br/images/sbi/boletim-docs/2020/junho_132.pdf
Magalhães ALB, Jacobi CM. Ornamental exotic fish introduced into Atlantic Forest water bodies, Brazil. Neotropical Biol Conserv. 2008; 3(2):73–77. Available from: http://revistas.unisinos.br/index.php/neotropical/article/view/5448/2684
Magalhães ALB, Jacobi CM. Asian aquarium fishes in a Neotropical biodiversity hotspot: impeding establishment, spread and impacts. Biol Invasions. 2013, 15:2157–63. https://doi.org/10.1007/s10530-013-0443-x
Magalhães ALB, Jacobi CM. Colorful invasion in permissive Neotropical ecosystems: Establishment of ornamental non-native poeciliids of the genera Poecilia/Xiphophorus (Cyprinodontiformes: Poeciliidae) and management alternatives. Neotrop Ichthyol. 2017; 15(1):e160094. https://doi.org/10.1590/1982-0224-20160094
Magalhães ALB, Orsi ML, Pelicice FM, Azevedo-Santos VM, Vitule JRS, Lima-Junior DP et al. Small size today, aquarium dumping tomorrow: sales of juvenile non-native large fish as an important threat in Brazil. Neotrop Ichthyol. 2017; 15(4):e170033. https://doi.org/10.1590/1982-0224-20170033
Marchetti MP, Light T, Feliciano J, Armstrong T, Hogan Z, Viers J et al. Homogenization of California’s fish fauna through abiotic change. In: Lockwood JL, McKinney ML, editors. Biotic homogenization. New York: Kluwer Academic/Plenum Publishers; 2001. p.259–78.
McKinney ML. Do humans homogenize or differentiate biotas? It depends. J Biogeogr. 2008; 35(11):1960–61. https://doi.org/10.1111/j.1365-2699.2008.02011.x
Melo FCA, Machado AC, Oliveira APC, Cruz JM, Latini AO. Ictiofauna do rio Carangola, bacia do rio Paraíba do Sul, no município de Carangola, Minas Gerais, Brasil. Lundiana. 2006; 7(2):133–44.
Mendonça NP, Thomé MPM. Caracterização dos piscicultores ornamentais da Zona da Mata mineira. Div Gestão. 2019; 3(1):47–62. Available from: https://itr.ufrrj.br/diversidadeegestao/wp-content/uploads/2019/09/DG035_Mendonca_Thome.pdf
Mendoza R, Luna S, Aguilera C. Risk assessment of the ornamental fish trade in Mexico: analysis of freshwater species and effectiveness of the FISK (Fish Invasiveness Screening Kit). Biol Invasions. 2015; 17:3491–502. https://doi.org/10.1007/s10530-015-0973-5
Menezes NA, Weitzman SH, Oyakawa OT, Lima FCT, Castro RMC, Weitzman MJ. Freshwater fishes of Mata Atlântica: Preliminary list of species and comments on conservation of Neotropical freshwater fishes. São Paulo: Universidade de São Paulo; 2007.
Methratta ET. Monitoring fisheries resources at offshore wind farms. BACI vs. BAG designs. ICES J Mar Sci. 2020; 77(3):890–900. https://doi.org/10.1093/icesjms/fsaa026
Meyers M. The pet industry view. In: Chao NL, Petry P, Prang G, Sonnesachien L, Teusty M, editors. Conservation and management of ornamental fish resources of the Rio Negro Basin, Amazônia, Brasil – Project Piaba. Manaus: Editora da Universidade do Amazonas; 2001. p.87–108.
Mittermeier RA, Robles Gil P, Hoffmann M, Pilgrim J, Brooks T, Mittermeier CG et al. Hotspots revisited: Earth´s biologically richest and most endangered terrestrial ecoregions. México City: CEMEX/Agrupación Sierra Madre; 2004.
Moraes MB, Polaz CNM, Caramaschi EP, Santos Júnior SS, Souza G, Carvalho FL. Espécies exóticas e alóctones da bacia do rio Paraíba do Sul: implicações para a conservação. Biodiv Bras. 2017; 7(1):34–54. https://doi.org/10.37002/biobrasil.v%25vi%25i.563
Nobile AB, Cunico AM, Vitule JRS, Queiroz J, Vidotto-Magnoni AP, Garcia DAZ et al. Status and recommendations for sustainable freshwater aquaculture in Brazil. Rev Aquac. 2020; 12(3):1495–517. https://doi.org/10.1111/raq.12393
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara RB et al.Vegan: Community Ecology Package [Internet]. R package version 2.0-8; 2013. Available from: http://CRAN.R-project.org/package=vegan
Olden JD, Kennard MJ, Pusey BJ. Species invasions and the changing biogeography of Australian freshwater fishes. Global Ecol Biogeogr. 2008; 17(1):25–37. https://doi.org/10.1111/j.1466-8238.2007.00340.x
Olden JD, Poff NL. Toward a mechanistic understanding and prediction of biotic homogenization. Am Nat. 2003; 162(4): 442–60. https://doi.org/10.1086/378212
Olden JD, Whattam E, Wood SA. Online auction marketplaces as a global pathway for aquatic invasive species. Hydrobiologia. 2020; 848:1967–79. https://doi.org/10.1007/s10750-020-04407-7
Padial AA, Vitule JRS, Olden JD. Preface: aquatic homogenocene -understanding the era of biological re-shuffling in aquatic ecosystems. Hydrobiologia. 2020; 847:3705–09. https://doi.org/10.1007/s10750-020-04413-9
Peres-Neto PR, Legendre P, Dray S, Borcard D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology. 2006; 87(10):2614–25. https://doi.org/10.1890/0012-9658(2006)87[2614:VPOSDM]2.0.CO;2
R Development Core Team. R: A language and environment for statistical computing [Internet]. Vienna, Austria: R Foundation for Statistical Computing; 2020. Available from: http://www.R-project.org
Rixon CAM, Duggan IC, Bergeron NMN, Ricciardi A, MacIsaac HJ. Invasion risks posed by the aquarium trade and live fish markets on the Laurentian Great Lakes. Biodivers Conserv. 2005; 14:1365–81. https://doi.org/10.1007/s10531-004-9663-9
Rocha CFD, Bergallo HG, Mazzoni R. Invasive Vertebrates in Brazil. In: Pimentel D, editor. Biological Invasions: Economic and Environmental Costs of Alien Plant, Animal, and Microbe Species. New York: Taylor & Francis; 2011. p.53–103.
Thomé PM, Paula MFPL, Rosa LMJ, Saroba CC. Caracterização da ictiofauna de um riacho de Mata Atlântica, no âmbito de uma central geradora de energia, Carmo-RJ, Brasil. Rev Interd Pens Cient. 2019; 5(5):804–19. http://dx.doi.org/10.20951/2446-6778/v5n5a61
Tuckett QM, Ritch JL, Lawson KM, Hill JE. Landscape-scale survey of non-native fishes near ornamental aquaculture facilities in Florida, USA. Biol Invasions. 2017; 19:223–37. https://doi.org/10.1007/s10530-016-1275-2
Vieira F, Rodrigues RR. A fauna de peixes dos afluentes do rio Paraíba do Sul no estado de Minas Gerais. MG Biota. 2010; 3(1):5–23.
Vitule JRS, Occhi TVT, Kang B, Matsuzaki SI, Bezerra LAV, Daga VS et al. Intra-country introductions unraveling global hotspots of alien fish species. Biodivers Conserv. 2019; 28(11):3037–43. https://doi.org/10.1007/s10531-019-01815-7
Vitule JRS, Skóra F, Abilhoa V. Homogenization of freshwater fish faunas after the elimination of a natural barrier by a dam in Neotropics. Divers Distrib. 2012; 18(2):111–20. https://doi.org/10.1111/j.1472-4642.2011.00821.x
Wickham H, Averick M, Bryan J, Chang W, McGowan LD, François R et al. Welcome to the Tidyverse. J Open Source Softw. 2019; 4(43):1686. https://doi.org/10.21105/joss.01686
Whittaker RH. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol Monogr. 1960; 30(3):279–338. https://doi.org/10.2307/1943563
Xiong W, Tao J, Liu C, Liang Y, Sun H, Chen K et al. Invasive Aquatic Plant (Alternanthera philoxeroides) facilitates the invasion of western Mosquitofish (Gambusia affinis) in Yangtze River, China. Aquat Ecosyst Health Manag. 2019; 22(4):408–16. https://doi.org/10.1080/14634988.2019.1700090
Authors

![]() André Lincoln B. Magalhães1
André Lincoln B. Magalhães1![]() ,
,![]() Luis Artur V. Bezerra2,
Luis Artur V. Bezerra2,![]() Vanessa S. Daga3,
Vanessa S. Daga3,![]() Fernando M. Pelicice4,
Fernando M. Pelicice4,![]() Jean R. S. Vitule3 and
Jean R. S. Vitule3 and![]() Marcelo F. G. Brito5
Marcelo F. G. Brito5
[1] Rua Professor Arduíno Bolívar, 80, Santo Antônio, 30350-140 Belo Horizonte, MG, Brazil. andrebiomagalhaes@gmail.com (corresponding author).
[2] Institute of Hydrobiology, Biology Centre of the Czech Academy of Sciences, České Budějovice, Czech Republic. larturr@yahoo.com.br.
[3] Laboratório de Ecologia e Conservação, Departamento de Engenharia Ambiental, Setor de Tecnologia, Universidade Federal do Paraná, Jardim das Américas, 81531-970 Curitiba, PR, Brazil. (VSD) vanedaga@yahoo.com.br, (JRSV) biovitule@gmail.com.
[4] Núcleo de Estudos Ambientais, Universidade Federal do Tocantins, Rua 03 Quadra 17, Jardim dos Ipês, 77500-000 Porto Nacional, TO, Brazil. fmpelicice@gmail.com.
[5] Laboratório de Ictiologia, Departamento de Biologia, Universidade Federal de Sergipe, Av. Marechal Rondon S/N, Jardim Rosa Elze, 49100-000 São Cristóvão, SE, Brazil. marcelictio@gmail.com.
Authors Contribution
André Lincoln Barroso Magalhães: Conceptualization, Formal analysis, Investigation, Methodology, Project administration, Writing-original draft, Writing-review and editing.
Luis Artur Valões Bezerra: Formal analysis, Methodology, Writing-review and editing.
Vanessa Salete Daga: Formal analysis, Methodology, Writing-review and editing.
Fernando Mayer Pelicice: Formal analysis, Methodology, Writing-review and editing.
Jean Ricardo Simões Vitule: Formal analysis, Methodology, Writing-review and editing.
Marcelo Fulgêncio Guedes Brito: Data Curation, Formal analysis, Writing-review and editing.
Ethical Statement
Fish individuals were collected by Sistema de Autorização e Informação em Biodiversidade (SISBIO – license number 61740).
Competing Interests
The authors declare no competing interests.
How to cite this article
Magalhães ALB, Bezerra LAV, Daga VS, Pelicice FM, Vitule JRS, Brito MFG. Biotic differentiation in headwater creeks after the massive introduction of non-native freshwater aquarium fish in the Paraíba do Sul River basin, Brazil. Neotrop Ichthyol. 2021; 19(3):e200147. https://doi.org/10.1590/1982-0224-2020-0147
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2021 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted June 22, 2021 by Emili Garcia-Berthou
Accepted June 22, 2021 by Emili Garcia-Berthou
![]() Submitted December 21, 2020
Submitted December 21, 2020
![]() Epub October 4, 2021
Epub October 4, 2021