Leandro Balboni1 ![]() , Facundo Vargas2
, Facundo Vargas2 ![]() and Darío Colautti3
and Darío Colautti3 ![]()
![]()
PDF: EN XML: EN | Cite this article
Abstract
The age and growth of Pseudoplatystoma corruscans, is analyzed on specimens landed in Puerto Antequera, Province of Chaco, Argentina. The study is based on length frequency distribution of 1192 individuals and growth marks of 293 pectoral spines. Previously to age assignation based on spines readings, we performed analyses that ruled out age-associated resorption of rings and corroborated the annual periodicity of mark formation. The average sizes of the radius of each ring were obtained, and the total length of fish were back-calculated to the time of the ring formation, by the regression model fitted between the total radius of the spines to the respective sizes of each fish. Such data showed a good fitting to growth models of von Bertalanffy, Gompertz and logistic for both sexes separately. Results indicate that the study of the species growth must be carried out for each sex separately and that the fishing regulations must consider this characteristic of the species since the current management guidelines could be promoting differential capture by sexes.
Keywords: Fisheries management, Growth models, Pectoral spine.
En este trabajo se analiza el crecimiento y la edad del Pseudoplatystoma corruscans, a partir de ejemplares desembarcados en el Puerto Antequera, provincia de Chaco, Argentina. El estudio se basó en la distribución de frecuencias de tallas de 1192 individuos y marcas de crecimiento en 293 espinas pectorales. Para asignar las edades a partir de las lecturas se realizaron análisis que permitieron descartar la reabsorción de anillos asociada a la edad y corroborar la periodicidad anual de la formación de marcas. Se obtuvieron los tamaños medios de los radios de cada anillo y se retrocalcularon las tallas al momento de su formación, mediante la relación del radio total de las espinas con las tallas respectivas de cada pez. Dichos datos mostraron un buen ajuste a las curvas de crecimiento de von Bertalanffy, Gompertz y logística para ambos sexos por separado. Los resultados demostraron que el estudio del crecimiento de la especie debe realizarse para cada sexo por separado y que las normas pesqueras deben contemplar esta característica de la especie ya que las actuales pautas de manejo podrían estar promoviendo la captura diferencial por sexos.
Palavras-chave: Espina pectoral, Manejo pesqueroo, Modelos de crecimiento.
Introduction
Pseudoplatystoma corruscans (Spix & Agassiz, 1829), known in the confluence of the Paraná and Paraguay rivers region as “surubí pintado”, is a species of great biological importance, due to its role as top predator, and it is also of high economic relevance in the centers where commercial and sport fishing activities take place throughout its distribution (Mateus, Petrere, 2004). It shows no external sexual dimorphism, and like many large species of the family Pimelodidae, it makes complex migratory movements for reproduction (Lowe-McConnel, 1975). Although the species has been studied by various scientific disciplines such as genetics (Barreta Pinto, 2005; Revaldaves et al., 2005; Pereira et al., 2009, Baggio et al., 2016, Escobar Lizarazo, 2016), physiology (Lundstedt et al., 2004), reproductive biology (Brito, Bazzoli, 2003; Godinho et al., 2007), conservation (Mello et al., 2009), and fishing (Cordiviola, 1966), its demographic aspects, linked to age and growth, have been estimated in few opportunities. These features are essential for the development of management guidelines that make possible the sustainable use of the species as was assessed by Mateus, Petrere (2004), who also determined the degree of exploitation of the species in the Pantanal, or by de Souza et al. (2012), that proposed a population dynamic model to optimize its management in Brazil.
In the literature on the growth of P. corruscans, the analyses have been carried out by reading the growth rings in bone structures and from distributions of size frequency (Resende et al., 1995; Godinho et al., 1997, 2007; Mateus, Petrere, 2004; Bechara et al., 2007; Navia et al., 2007; Oldani et al., 2013). The uses of size frequency distribution or spines as non-lethal methods of age determination and growth should be analysed and validated for this species, because the data can be obtained from specimens that are released after capture, what would be useful for the management and conservation of this species. In the cases where the von Bertalanffy curve was fitted to length-age data, the parameters obtained showed relatively disparate results in asymptotic length and growth constant. Despite such differences could be linked to the methodology and/or the differences in growth of this species depending on the study area, there were also intersexual differences in the referred parameters when growth models were fitted separately to each sex.
In Puerto Antequera (Province of Chaco, Argentina) P. corruscans represents more than 40% of the commercial fishery landings. A similar situation occurs in the landing sites of the city of Corrientes (Province of Corrientes, Argentina) located just on the opposite bank of the Paraná River, where also the sport-recreational fishing of the species represents a tourist attraction at national and international level (Bechara et al., 2007).
There is a shared regulation among provinces regarding the fishing of this species in its commercial and sports-recreational versions. The management guidelines for the species have been prepared considering general knowledge on its biology or based on P. corruscans data from other sectors of the basin, including those associated with the maximum and minimum catch sizes.
The available information regarding the age and growth of P. corruscans in the area where the main fisheries for this species operate in Argentina is still scarce (Bechara et al., 2007; Oldani et al., 2013). Despite differential growth by sex has been mentioned for the species (Bonetto et al., 1965; Cordiviola, 1966), this sexual dimorphism has not been taken into account in setting fishing regulations, likely because there is still no validated study on P. corruscans growth that considers sexual differences, in the area.
The main goal of this paper is to analyze the age and growth of P. corruscans, in the confluence area of the Paraná and Paraguay rivers, analyzing its temporal dynamics, as well as likely influencing environmental variables and the potential existence of differences in growth of each sex. Finally, the possible implications of results on species management are discussed.
Material and methods
The Government of the Province of Chaco has been monitoring commercial fishing through Dirección de Fauna y Áreas Naturales Protegidas (DFyANP, SDTyA), constantly since 2010 in the main landing port, the San Pedro Pescador fishing district, known as Puerto Antequera, 27о27’44.2”S 58о52’00.4”W. The fishing areas where the landings come from are shown in Fig. 1. In this sector, the Paraná River has a single watercourse approximately 1.5 km wide. Its bottom is sandy, with two well-defined channels, a deeper one on the left margin that can reach 25 m, and another shallower on the right margin of about 15–20 m depth (Rayano, 2003).
The fishing zones are areas approximately 1,500 m long by 400 m wide called “canchas”. Their major axes are parallel to the riverbed and they have no obstacles such as logs or roots that may be deposited in the riverbed. There are about four canchas used in the area.
The studied specimens were obtained from catches of commercial fishing carried out with hand-woven nylon thread nets, 150 to 300 m long and 3 m high. They have different sizes of mesh opening, depending on the target species and the time of the year, from 18 cm to 33 cm between opposite knots, and operate drifting along the canchas. Other less used fishing modalities for this species are rods of Tessaria integrifolia (river alder) called “simbras”, digged in the coasts, with a twine and a hook, and bottom long-lines with hooks of different sizes. Measurements of P. corruscans specimens and sample extractions were carried out within the monitoring program of the Province of Chaco in Puerto Antequera from February 2010 to February 2012. Total length (TL), measured from the extreme of the snout to the end of the stretched caudal fin, with a precision of one millimeter, and sex of each specimen were recorded. Pectoral spines were removed with a proximal cross section or by disarticulation from the pelvic girdle; then they were kept dry in paper envelopes. In addition, monthly mean air temperature (Temp.), daylight hours (Hs.L.), and hydrometric level (HL) records were obtained for the study area between 2000 and 2012.

FIGURE 1| Geographic location of Antequera port, the study area, and detail of the fishing area.
The size frequency distributions of the specimens, grouping them by 50 mm intervals and discriminated by sex were obtained. The differences between both distributions were determined using the Chi Square Test.
For age determination, the spines were immersed for 2 h in a 10% sodium hypochlorite solution and the excess epidermal and muscular tissue was removed with the help of a stiff-bristle brush and tweezers. From the proximal portion of the spine body, 3 to 5 sections 0.5 to 1 mm thick were cut at the beginning of the foramen (Bisbal, Gomez, 1986), transversely to the major axis of the spine, with a diamond disc (Fig. 2). Each section was fixed to a slide with contact adhesive and polished with waterproof sandpaper with different grain size (from 300 to 1,000), until reaching 0.2-0.3 mm thickness, to eliminate the striae produced during the cut and clearly identify the marks.

FIGURE 2| A. Detail of a spine of Pseudoplatystoma corruscans in upper and lower views and B. detail of the proximal portion of the spine. Lines indicate the cuts positions. C. View of spine cut showing the ventral radius of the dorso-ventral axis, where distances from the estimated center of the spine foramen to the edge (Rt) and to each growth ring (Rn) were measured.
To perform spine readings, the slides were immersed in water in a Petri dish and the identification of marks was performed under an Olympus SZX10 stereomicroscope with incident light and camera Olympus Q color 5. Measurements were made with the software of digital analysis Q-Capture pro7. For the identification of marks, the criterion of Casselman (1983) was followed, recognizing wide opaque areas of fast growth and narrow translucent areas of slow growth. Double and/or discontinuous rings were discarded. Only the spines in which the number of growth rings was repeated in at least two of the prepared sections of the spine were used. The distances from the estimated center of the spine foramen to the edge (Rt) and to each growth ring (Rn) were obtained over the ventral radius of the dorso-ventral axis of the spine cut, according to Penha et al. (2004) (Fig. 2).
As the pectoral spine is a conical structure, cuts made in the proximal-distal axis have different Rt, therefore, to compensate the change in shape, we calculated the percentage corresponding to the distance of each ring to the center in each cut, with respect to the Rt, according to the formula:

Then, the percentage values of each ring within the spine were transformed to distances (Rn*) according to the following formula:

Where Rt* is the total radius estimated for a fish of a determined TL, through the equation of the line fitted to the Rt-TL pairs according to the formula:

Given that the expansion of the central foramen, which determines the possible loss of the first growth rings, is common in the spines of the oldest specimens, we used the method proposed by Penha et al. (2004) to evaluate the occurrence or not of this phenomenon, which could be the cause of erroneous age assignments. This technique assumes that if the first rings near the center of the spine in the oldest specimens were resorpted by the foramen, the oldest ones would have a significantly longer radius for the first ring than the youngest specimens. This assumption was verified by means of a one-way ANOVA test, defining age as a factor and the mean radius of the first ring as the response variable.
To confirm whether ring formation periodicity is annual and thus, validate the age assignments, the percentage marginal increase (MI%) was calculated according to the formula:

Where Rn*max is the corrected radius length of the last ring measured.
The monthly mean values of MI% were estimated to know the temporal dynamics of ring formation and, therefore, to evaluate the possible existence of differential growth throughout the annual cycle. To quantify the explained variation of MI% by the environmental forcing variables, the monthly average values of MI% were regressed by mean of a generalized linear model (GLM), to the monthly average values of air temperature (Temp.), daylight hours (Hs.L.) and hydrometric levels (HL), (period 2000–2012).
The average Rn* for each age and its deviations were estimated and then converted to total lengths (back calculation) from the best fit function to the data pairs Rt*-TL.
By means of the FSA (Fisheries Stock Assessment) (Ogle et al., 2020) R package, the age-total length key for females and males were fitted by nonlinear minimum square method to the von Bertalanffy (VBF), Gompertz (GPZ) and logistic (LOG) growth models according to the following equations:
Von Bertalanffy

Gompertz

Logistic

Where:
TLt = Total length (mm) of fish at the t moment.
t = time (years).
TL∞ = asymptotic total length (mm).
k = growth constant.
t0 = hypothetical time (years) at zero length.
To select the best model, the Akaike Information Criterion corrected index (AICc) was applied.
Based on the fitted growth curves and the mean total lengths when the 50% and 100% of females and males of P. corruscans are adults respectively (TL50 and TL100) reported in the study by Barzotto et al. (2016), the corresponding ages of first maturity were calculated.
Finally, the parameters and curves of the von Bertalanffy model obtained by other authors at different locations of the natural distribution of the P. corruscans, were compiled in a table to compare them with those obtained in this work.
Results
During the study period 1,192 specimens of P. corruscans were measured. The size distribution was bimodal in the range between 57 and 1,640 mm (TL). The best represented TL were in the range of 850–1,550 mm, and the modal TL were 1,000 and 1,300 mm., respectively. Size frequency distribution considering sexes show that, although both genders contribute to the general scheme, males predominate in the first mode and females in the second (Fig. 3). The Chi Square Test confirmed the existence of significant differences between the sizes of females and males (Chi Square=2133.82; gl.=26; p<0.01).
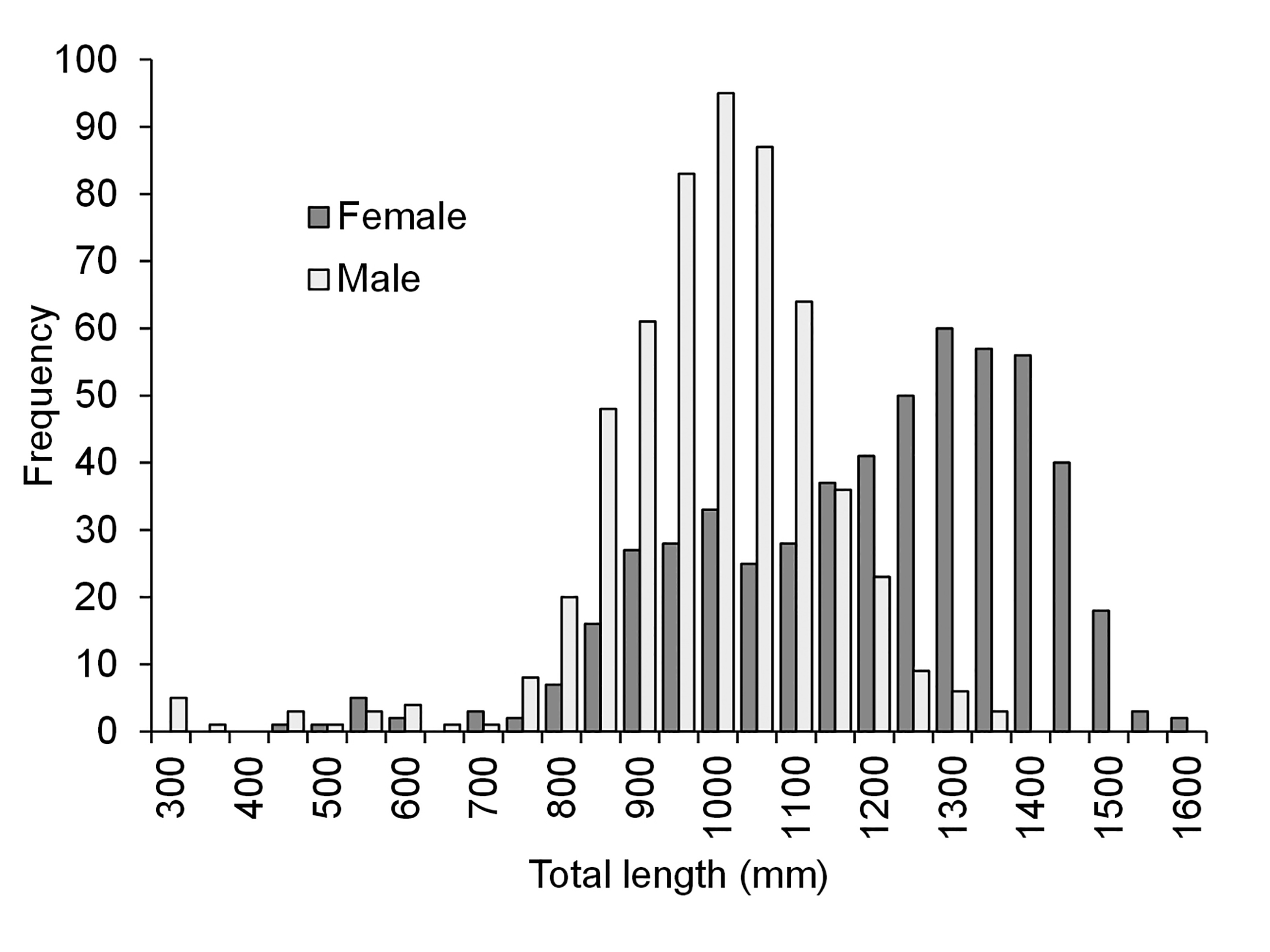
FIGURE 3| Total length (TL) frequency distribution of Pseudoplatystoma corruscans specimens discriminated by sexes.
The analysis of data from the readings of 293 spines indicate that no significant differences were found in the first ring mean size, among the specimens with different number of rings on their spines, F=1.85; gl=6; p=0.09 (Fig. 4). Therefore, the occurrence of the phenomenon of resorption of the rings closest to the center of the spine, which could be associated with the growth of individuals, was ruled out.
The analysis of the marginal increment showed a seasonal growth pattern. It begins with a constant rapid increase of the edge of the spine from February to June. At that time, marginal increment starts to fall abruptly until August, when, although it continued to decrease, it was gradual, reaching its lowest level in January. This confirms that the formation of the marks on the spine has an annual frequency (Fig. 5).
The GLM results indicated that 94% (p<0.01) of the observed variation can be explained through the adjusted model, being identified as significant predictive variables the light hours in the first place, and the hydrometric level in the second place. Temperature was not selected by the GLM as a significant variable explaining the MI% variation. The parameters of GLM fitted are shown in Tab. 1 and the monthly variations of MI% and their explanatory variables are plotted in Fig. 5.
The back calculation models for the pair of data TL-Rt* per sex were the following: TLmales= 250.68 Rt*+240.23 – R²=0.77 – N=74, and TLfemales= 351.75 Rt*+46.64 – R²=0.82 – N=95.
Tab. 2 shows the mean values (Rn*mean) and standard deviations (SD) of the Rn* recorded for each sex, and their respective transformations to TL values. The three growth models fitted significantly to the age-length data for both sexes (r2>0.98; p<0.01). The respective fitting parameters for each model and sex are shown in Tab. 3 and plotted in Fig. 6. The AIC method indicated that the best model was the logistic, followed by the Gompertz and the von Bertalanffy models for both, females and males. However, the difference in AIC between models was very small.

FIGURE 4| Distances from the center of the spine to the first ring (black diamonds) in individuals of Pseudoplatystoma corruscans with different numbers of growth rings and their respective mean values (squares). The horizontal line represents the general mean value.
TABLE 1 | Parameters of general linear model (GLM) fitted to analyze the monthly marginal increment variations (MI%) and their explanatory variables, daylight hours (Hs.L.) and hydrometric levels (HL).
Parameter | Value | Std. Err. | p |
Intercept | 106,877 | 34,333 | 0,014 |
Hs.L. | -12,181 | 3,068 | 0,004 |
HL. | 17,536 | 6,786 | 0,032 |

FIGURE 5| Monthly averages of the percentage of marginal increase (MI%) in spines of Pseudoplatystoma corruscans, hours of day light (Hs.L.) and hydrometric levels of the Paraná River (HL) during the annual cycle.
TABLE 2 | Mean values and standard deviations of distances from core to each annual growth marks (rings) and back-calculated mean total lengths and standard deviations at age (individuals) for Pseudoplatystoma corruscans, per sex. Abbreviations: Rn*=corrected spine radius length TL=total length (cm), SD=standard deviation.
| Males |
| Females | ||||||
| Spines | Individuals |
| Spines | Individuals | ||||
Ring | Rn* | SD | TL | SD |
| Rn* | SD | TL | SD |
1 | 1.28 | 0.36 | 561.3 | 156.7 |
| 1.40 | 0.40 | 539.8 | 155.3 |
2 | 1.89 | 0.49 | 714.7 | 185 |
| 1.98 | 0.47 | 744.6 | 176.3 |
3 | 2.39 | 0.49 | 839.9 | 173.4 |
| 2.49 | 0.43 | 922 | 159.1 |
4 | 2.72 | 0.39 | 922.3 | 133.8 |
| 2.92 | 0.40 | 1073.3 | 145.9 |
5 | 2.90 | 0.47 | 966.2 | 157.4 |
| 3.08 | 0.35 | 1130.7 | 127.1 |
6 | 2.80 | 0.68 | 941.7 | 227.5 |
| 3.19 | 0.27 | 1168.6 | 141.2 |
TABLE 3 | Parameters growth models Von Bertalanffy (VBF), Gompertz (GPZ) and logistic (LOG), fitted to the mean total length (TL) at age for females and males of Pseudoplatystoma corruscans. Asymptotic total length mm (TL∞), growth constant (k), = hypothetical time at zero length in years (t0), determination coefficient (r2), Akaike information criterion corrected index (AICc).
Sex | Model | TL∞ (mm) | k | t0 (years) | r2 | AICc |
| VBF | 1318.330 | 0.340 | 0.548 | 0.995 | 98 |
Females | GPZ | 1252.368 | 0.529 | 0.705 | 0.997 | 94 |
| LOG | 1216.173 | 0.711 | 1.337 | 0.999 | 90 |
| VBF | 992.44 | 0.505 | 0.647 | 0.985 | 99 |
Males | GPZ | 988.155 | 0.643 | 0.147 | 0.988 | 97 |
| LOG | 978.264 | 0.781 | 0.651 | 0.991 | 96 |

FIGURE 6| Back calculated total length from the pectoral spines readings (B-C TL) and back calculated mean total lengths at age (B-C mean TL) for Pseudoplatystoma corruscans females (A) and males (B), and the respective fitted growth models: Von Bertalanffy (VBF), Gompertz (GPZ) and logistic (LOG).
The estimated ages of first maturity for P. corruscans combining the growth results of this study with the TL50 and TL100 reported by Barzotto et al. (2016), were 1.2 and 2.5 years for females and 1.47 and 3.16 year for males, respectively. These ages could be considered very young for maturity in a big size fish like P. corruscans, however it should be also considered that the estimated ages represent approximations because were obtained from reference lengths of individuals belonging to a lower latitude population.
In Tab. 4 the parameters of the growth models of von Bertalanffy obtained in this study are compared to those obtained by other authors.
Discussion
The total length frequency distributions of P. corruscans landings discriminating by sex, clearly shows that larger sizes are mostly dominated by females and intermediate sizes correspond mainly to males. Although the abundance of specimens by size class may be significantly influenced by the types of gear used in the fishery, the sex ratios of each size class would not be as strictly subject to gear selectivity and would therefore reflect the approximate sex ratio for each size class. In many species of fish, females reach larger sizes (Nikolski, 1963) than males, and in P. corruscans this biological trait seems to be especially expressed as was observed by other authors (Bonetto et al., 1965; Cordiviola, 1966; Resende et al., 1995; Mateus, Petrere,2004). This scenario has major implications when analyzing the growth of this species since, if the maximum sizes reached by each sex are significantly different, the respective growth dynamics may also be different. The absence of males within the intervals of size close to the maximum in the P. corruscans population analyzed, indicates that they have a smaller infinite size than females.
The analysis of marginal increase indicated that the formation of growth marks has an annual periodicity and therefore each mark represents one year. The time of appearance of the marks turned out to be the second half of winter, and the sharp reduction in the value of the index in the spring months indicates the start of a new period of growth. This annual dynamic is in tune with the seasonal variations in light hours and, secondly, with the hydrological behavior of the river. The aforementioned means an important difference regarding the results obtained by Resende et al. (1995) for the population from the Pantanal. These authors studied the annual growth dynamics and found that it is strongly associated with the annual hydrological cycle of drought and flood that condition the space and availability of food. On the contrary, in the Paraná River besides light hours, and hydrology, other aspects of the life history of the P. corruscans, such as migrations and reproduction, have influence on the growth biology as mentioned by Bonetto et al. (1965), Cordiviola (1966), and Oldani et al. (2013), but an interesting, obtained result was that temperature is not an environmental factor conditioning significantly the seasonal species growth.
TABLE 4 | Parameters of the von Bertalanffy curve obtained by different authors for Pseudoplatystoma corruscans and those corresponding to this work.
Author | Sex | TL∞. (mm) | k | t0 (years) | Study area |
Resende et al. (1995) | Females | 1115 | 0.27 | 1.07 | Rio Miranda, Pantanal, Mato Grosso do Sul, Brazil |
Males | 1001 | 0.21 | 2.58 | ||
Mateus, Petrere (2004) |
| 1830 | 0.08 | 3.27 | Rio Cuiabá, Pantanal, Mato Grosso do Sul, Brazil |
Bechara et al. (2007) |
| 1617 | 0.18 | 2.12 | Río Paraná, Chaco, Argentina |
Vargas (2010) | Females | 1448 | 0.22 | 1.09 | Río Paraná, Chaco, Argentina |
Males | 944 | 0.47 | 0.73 | ||
Oldani et al. (2013) |
| 1850 | 0.06 | 3.40 | Río Paraná, from Ituzaingó to Santa Fé, Argentina |
This paper | Females | 1318.33 | 0.34 | 0.54 | Río Paraná, at confluence of Paraná and Paraguay rivers, Argentina and Paraguay |
Males | 992.44 | 0.51 | 0.65 |
The age assignment in this species was carried out by different methods, based on the readings of hard structures and population analysis. However, this issue is still controversial. In this sense, Oldani et al. (2013), suggest that studies carried out by various authors based on age dating from spines (Bechara et al., 2007; Cordiviola, 1966; Mateus, Petrere, 2004; Vargas, 2010), and/or vertebral bodies (Navia et al.,2007), have overestimated growth because they do not take into account the resorption existing in those hard structures. In an attempt to resolve the issue, Oldani et al. (2013) carried out a study of growth by reading the radii of spines and by pseudocohort analysis, validating the results based on the coincidence found in the age assignment by both methods. However, this coherence was achieved by applying an age correction in the case of fish older than 5 years. It is based on the fact that the growth rings near the foramen are resorpted in the spines of large fish, and therefore years must be added depending on the thickness of the spine. Although Oldani et al. (2013) obtained evidence for this resorption phenomenon from a percentage relationship of distances between the foramen, the mark and the edge of the spine, the formula is not specified in their work. Thus, in this study, such estimates could not be made for comparative purposes. The option was to apply proven methods to evaluate the resorption of rings near the foramen in siluriforms. As a result, it was observed that this phenomenon is not generalized in the case of the P. corruscans, at least until the sixth year of life. This is not a minor detail since it implies assignments of different ages for the size-groups belonging to each cohort, resulting in growth curves with significantly different parameters.
When the three growth models were fitted to the data all of them demonstrated to be adequate to describe the individual growth of the species for females and males. Despite the logistic curve was selected as the best by the AICc, and the von Bertalanffy model was positioned as the last, it was used to compare with other works because was the only applied for the species before this study. In addition, AICc support for a particular model was not high, especially for males. The growth curves obtained for males and females were similar to those adjusted by Resende et al. (1995) and Vargas (2010). This may be due in part to the fact that these authors chose to fit curves for each sex separately, which solves a large part of the variation in sizes by age. Consequently, more precise settings and more reliable parameters can be obtained. In contrast, in those cases in which a single curve was adjusted for the species (Mateus, Petrere, 2004; Bechara et al., 2007; Oldani et al., 2013), the resulting values of infinite size were far from the observed maximum sizes, and negative t0 values were higher than 2 and 3 years, which has the effect of forcing the model to assume low growth rates (k). However, the assignment of ages to the different size groups, back-calculated or found in the population, also has a direct effect on the growth estimates. Thus, the size-age keys that were made from the reading of hard structures without considering the reabsorption of marks as a phenomenon that eliminates the first growth marks, resulted in similar curves, while those such as the ones reported by Oldani et al. (2013) resulted in a slower growth.
Beyond the differences associated with methodological issues, another question that arises when comparing the curves is that there seems to be a difference in growth between P. corruscans of the middle Paraná and those of the Pantanal. In this sense, when comparing the curves, it is observed that individuals of the populations located further south generally reach larger sizes in less time. This differential growth could be explained by the phenomenon of latitudinal counter gradient described by Conover (1992), which states that, in species with a wide latitudinal distribution exposed to different seasonal regimes, the duration of the growing season differs, and therefore also the growth patterns. Besides, it is likely the growth differences found could be also linked to populations genetics.
Considering together the frequency length distributions by sex of landings (Fig. 3), growth curves (Fig. 6) and the lengths and ages of maturity, it is possible to deduce that the fishery of the confluence area, currently directs its fishing effort to individuals of 3 years or older that are almost all 100% adults according to the reference lengths reported by Barzotto et al. (2016). Nevertheless, the differential growth of males and females indicates that all research on the growth of this species, focused on obtaining parameters to develop population models and establish management guidelines, should consider this sexual dimorphism. In this sense, current fishing regulations for this species that establish a minimum of 1,000 mm and maximum of 1,200 mm catch size deserve to be discussed, since they could have negative consequences on the dynamics of the population. This situation would occur because this regulation promotes the differential extraction of specimens by sex, altering the natural relationship between males and females. Furthermore, although the arguments of the regulations that limit the maximum sizes tend to protect the “mega-breeders”, it only protects female “mega-breeders”, and directs the extraction to male “mega-breeders”, which could cause an effect opposite to the one expected. In fact, Santos et al. (2012), after practicing various management models for P. corruscans, proposes to protect the oldest and largest individuals, and the youngest as a better alternative. However, although the models were correct, their work is developed using common growth parameters without discriminating by sex, falling into the same problem raised above of promoting selective fishing by sex. According to this study, the strategy of defining a group of harvestable sizes in the population, based on simultaneously protecting immature individuals and older mature individuals to promote sustainability and a better use of the resource, in the case of P. corruscans would not have the expected benefits. From this study, emerges the necessity of further population-level analyses that incorporate differential growth by sexes, in addition to studies weighing the relative contributions of male and female mega-breeders to population growth. Such population-level analyses are needed to update recommend size limits and management strategies considering the growth differences between sexes observed in this work. This could contribute to improve the management guidelines to maximize the sustainability of the fishery.
Acknowledgments
The authors would acknowledge to Leandro Bher, Monica Bher y Cristian Zarnichonwoscky for the fish sampling, to Dr. Sebastian Sanchez and Dra Natalia Silva, Dr. Juan Jose Rosso for their kind support and advice, to Vivian Yorojo Moreno for their help with R software estimations, to the authorities of DFyANP, SDTyA and DPP, MAGyP for supporting landings sampling program and the study development.
References
Baggio RA, Moretti CB, Bialetzki A, Boeger WA. Hybrids between Pseudoplatystoma corruscans and P. reticulatum (Siluriformes: Pimelodidae) previously reported in the Upper Paraná River are likely escapes from aquaculture farms: evidence from microsatellite markers. Zoologia. 2016; 33(2):e20150200. https://doi.org/10.1590/S1984-4689zool-20150200
Barreta Pinto J. Variabilidad genética de Pseudoplatystoma fasciatum y Pseudoplatystoma trigrinum en el Alto Madera de la Amazonia boliviana por análisis comparativo del polimorfismo de longitud de secuencia de intrones (EPIC-PCR) y electroforesis enzimática. [Master Dissertation]. La Paz: Universidad Mayor de San Andrés; 2005. Available from: https://horizon.documentation.ird.fr/exl-doc/pleins_textes/divers17-11/010037080.pdf
Barzotto E, Oliveira M, Mateus L. Reproductive biology of Pseudoplatystoma corruscans (Spix and Agassiz, 1829) and Pseudoplatystoma reticulatum (Eigenmann and Eigenmann, 1889), two species of fisheries importance in the Cuiabá River Basin, Brazil. J. Appl Ichthyol. 2016, 33(1):29–36. https://doi.org/10.1111/jai.13162
Bechara JA, Vargas F, Quintana CF. Biología pesquera de las principales especies de importancia económica en el área de la confluencia de los ríos Paraná y Paraguay. Subsecretaría de Pesca y Acuicultura de la Nación. Buenos Aires; 2007. Available from: https://www.magyp.gob.ar/sitio/areas/pesca_continental/proyectos/_archivos//000001-Biologia%20pesquera%20de%20las%20principales%20especies%20de%20importancia%20economica%20en%20el%20area%20de%20la%20confluencia%20de%20los%20rios%20parana%20y%20paraguay%202006-2007.pdf
Bisbal G, Gómez S. Morfología comparada de la espina pectoral de algunos Siluriformes bonaerenses (Argentina). Physis. 1986; 44:81–93.
Bonetto AA, Piganalberi C, Cordiviola E. Notas preliminares para un estudio biológico y pesquero del “surubí” (Pseudoplatystoma corruscans y P.fasciatum) en el Paraná Medio. Pisces Pimelodidae. Anais do II Congreso Latino-Americano de Zoología. São Paulo. 1965; II:125–29.
Brito MFG, Bazzoli N. Reproduction of the surubim catfish (Pisces, Pimelodidae) in the São Francisco River, Pirapora Region, Minas Gerais, Brazil. Arq Bras Med Vet Zootec. 2003; 55(5):624–33. https://dx.doi.org/10.1590/S0102-09352003000500018
Casselman JM. Age and growth assessment of fish from their calcified structures – techniques and tools. NOAA Technical Report NMFS. 1983; 8:1–17. Available from: https://www.st.nmfs.noaa.gov/spo/SPO/tr8opt.pdf
Conover DO. Seasonality and the scheduling of life history at different latitudes. J Fish Biol. 1992; 41:161–78. https://doi.org/10.1111/j.1095-8649.1992.tb03876.x
Cordiviola E. Nuevos aportes al conocimiento de la biología pesquera del “surubí” (Pseudoplatystoma corruscans) en el Paraná medio (Pisces, Siluriformes). Physis 1966; XXVI. (71):237–44.
Escobar Lizarazo M, Taphorn D. Manejo de las poblaciones de bagres Pseudoplatystoma fasciatum y P. tigrinum (Siluriformes, Pimelodidae) del Orinoco, a partir de análisis de variabilidad genética. Mem Fund La Salle de Cienc Nat. 2016; 164:79–100. Available From: https://aquatic-experts.com/AQUATIC-EXPERTS%20PDF/Donald%20Taphorn%20Reprints/2006%20Pseudoplatystoma%20variabilidad%20genetica.pdf
Godinho AL, Kynard B, Godinho HP. Migration and spawning of female surubim (Pseudoplatystoma corruscans, Pimelodidae) in the São Francisco river, Brazil. Environ Biol Fish. 2007; 80:421–33. https://doi.org/10.1007/s10641-006-9141-1
Godinho HP, Godinho AL, Miranda MTO, Santos JE. Fisheries and biology of the surubim Pseudoplatystoma corruscans in the São Francisco River at Pirapora, MG, Brazil. In: Miranda MTO, editor. Surubim. IBAMA, Brasília, Brazil, 1997; 27–42.
Lowe-McConnel RH. Fish communities in tropical freshwaters: their distribution, ecology and evolution. London: Logman, 1975.
Lundstedt L, Melo J, Moraes G. Digestive enzymes and metabolic profile of Pseudoplatystoma corruscans (Teleostei: Siluriformes) in response to diet composition. Comp Biochem Phys B. 2004; 137:331–39. https://doi.org/10.1016/j.cbpc.2003.12.003
Mateus LA, Petrere Jr M. Age, growth and yield per recruit analysis of the pintado Pseudoplatystoma corruscans (Agassiz, 1829) in the Cuibá River basin, Pantanal Matogrossense, Brazil. Braz J Biol. 2004; 64(2):257–64. https://doi.org/10.1590/S1519-69842004000200011
Mello PH, Venturieri RLL, Honji RM, Moreira RG. Threatened fishes of the world: Pseudoplatystoma corruscans (Agassiz, 1829) (Siluriformes: Pimelodidae). Environ Biol Fish. 2009; 85:359–60. https://doi.org/10.1007/s10641-009-9508-1
Navia C, Van Damme PA, Nuñez J. Aspectos biológicos de Pseudoplatystoma fasciatum y P. corruscans (PISCES, Pimelodidae) en la laguna Cáceres (Pantanal boliviano). In: Feyen J, Aguirre LF, Moraes M, editors. Congreso Internacional sobre Desarrollo, Medio Ambiente y Recursos Naturales: Sostenibilidad a múltiples niveles y escalas”, Cochabamba, Bolivia. 2007. p.1243–51.
Nikolski GV. The ecology of fishes. London: Academic Press. 1963.
Ogle DH, Wheeler P, Dinno A. 2020. FSA: Fisheries Stock Analysis. R package version 0.8.31. https://github.com/droglenc/FSA
Oldani NO, Ecclesia O, Baigún CRM. Edad, crecimiento, mortalidad e incidencia de las variaciones del nivel hidrométrico del río Paraná en la abundancia del surubí pintado (Pseudoplatystoma corruscans). Museo Provincial de Ciencias Naturales Florentino Ameghino. Rev Biológica. 2013; 16:25–38. Available from: https://ri.conicet.gov.ar/handle/11336/8793
Penha J, Mateus L, Petrere M. A Procedure to improve confidence in identification of the first annulus in fin-spines of fishes. Fisheries Manag Ecol. 2004; 11:135–37. https://doi.org/10.1046/j.1365-2400.2003.00374.x
Pereira LHG, Foresti F, Oliveira C. Genetic structure of the migratory catfish Pseudoplatystoma corruscans (Siluriformes: Pimelodidae) suggests homing behavior. Ecol Freshw Fish. 2009; 18:215–25. https://doi.org/10.1111/j.1600-0633.2008.00338.x
Rayano MF. Geomorfología y sedimentología del Río Parana en el área del segundo puente interprovincial Chaco-Corrientes. [Master Dissertation]. Facultad de Ingeniería. Universidad Nacional del Nordeste; 2003.
Resende EK, Catella AC, Nascimento FL, Palmeira SS, Candido RA, Lima MS, Almeida VLL. Biología do curimbatá (Prochilodus lineatus), pintado (Pseudoplatystoma corruscans) e cachara (Pseudoplatystoma fasciatum) na bacia hidrográfica do rio Miranda, Pantanal, Mato Grosso do Sul, Brasil. EMBRAPA-CPAP. Bol Pesqui. 1995; 2:5–75. Available from: https://www.infoteca.cnptia.embrapa.br/bitstream/doc/789558/1/BP02.pdf
Revaldaves E, Pereira LHG, Foresti F, Oliveira C. Isolation and characterization of microsatellites loci in Pseudoplatystoma corruscans (Siluriformes: Pimelodidadae) and cross-species amplification. Mol Ecol Notes. 2005; 5:462–65. https://doi.org/10.1111/j.1471-8286.2005.00883.x
Santos RVR, Martins S, Pompeu P. An individual-based model for evolutionary effects of selective fishing applied to Pseudoplatystoma corruscans. Physica A: Statistical Mechanics and its Applications. 2012; 391:5112–20. https://doi.org/10.1016/j.physa.2012.05.037
De Souza AA, Martins SGF, Pompeu PS. An individual-based model applied to the study of different fishing strategies of Pintado Pseudoplatystoma corruscans (Agassiz, 1829). BioSystems 2012; 110:51–59. https://doi.org/10.1016/j.biosystems.2012.07.001
Vargas F. Informe final pesquería. Dirección de fauna y áreas naturales protegidas, Subsecretaria de Recursos Naturales. Ministerio de la Producción y Ambiente. Chaco; 2010. Available from: https://docs.google.com/open?id=0B2nds01wkdEfUlJBSXhlV05TTEtHZnNVTHRmSWNDUQ
Authors

Leandro Balboni1 ![]() , Facundo Vargas2
, Facundo Vargas2 ![]() and Darío Colautti3
and Darío Colautti3 ![]()
![]()
[1] Laboratorios de Pesca Continental, Dirección de Planificación Pesquera, Ministerio de Agricultura, Ganadería y Pesca (DPP, MAGyP), Alférez de Navío, Francisco Pareja 125, C1107AQA Ciudad Autónoma de Buenos Aires, Buenos Aires, Argentina.
(LB) leanbalboni@yahoo.com.ar.
[2] Departamento de Fauna, Dirección de Fauna y Áreas Naturales Protegidas, Subsecretaria de Ambiente (DFyANP, SDTyA), Remedios de Escalada 46, CP 3500, Resistencia, Chaco, Argentina. (FV) vargasfacundo@yahoo.com.ar.
[3] Instituto de Limnología Dr. R. Ringuelet, CONICET, Universidad Nacional de LaPlata (ILPLA), Boulevard 120 y 62, CP 1900 – CC 712, La Plata, Buenos Aires, Argentina. (DC) colautti@ilpla.edu.ar (corresponding author).
Authors Contribution
Leandro Balboni: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Writing-original draft, Writing-review and editing.
Facundo Vargas: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Writing-original draft, Writing-review and editing.
Darío Colautti: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Supervision, Writing-original draft, Writing-review and editing.
Ethical Statement
Information of animals analyzed in our study was obtained by the Chaco Province fisheries Authority from landings of the artisanal fishery. Following the provincial normative and regulations.
Competing Interests
The authors declare no competing interests.
How to cite this article
Balboni L, Vargas F, Colautti D. Age and growth of Pseudoplatystoma corruscans (Siluriformes: Pimelodidae) at the confluence of the Paraná and Paraguay rivers. Neotrop Ichthyol. 2021; 19(2):e200101. https://doi.org/10.1590/1982-0224-2020-0101
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2021 The Authors.
Diversity and Distributions Published by SBI
![]() Submitted September 24, 2020
Submitted September 24, 2020
![]() Accepted March 14, 2021 by Franco Teixeira de Mello
Accepted March 14, 2021 by Franco Teixeira de Mello
![]() Epub Jun 21, 2021
Epub Jun 21, 2021