Kyara Martins Formiga[1][2][4] ![]()
![]() , Jacqueline da Silva Batista[1][2][4]
, Jacqueline da Silva Batista[1][2][4] ![]() and José Antônio Alves-Gomes[1][3][4]
and José Antônio Alves-Gomes[1][3][4] ![]()
PDF: EN XML: EN | Supplementary: S1, S2, S3, S4 | Cite this article
Abstract
The migratory catfish Brachyplatystoma vaillantii is one of the most important fishery resources in the Amazon. Intense capture occurs associated to its life cycle. In order to know the genetic status, we sequenced the mitochondrial DNA control region from 150 individuals of B. vaillantii, collected in five fishing landing locations, covering the length of the Solimões-Amazonas River in Brazil. Genetic diversity parameters suggest there is no genetic differentiation between the five localities. Population’s expansion indicated by R2 and Fu’s Fs tests was also confirmed by the high number of unique haplotypes found. The Analyses of molecular variance indicated that nearly all variability was contained within locations (99.86%), and estimates of gene flow among B. vaillantii were high (FST = 0.0014). These results suggest that Brachyplatystoma vaillantii forms a panmitic population along the Solimões-Amazonas River and, has greater genetic variability than other species of the Brachyplatystoma genus available so far. Although the influence of different tributaries on B. vaillantii migration patterns remains uncertain, a single population in the main channel should be consider in future policies for management of this resource. However, since the species’ life cycle uses habitats in several countries, its management and conservation depend greatly of internationally joined efforts.
Keywords: Control region, Fishery resources, mtDNA, Piramutaba, Population genetics.
O bagre migrador, Brachyplatystoma vaillantii, é um dos mais importantes recursos pesqueiros da Amazônia. Intensa captura ocorre associada ao seu ciclo de vida. Para conhecer seu status genético, sequenciamos a região de controle do DNA mitocondrial de 150 indivíduos, coletados em cinco locais de desembarque pesqueiro, abrangendo toda a extensão do rio Solimões-Amazonas no Brasil. Os parâmetros de diversidade genética sugerem que não existe diferenciação genética entre as cinco localidades amostradas. A expansão populacional indicada pelos testes R2 e Fs de Fu, também foi confirmada pelo elevado número de haplótipos únicos encontrados. A análise de variância molecular indicou que quase toda a variabilidade estava contida nas localidades (99,86%), e as estimativas de fluxo gênico desta espécie eram altas (FST = 0,0014). Esses resultados sugerem que Brachyplatystoma vaillantii forma uma população panmítica ao longo do rio Solimões-Amazonas com maior variabilidade genética que outras espécies do gênero Brachyplatystoma disponíveis no momento. Embora a influência dos diferentes afluentes na migração de B. vaillantii permaneça incerta, em futuras políticas de gestão deste recurso deve-se considerá-lo como uma única população no canal principal. Entretanto, uma vez que seu ciclo de vida abrange habitats em vários países, seu manejo e conservação dependem muito de esforços internacionais em conjunto.
Palavras-chave: DNAmt, Genética populacional, Piramutaba, Recursos pesqueiros, Região controle.
Introduction
The Amazon Basin has features such as altitude, rainfall, temperature, forest cover and water chemistry that affect the diversity of local assemblages (Ríos-Villamizar et al., 2020). Aquatic organisms have not only adapted to these features, but also developed a variety of life cycles as a result (Reis et al., 2016). As a putative consequence of habitat diversity and life strategies, Amazonian ichthyofauna also became extremely diverse, with around 3000 described species (Dagosta, De Pinna, 2019). With the exception of the giant Osteoglossiform Arapaima gigas, the largest Amazonian fishes known belong to the Family Pimelodidae, of the Order Siluriformes (catfishes). In the Amazon the pimelodids, or goliath catfishes, mostly inhabit the main channels, and several species developed seasonal migratory behavior and use the entire length of the Amazon River System to complete their life cycle (Winemiller, 1995, 1989; Ruffino, Isaac, 2000; Klautau et al., 2016).
Brachyplatystoma vaillantii (Valenciennes, 1840) (piramutaba in Portuguese, pirabutón in Spanish, laulao catfish in English), an Amazonian pimelodid, is considered the most important freshwater fish exported from Brazil according to the annual capture records for the species (Frédou et al., 2010; Klautau et al., 2016). The life cycle of B. vaillantii, and other migratory pimelodid catfishes, as currently understood, was originally proposed based upon an analyses of several thousand individual samples obtained from rivers across the Amazon Basin (Barthem, Goulding, 1997). This hypothesis established a migratory pattern, where the headwater tributaries of the Solimões-Amazonas River, near the Andes, were considered the main spawning area. The mouth of the Amazon River, an estuarine area with high primary production, is the nursery area for young fish, while the main channel of the lower Solimões-Amazonas River forms the feeding habitat for juveniles and adults, due to a high biomass of prey fish (Barthem, Goulding, 1997, 2007). Currently, the greatest distances measured between spawning and nursery areas for B. vaillantii are 3,129 km in the Madeira River (Barthem et al., 2017), 4,754 km in Napo River and 4,847 in Marañon River (Maggia et al., 2017), determined by the presence of larvae and juveniles in those areas.
This migratory hypothesis for B. vaillantii has been reassessed by Hegg et al. (2015), who compared Strontium isotope signatures (87Sr/86Sr), and found that the juvenile B. vaillantii had strontium isotope signatures which belong to the Western Andean tributaries and the Amazon River main channel. Analyzes of the link between size classes distribution of larvae, juveniles and adults in the Amazon have also found evidence reassuring the migratory movements of several Amazonian goliath catfishes species, including B. vaillantii. The presence of mature adults in the Marañón and Napo rivers in Peru and Ecuador, respectively, also reinforces the evidence of spawning sites in the far Western Amazon (Barthem et al., 2017). In addition, the presence of B. vaillantii larvae in these two tributaries was also verified from DNA metabarcode analyzes (Maggia et al., 2017).
Although there is considerable evidence corroborating the existence of long migratory events for piramutaba, several details associated to it remain unclear. For instance, there is no information available concerning the number of times a single individual might complete a full migratory cycle, nor to what locations within the Basin the adults may move after spawning. The potential role of the different tributaries of the Amazon in segregating stocks or populations is also unknown (Araujo-Lima, Ruffino, 2003; Córdoba et al., 2011, 2000). In parallel to its migration pattern, or perhaps as a consequence of it, B.vaillantii is caught during the low water season in various Amazonian regions along the entire length of the Solimões-Amazonas River axis, under different fishing pressures. For instance, it is captured by the artisanal fishing fleet in Western Amazon, and predominantly by industrial fishing boats near the estuary at the mouth (Eastern) of the Amazon (Frédou et al., 2010; Klautau et al., 2016). Due to intense levels of fishing over several decades, B. vaillantii is currently considered to be overfished in the Amazon (Alonso, Pirker, 2005; Klautau et al., 2016). However, although the intense level of exploitation, little knowledge exists concerning how this species fully uses its habitat during their life cycle. Identification of the genetic structure of stocks and knowledge regarding the area covered by the species is of key importance for the implementation of effective strategies to manage and maintain this important natural resource (Ruffino et al., 2000). In this context, studies using DNA markers to estimate the genetic variability of species and populations can help identify and better manage fish stocks in the Amazon Basin. Some sequence-based mitochondrial DNA studies have been of key importance in elucidating various biological aspects of commercially important fishes in the Amazon as Colossoma macropomum (Cuvier, 1816) (Farias et al., 2010; Santos et al., 2018, 2007), Zungaro zungaro (Humboldt, 1821) (Boni et al., 2011), Nannostomus eques Steindachner, 1876 (Terencio et al., 2012), Triportheus albus Cope, 1872 (Cooke et al., 2012), Cichla temensis Humboldt, 1821 (Willis et al., 2015), Prochilodus nigricans Spix & Agassiz, 1829 (Machado et al., 2017), Hypophthalmus marginatus Valenciennes, 1840 (Santos et al., 2019), Myloplus spp. (Ota et al., 2020) including also migratory catfishes as Pseudoplatystoma spp. (García-Dávila et al., 2013), Brachyplatystoma rousseauxii (Castelnau, 1855) (Batista, Alves-Gomes, 2006; Batista, 2010), Brachyplatystoma filamentosum (Lichtenstein, 1819), Brachyplatystoma capapretum Lundberg & Akam, 2005 (Huergo et al., 2011), among others.
Accordingly, the current study aims to estimate the genetic variability of B. vaillantii in the Solimões-Amazonas River system (SARS) using mitochondrial DNA (mtDNA) control region sequences. Our main goal is to evaluate whether the species is composed of a single stock (genetically homogeneous), or if the fisheries industry exploit more than one stock (genetically differentiated), in the SARS.
Material and methods
Sampling and data collection. Muscle tissues from 150 Brachyplatystoma vaillantii individuals were collected between 1999 and 2002 near fishing harbors or boat landing sites in 14 localities along the full length of the Solimões-Amazonas System in Brazil, in order to contemplate the main migratory area of piramutaba in the Brazilian Amazon. These 14 landing sites, were further grouped into five locations, which reflected geographical proximity and/or common commercial grounds as follow: Tabatinga (1 –Benjamin Constant and 2 – Tabatinga), Tefé (3 – Mucura Lake, 4 – Tefé, and 5 – Vila Nova), Manaus (6 – Manaus and 7 – Careiro da Várzea), Santarém (8 – Santarém and 9 – Tapará) and Estuary (10 – Almerim, 11 – Gurupá, 12 – Breves, 13 – Belém and 14 –Salvaterra). Each main location is represented by a different color in Fig. 1. All specimens were collected in accordance with Brazilian laws, using a collection license approved by the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA) via permit numbers 130 and 14278–1. Muscle tissue samples were preserved in 95% ethanol in the field, subsequently were conserved at -20 °C and deposited in the Genetic Resources Collection of the National Institute of Amazonian Research (CRG/INPA) (details described in S1).
Laboratory analysis and DNA sequencing. The DNA extraction protocol used was SDS and phenol-chloroform (Sambrook et al., 1989), as described in Alves-Gomes et al. (1995), with minor adaptations. Total DNA concentration was diluted to between 10 and 20 ng/μL. This DNA extract underwent polymerase chain reaction (PCR) with buffer (1X), MgCl2 (2 mM), Taq DNA polymerase (0.02U/μL), dNTPs (0.1 mM), and 0.25 μM of each primer. We designed the forward primer to amplify the control region: CytbPD- L (5 ́ CAC CTG AAT CGG AGG CAT GCC CGT 3 ́), and used the reverse primer DLR1-H (5` GGA TAC TTG CAT GTA TAA ATT GG 3`) (Huergo et al., 2011). Amplification was performed for 30 cycles, with the following temperature profile: the first five cycles were run for 1 min at 94 °C, 1 min at 53 °C and 1.5 min at 72 °C. In the remaining 25 cycles, the annealing temperature was 50 °C. PCR products were purified using a GFX kit (GE Healthcare), following the manufacturer’s protocol. Primers FTTP-L (5` CCA AGC GCC GGT CTT GTA A 3`) (Huergo et al., 2011) and F-12R (5` GTC AGG ACC ATG CCT TTG TG 3`) (Sivasundar etal., 2001) were used as internal primers for sequencing., cycle-sequencing using a DYEnamic ET dye terminator kit (GE Healthcare) under the manufacturers’ protocols and submitted to capillary electrophoresis on a MegaBACE 1000 DNA sequencer (GE Healthcare).

FIGURE 1 | Sampling sites for Brachyplatystoma vaillantii along the Solimões-Amazonas River axis. The localities were grouped in five fishing landing towns as follow: Red circles: Tabatinga (1 – Benjamin Constant, 2 – Tabatinga), Orange circles: Tefé (3 – Mucura Lake, 4 – Tefé, 5 – Vila Nova), Green circles: Manaus (6 – Manaus, 7 – Careiro da Várzea), Purple circles: Santarém (8 – Santarém, 9 – Tapará) and Blues circles: Estuary (10 – Almeirim, 11 – Gurupá, 12 – Breves, 13 – Belém, 14 – Salvaterra).
Data Analysis. The sequence data were checked and compiled in BIOEDIT 7.0.5 (Hall, 1999) and aligned using MUSCLE (Edgar, 2004). Sequences from each of the 14 localities were grouped into their five respective main areas in order to calculate the genetic parameters (see color code in Fig. 1). Genetic variability was estimated by the following parameters: number of haplotypes, number of single haplotypes, polymorphic sites, haplotype diversity (Nei, Tajima, 1981), nucleotide diversity (Nei, 1987) and average pairwise distance estimated via ARLEQUIN 3.5.2 (Excoffier, Lischer, 2010) and DnaSP 6 (Rozas et al., 2017).
Analyses were performed using DnaSP 6 (Rozas et al., 2017) to calculate the R2 and Fu’s FS (Fu, 1997) statistics. These two statistics are considered to be among the most useful for detecting population expansion events in nonrecombining regions of the genome under a variety of different circumstances, especially when population sample sizes are large (Fu’s Fs) or when sample sizes are small (R2) (Ramos-Onsins, Rozas, 2002).
The relationships among haplotypes and their geographic distribution was assessed using statistical parsimony as implemented in TCS 1.21 (Clement et al., 2000), with a 95% connection limit.
To test for the presence of population structure, we estimated the number of clusters (k) using Bayesian posterior probability in BAPS 6.0 (Cheng et al., 2013; Corander et al., 2013). A mixture model was applied to test for groups of individuals using four interactions of k ranging from one to ten. No prior information about sampling locations was provided. In order to verify possible lineages, a Maximum Likelihood (ML) tree was performed, under model HKY+I+G as evaluated in JModelTest 2 (Darriba et al., 2012), using RAXML-NG (Kozlov et al., 2019). The reliability of the ML tree was assessed by bootstrap analysis including 200 replications. The resulting best tree was visualized using FigTree v1.4.4 (Rambaut, 2018). Still using the hierarchical Bayesian clustering algorithm, we used FASTBAPS program (Tonkin-Hill et al., 2019) implemented in R 4.0.3 (R Core Team, 2019) with packages APE, GGPLOT2, GGTREE, MAPS and PHYTOOLS to infer genetic clusters for the data set.
The structure of genetic variation was quantified, in Arlequin 3.5.2, using analysis of molecular variance (AMOVA) with 10,000 permutations (Excoffier et al., 1992), considering haplotype frequency and sequence divergence. AMOVA produces estimates of covariance components and calculates φST values that are analogues of Weir, Cockerham’s FST estimator (Weir, Cockerham, 1984).
To estimate genetic exchange rates between localities, we used the program Migrate-N 4.4.3 (Beerli, Palczewski, 2010), under a bayesian inference coalescent model, and estimated effective population sizes and migration rates (migration/mutation). We used a one long chain, uniform prior distribution, 1,000,000 visited genealogies, 10,000 recorded genealogies with a burn-in of 100,000. In this analysis, the migration rates were only inferred between adjacent, hydrographically connected localities (forcing migration between non-adjacent groups to first occur through geographically intermediate localities).
Results
For the 150 samples of Brachyplatystoma vaillantii, the mitochondrial DNA control region varied from 936 to 941 base pairs (bp). To align, one to six gaps in the sequences were added, representing insertion and exclusion events (indels), resulting in a final matrix with 942 bp. Within the final data matrix, 109 sites were variable, once indels were considered a fifth character state and, among those, we observed indels at 10 sites.
TABLE 1 | Summary of genetic parameters estimated from the Brachyplatystoma vaillantii mtDNA control region for five localities on the Solimões-Amazonas River. N = number of samples, H = number of haplotypes, uH = number of unique haplotypes, S = number of polymorphic sites, ETA = Total Number of Mutations sites, HD = haplotype diversity, π = nucleotide diversity, k = average number of nucleotide differences, * = p<0.02.
Localities | N | H | uH | S | ETA | HD | π | k | R2 | Fu’s Fs |
Estuary | 30 | 30 | 27 | 63 | 63 | 1.000±0.008 | 0.014±0.007 | 13.669±6.316 | 0.096 | -20.12* |
Santarém | 30 | 26 | 19 | 66 | 67 | 0.991±0.011 | 0.016±0.008 | 15.183±6.981 | 0.105 | -8.59* |
Manaus | 30 | 29 | 25 | 61 | 61 | 0.997±0.009 | 0.015±0.007 | 14.602±6.725 | 0.101 | -15.81* |
Tefé | 30 | 29 | 27 | 60 | 60 | 0.997±0.009 | 0.014±0.007 | 13.579±6.370 | 0.096 | -16.52* |
Tabatinga | 30 | 30 | 27 | 64 | 64 | 1.000±0.008 | 0.015±0.007 | 14.664±6.753 | 0.103 | -19.17* |
All | 150 | 134 | 125 | 109 | 111 | 0.997±0.001 | 0.015±0.008 | 14.399±6.487 | 0.059 | -23.98* |
Haplotype diversity (HD) at individual localities varied from 0.991 at Santarém to 1.000 at Estuary and Tabatinga. Nucleotide diversity ranged from 0.014 in Estuary and Tefé to 0.016 in Santarém. Results are summarized in Tab. 1.
The negative and significant Fs values (P <0.02; Tab. 1) for the Fu test indicate that the species is undergoing population expansion at all sampled locations. The low R2 obtained for the sampled locations corroborate the FS values of the Fu statistical tests, suggesting that B. vaillantii has gone through a population expansion. Furthermore, since R2 statistics is based on the difference between the number of singleton mutations and the average number of nucleotide differences, the high number of singletons seen in B. vaillantii were also taken as evidence of a population expansion.
We identified 134 haplotypes in the control region sequences (details described in S1). Most haplotypes (125) were unique and there were only nine shared haplotypes among the different localities. Network’s topology showed a random distribution of haplotypes across the five localities without association to geographic location (Fig. 2).
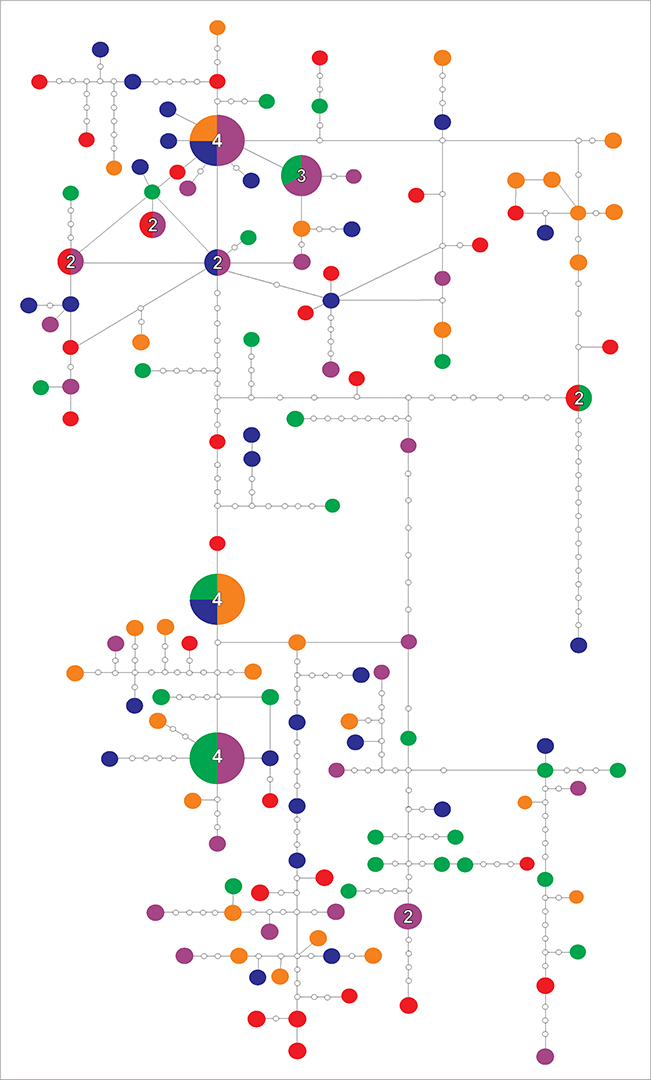
FIGURE 2 | Network of Brachyplatystoma vaillantii haplotypes. The number shown within a circle identifies the number of specimens sharing those haplotypes; circles without numbers represent unique haplotypes. White circles represent hypothetical intermediate haplotypes. Each locality is represented by the same colors in Fig. 1: red – Tabatinga, orange – Tefé, green – Manaus, purple – Santarém and blue – Estuary.
BAPS analysis defined three clusters (K = 3) distributed over the sampled localities (Fig. 3) showing no geographic segregation. The final ML tree recovered topology presenting weak support within clades (details described in S2). The hierarchical Bayesian clustering generated by FASTBAPS program also could not recover high bootstrap support (Fig. 4).

FIGURE 3 | Analysis of the BAPS 6.0 program showing three clusters (green, red, and blue) distributed between the five sampled locations of Brachyplatystoma vaillantii.

FIGURE 4 | Analysis of the FASTBAPS program. Numbers are individual sequence of Brachyplatystoma vaillantii. Colors ranging from red for the lowest probabilities and clear yellow for the highest probabilities support for bootstrap.
The AMOVA results showed that most genetic variation is within sampled localities (99.86%) and no genetic differentiation was identified between the localities (FST = 0.0014). Estimates of population divergence with pairwise FST values revealed that all localities were not significantly divergent from each other (Tab. 2). A high gene flows levels between adjacent localities according to the migration rate values (M) obtained with Migrate-N program (Tab. 3).
Discussion
Our study of Brachyplatystoma vaillantii genetic variability offers the most comprehensive genetic evaluation for a commercially important resource with a wide distribution across the Amazon Basin. Molecular data became a powerful tool for inferring populational structure, or the lack thereof. Using similar sample sizes, studies of different Neotropical fishes have be able to detect phylogeographic structure in species such as Triportheus albus(Cooke et al., 2012) and Nannostomus eques (Terencio et al., 2012). Further, it has been possible to detect genetic structuring in migratory and abundant freshwater fish species such as in the genus Prochilodus Agassiz, 1829 (Sivasundar et al., 2001) as well as in Arapaima gigas (Shinz, 1822)(Hrbek et al., 2005),a fish which also occurs over semi-continental geographic distances.
All estimated parameters associated with the B. vaillantii genetic diversity suggest that there is no genetic differentiation between the five localities sampled (Tab. 1).
TABLE 2 | Pairwise FST values between the Brachyplatystoma vaillantii individuals from the localities sampled.
Localities | Estuary | Santarém | Manaus | Tefé |
Estuary | – |
|
|
|
Santarém | 0.0000 | – |
|
|
Manaus | 0.0109 | 0.0000 | – |
|
Tefé | 0.0000 | 0.0000 | 0.0168 | – |
Tabatinga | 0.0000 | 0.0000 | 0.0285 | 0.0016 |
TABLE 3 | Estimates of effective number immigrating individuals per generation for Brachyplatystoma vaillantii. Source population are in the row, recipient population in the columns. The signal “+” represents receiving localities. The signal “–” represents without comparison. Migration was defined between adjacent localities.
Localities | Estuary + | Santarém + | Manaus + | Tefé + | Tabatinga + |
Estuary | – | 123.3 | – | – | – |
Santarém | 923.4 | – | 52.1 | – | – |
Manaus | – | 955.9 | – | 935.7 | – |
Tefé | – | – | 251.9 | – | 932.6 |
Tabatinga | – | – | – | 76.2 | – |
The population’s expansion indicated by the values of R2 and Fu’s Fs, can be also confirmed by observation of the number of unique haplotypes, which is either equal or close to the number of individuals collected. Although BAPS program analysis found three clusters, they have representatives from all five locations without geographic segregation. The generated ML tree did not have bootstrap support and presented a large amount of polytomies among analyzed individuals (S2). In the same way, the clades generated by the analysis performed in FASTBAPS program, presented low support values for bootstrap (Fig. 4). The three clusters are present in all the sampled locations along the Solimões-Amazonas River suggests high gene flow. The AMOVA results confirms this hypothesis showing 99.86% of genetic variation occurs within sampled localities, also indicating that specimens form a single genetic stock. Other studies of Amazonian fishes have found similar patterns and DNA polymorphism in mtDNA control region such as Brachyplatystoma rousseauxii (Batista, 2010; Batista, Alves-Gomes, 2006), Serrasalmus rhombeus (Linnaeus, 1766) (Hubert et al., 2007) and Colossoma macropomum (Santos et al., 2007).
In a similar study with C. macropomum (tambaqui), the mtDNA control region of 48 individuals collected in four locations in the Amazon Basin (Tabatinga, Parintins Oriximiná and Santarém), found 31 haplotypes, demonstrating high levels of genetic diversity for all locations sampled (Santos et al., 2007). This pattern can also be observed in another study (Santos et al., 2018) of the same species with larger sampling numbers along the Solimões/Amazonas River (Mexiana, Almeirim, Santarém, Manaus, Tefé and Tabatinga) including sequences of mtDNA control region and ATPase 6/8 concatenated gene. Likewise, there was very little clustering among the haplotypes found at most localities, resulting in haplotype diversity varying between 0.986 and 1,000. The haplotype network also showed numerous reticulations between the Colossoma Eigenmann & Kennedy, 1903 haplotypes implying a high degree of gene exchange. The data suggests that, as C. macropomum migrates over a much smaller geographic area than B. vaillantii, the relatively high rate of gene flow between locations also occurs in this species.
High genetic variability was also found in mtDNA control region of curimatã (Prochilodus nigricans) from the Solimões-Amazonas, Madeira, Purus and Juruá rivers (Machado et al., 2017). As in B. vallantii, P. nigricans in the Solimões-Amazonas Systemis genetically homogeneus and, albeit such high polymorphism have occurred mostly within populations, it was found that the upper Madeira and the Purus rivers may have restricted genetic flow.
For both C. macropomum and P. nigricans there are different scenarios of gene flow between the Solimões-Amazonas River and its tributaries. The genetic diversity of B. vallantii in the distinct tributaries is unknown so far, and this information is crucial for its conservation, since the species reproduction occurs in these areas.
Another example in the Amazon, the zebra shovelnose Brachyplatystoma tigrinum (Britski, 1981) is a commercially important catfish for the ornamental fish trade, especially in Peru. Although the species is listed among those fish whose export or capture from natural populations is prohibited, commercial pressure on this species continues to increase in this country. This is of concern since molecular data showed that this pimelodidae has a genetic variability much reduced in relation to B. vaillantii (Mori-Marin et al., 2012). The haplotype diversity is a clear example of this fact, since this index observed in B. tigrinum (0.527) is practically half of the value for B. vaillantii (0.997)and this value may, in the near future, compromise the recovery of genetic diversity in view of the continuous decrease in the population size of the species.
For the two pimelodid species commonly known as piraíba (B. filamentosum and B. capapretum), haplotype diversity (HD) values are also high (>0.95), and the nucleotide diversity (π) of B. filamentosum (0.0118) (Huergo et al., 2011) is closer to the value found in B. vaillantii (0.015) (details described in S3). However, the nucleotide diversity found for B. capapretum (0.0041) (Lira-Cordeiro, 2013) is only about between 1/3 and ¼ of that found in the previously mentioned species, respectively. Like in B. vaillantii, both piraíbas, also appear to have being overfished (Petrere et al., 2004), but unfortunately no study about their fisheries, up to date, has considered the piraíba as two distinct species (B. filamentosum and B. capapretum), and the current conservational status of each species remains unknown.
Another congener, the catfish known regionally as babão, Brachyplatystoma platynemum Boulenger, 1898, also has high levels of haplotype diversity, although two structured populations from the Amazon and Madeira rivers have been suggested (Ochoa et al., 2015). If we compare B. platynemum only in the main channel of the Solimões-Amazonas River with the data of B. vaillantii from our study, there is a single population in both species, but the haplotypesdiversity of B. vaillantii (0.997) is slightly greater than B. platynemum (0.905). Influence of different tributaries on piramutaba genetic diversity composition remains unknow and this information is important to understand the real and complete its life cycle.
The genetic parameters and the DNA polymorphism analysis indicate that levels of genetic variability for B. vaillantii are high and homogeneous along the main channel of the SARS. This contrasts the data for dourada B. rousseauxii, which shows a significant decrease in genetic diversity towards the Western part of the Amazon (Batista, Alves-Gomes, 2006). A possible genetic segregation in B. vaillantii associated to the different tributaries or to a partial homing behavior, as already suggested for B. rousseauxii (Batista, Alves-Gomes, 2006), shall not be discarded until further data, collected in the headwaters of the Solimões system, be analyzed.
To evaluate gene flow levels between localities we initially considered a simple model of estimation of pairwise FST, which revealed absence of divergence. These results already indicate that B. vaillantii in the Solimões-Amazonas River behaves as a large and unique population. The hypothesis pattern is also confirmed by Migrate-N analysis results, which was similar to the pattern in the FST matrix, implying very high levels of gene exchange between localities.However, as this model essentially determine if a gene lineage is present in any given locality because of mutation in situ or immigration from another locality, in datasets with high genetic diversity, it can be difficult to distinguish which processes resulted in the observed patterns. Moreover, these estimates are based upon a single locus only, and only for estimates of female immigration. These facts likely contain unknown biases that can only be discovered with additional, multi-locus data.
Although we have information from a single locus, our data appear to be sufficiently robust pointing that B. vaillantii along the Solimões-Amazonas River is composed by one, large and panmitic population, with a genetically diverse and geographically homogeneous organization. In consequence, we suggest that, for all management purposes, that B. vaillantii should be considered as a single population along the main Amazon-Solimões River axis.
In general, the results for B. vaillantii are similar to those found in analyses of the control region from other Amazonian species, although its values are consistently higher. In fact, among the species of genus Brachyplatystoma so far tested, B. vailantii shows the highest genetic variability in the Solimões-Amazonas River (details described in S4 and S3) (Batista, Alves-Gomes, 2006; Batista, 2010; Huergo et al., 2011; Mori-Marin et al., 2012; Lira-Cordeiro, 2013; Ochoa et al., 2015).
Brachyplatystoma rousseauxii and B. vaillantii are believed to have similar life cycles, and the fishing efforts over both resources have evolved taking into account their migration routes and seasonality. The geographic extent of the routes used by these two species includes the whole Solimões-Amazonas River and their headwater tributaries. Basically, these two species use the entire Amazon from the estuary next to the Atlantic Ocean to the headwater’s streams near the Andes in order to complete their life cycle (Barthem, Goulding, 1997). However, some studies and local reports suggest that there are migratory differences between B. rousseauxii and B. vaillantii. The former do not form large schools; they are found near the surface or mid-water year-round along the SARS and their frequency increases during the dry season (Telles et al., 2011; Carvajal-Vallejos et al., 2014; Barthem et al., 2017). On the other hand, B. vaillantii is a river-bottom swimmer, but can explore the entire water column in search of prey, probably preferring preys that inhabits the range of 5–10 m of depth (Barthem, Petrere Jr, 1995; Barthem, Goulding, 1997). Brachyplatystoma vaillantii tends to migrate in large schools, but they are found in large numbers mainly during the low water season (July to December) (Chaves et al., 2003; Barthem, Goulding, 2007; Leite et al., 2007; Maggia et al., 2017; De Pinna, 2019).
It is well established that B. rousseauxii’s maximum length (192 cm) is larger than that of B. vaillantii (105 cm)(Barthem, Goulding, 1997). On the other hand, B. vaillantii reaches the first sexual maturation at an earlier age (2 years) than B. rousseauxii (3 years) (Alonso, Pirker, 2005). Differences in genetic diversity between B. rousseauxii (π = 0.0079, Haplotype diverstity = 0.933 S (Polymorphic sites) = 54) and B. vaillantii (π = 0.0150, Haplotype diverstity = 0.997 S = 133) may be reflecting peculiarities inherent to each species, and are very probably associated with subtle differences in their life cycles. Based on coalescence theory Batista et al. (2004) estimated the evolutionary age, using Watterson’s population genetics estimate, suggesting a relative age of approximately 242,600 and 335,200 years for B. rousseauxii and B. vaillantii, respectively. If the mutation rates were equal for both species, this difference in evolutionary age could represent more time for B. vaillantii to accumulate mutations, resulting, consequently, in higher levels of genetic variability reflected in nucleotide and haplotype diversity. Nevertheless, other factors may also be involved in different genetic variability of these species, such as the time of sexual maturation, the number of offspring produced per life span, and potentially different intrinsic mutational rates.
Despite being overfished, B. vaillantii is still genetically more diverse than any other commercially important species within the genus Brachyplatystoma studied so far: dourada (B. rousseauxii), piraíba (B. filamentosum), filhote capapreta (B. capapretum), babão (B. platynemum) and dourada zebra (B. tigrinum). Such situation suggests that either the B. vaillantii possesses an intrinsic mechanism, such as, for example, a mutation rate that is faster than that of other conspecific species or, alternatively, the other species are even more highly overfished and their stock are even more depleted genetically than B. vaillantii. Such a scenario requires serious consideration by those tasked with effective long-term fish-stock management in the Amazon, especially considering that these migratory resources overpass political borders and require joined efforts and policies from the Amazonian countries.
The intense exploitation of B. vaillantii in the estuary, performed mainly by industrial fleet, should also be considered a critical issue that needs to be addressed urgently. The industrial fleet mostly catches small (20–30 cm) and medium-sized (30–50 cm) specimens. In an attempt to minimize this impact, exclusion areas for the industrial fleet were implemented, but there are no studies or guarantees demonstrating that those defined areas actually correspond to habitats naturally occupied by the species (Barthem et al., 2015).
As with several other migratory species in the Amazon, the current situation of B. vaillantii stock requires a multidisciplinary approach and a complex case evaluation. The several challenging issues to be considered include, but are not limited to: the increasing demand for fish on the national and international market; the increasing capture effort of the fishing fleet, as well as their numerical growth in the Amazon, without any compensatory policies to increase the stocks; the lack of control over the size and amount of fish captured; a lack of detailed information about the life cycle of the resource; the absence of fishery statistics in the entire Amazon; the uncontrolled land use and forest clearing in several basins and sub-basins in the Amazon resulting in habitat deterioration, especially those areas of spawning; the construction of large dams for electricity generation in key rivers, which threatens migration routes and access to spawning habitats and; the fact that these fish do not recognize political borders and require coordinated international planning and action (Klautau et al., 2016; Forsberg et al., 2017; Silva-Júnior et al., 2017).
In summary, the sustainability of these fish resources/population and the economical chain they support, especially across international borders, continue to depend on accurate population assessments, including their genetic parameters, and detailed knowledge of their behavior and ecology (Pelicice et al., 2015; Barthem et al., 2017).
Any discussion of a fishery resource management must include, especially in the Amazon, at least two contexts: the complex commercial and social interactions associated with the fishing activity itself, and the ecological interconnections that characterize the species under consideration. For a migratory fish such as B. vaillantii, both fishing activity and the ecology of the species are subject to changes from one region to another, since the fish only spawn in the headwaters of the Amazon River’s affluents, but grow up in the mouth of the Amazon River. Thus, in order to construct a series of policies that will provide full and effective management and conservation of this resource, a well-coordinated series of actions among the countries involved is mandatory.
Acknowledgments
This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq/National Genome Project (#480183/00–7 awarded to JAAG) IBAMA/PROVÁRZEA awarded to JAAG, International Foundation for Science awarded to JAAG, an MCTI-INPA (grant PPI #1–3550 awarded to JAAG), FAPEAM/SEDECTI/Governo do Estado do Amazonas – Edital PAPAC 005/2019 to JSB/PPG GCBEv and Bolsas Funbio – Conservando o Futuro Program to KMF. The authors thank Dr. Eduardo Antonio Ríos-Villamizar (INPA), Dr. Romina Batista (INPA) and MsC. Erico Polo (UFAM) for suggestions on the manuscript.
References
Alonso JC, Pirker LEM. Dinâmica populacional e estado atual da exploração de piramutaba e de dourada. In: Fabré NN, Barthem RB, organizers. O manejo da pesca dos grandes bagres migradores: piramutaba e dourada no eixo Solimões-Amazonas. Manaus: IBAMA/Pro Várzea, 2005. p.21–28.
Alves-Gomes JA, Orti G, Haygood M, Heiligenberg W, Meyer A. Phylogenetic analysis of the South American electric fishes (Order Gymnotiformes) and the evolution of their electrogenic system: a synthesis based on morphology, electrophysiology, and mitochondrial sequence data. Mol Biol Evol. 1995; 12(2):298–318. https://doi.org/10.1093/oxfordjournals.molbev.a040204
Araujo-Lima CRM, Ruffino ML. Migratory fishes of the Brazilian Amazon. In: Carolsfeld J, Harvey B, Ross C, Baer A, editors. Migratory fishes of South America: biology, fisheries, and conservation status. Washington DC: The International Bank for Reconstruction and Development / The World Bank; 2003. p.233–301.
Barthem RB, Goulding M. Um ecossistema inesperado: A Amazônia revelada pela pesca. Lima: Gráfica Biblos; 2007.
Barthem RB, Goulding M. The Catfish Connection: Ecology, Migration, and Conservation of Amazon Predators. New York: Columbia University Press; 1997.
Barthem RB, Goulding M, Leite RG, Cañas C, Forsberg B, Venticinque E et al. Goliath catfish spawning in the far western Amazon confirmed by the distribution of mature adults, drifting larvae and migrating juveniles. Sci Rep. 2017; 7:41784. https://doi.org/10.1038/srep41784
Barthem RB, Mello Filho A, Assuncao W, Fazzi Gomes PF, Campos Barbosa CA. Estrutura de tamanho e distribuição espacial da piramutaba (Brachyplatystoma vaillantii) na foz amazônica: implicações para o Manejo da pesca. Bol Inst Pesca. 2015; 41(2):249–60. Available from: https://www.pesca.sp.gov.br/41_2_249-260.pdf
Barthem RB, Petrere Jr M. Fisheries and population dynamics of Brachyplatystoma vailantii (Pimelodidae) in the Amazon Estuary. In: Armantrout NB, Wolotira RJ, editors. Cond World’s Aquat habitat Proc World Fish Condition of the world’s aquatic habitats: proceedings of the World Fisheries Congress, Vol. 1. New Delhi: Oxford & IBH Pub. Co.; 1995. p.329–40.
Batista JS. Caracterização genética da dourada-Brachyplatystoma rousseauxii (Siluriformes: Pimelodidae), na Amazônia por meio de marcadores moleculares mitocondriais e microssatélites: subsídios para conservação e manejo. [PhD Thesis]. Manaus: Instituto Nacional de Pesquisas da Amazônia; 2010. Available from: https://koha.inpa.gov.br/cgi-bin/koha/opac-detail.pl?biblionumber=5388
Batista JS, Alves-Gomes JA. Phylogeography of Brachyplatystoma rousseauxii (Siluriformes – Pimelodidae) in the Amazon Basin offers preliminary evidence for the first case of “homing” for an Amazonian migratory catfish. Genet Mol Res. 2006; 5(4):723–40. Available from: https://www.geneticsmr.com/sites/default/files/articles/year2006/vol5-4/pdf/gmr0231.pdf
Batista JS, Formiga-Aquino K, Farias IP, Alves-Gomes JA. Genetic variability studies of piramutaba – Brachyplatystoma vaillantii and dourada – B. rousseauxii (Pimelodidae: Siluriformes) in the Amazon: basis for management and conservation. Extended abstract submitted to the VI International Congress on the Biology of Fish; 2004. p.253–58.
Beerli P, Palczewski M. Unified framework to evaluate panmixia and migration direction among multiple sampling locations. Genetics. 2010; 185(1):313–26. https://doi.org/10.1534/genetics.109.112532
Boni TA, Padial AA, Prioli SMAP, Lucio LC, Maniglia TC, Bignotto TS et al. Molecular differentiation of species of the genus Zungaro (Siluriformes, Pimelodidae) from the Amazon and Paraná-Paraguay River basins in Brazil. Genet Mol Res. 2011; 10(4):2795–805. https://doi.org/10.4238/2011.November.10.2
Carvajal-Vallejos FM, Duponchelle F, Desmarais E, Cerqueira F, Querouil S, Nuñez J et al. Genetic structure in the Amazonian catfish Brachyplatystoma rousseauxii: Influence of life history strategies. Genetica. 2014; 142:323–36. https://doi.org/10.1007/s10709-014-9777-2
Chaves RA, Silva KCA, Ivo CTC, Cintra IHA, Aviz JS. Sobre a pesca da piramutaba, Brachyplatystoma vaillantii (Valenciennes, 1940) em pescarias da frota industrial no estado do Pará. Boletim Técnico Científico CEPNOR. 2003; 3(1):163–77.
Cheng L, Connor TR, Sirén J, Aanensen DM, Corander J. Hierarchical and spatially explicit clustering of DNA sequences with BAPS software. Mol Biol Evol. 2013; 30(5):1224–28. https://doi.org/10.1093/molbev/mst028
Clement M, Posada D, Crandall KA. TCS: a computer program to estimate gene genealogies. Mol Ecol. 2000; 9(10):1657–59. https://doi.org/10.1046/j.1365-294x.2000.01020.x
Cooke GM, Chao NL, Beheregaray LB. Divergent natural selection with gene flow along major environmental gradients in Amazonia: Insights from genome scans, population genetics and phylogeography of the characin fish Triportheus albus. Mol Ecol. 2012; 21(10):2410–27. https://doi.org/10.1111/j.1365-294X.2012.05540.x
Corander J, Lu Cheng L, Marttinen P, Tang J. BAPS : Bayesian Analysis of Population Structure. Manual v 6.0. Bioinformatics. 2013:1–28.
Córdoba EA, Coy YS, Páez CLS, Sosa DLM, González JCA, Díaz MEA et al. Bagres de la Amazonia Colombiana: Um recurso sin fronteiras. Bogotá: Instituto Amazónico de Investigaciones Científicas SINCHI; 2000.
Córdoba EA, Páez CLS, Fernández CAR, Bonilla-Castillo CA, Hurtado GAG. Diagnóstico de la pesquería en la cuenca del Amazonas. In: Lasso CA, Paula-Gutiérrez F, Morales-Betancourt MA, Agudelo E, Ramírez-Gil H, Ajiaco-Martínez RE, editors. Pesquerías continentales de Colombia: cuencas del Magdalena Cauca, Sinú, Canalete, Atrato, Orinoco, Amazonas y vertiente del Pacífico. Bogotá: Instituto de Investigación de los Recursos Biológicos Alexander von Humboldt; 2011. p.143–67.
Dagosta FCP, De Pinna M. The Fishes of the Amazon: Distribution and biogeographical patterns, with a comprehensive list of species. Bull Am Museum Nat Hist. 2019; (431):1–163. https://doi.org/10.1206/0003-0090.431.1.1
Darriba D, Taboada GL, Doallo R, Posada D. JModelTest 2: More models, new heuristics and parallel computing. Nat Methods. 2012; 9(8):772. https://doi.org/10.1038/nmeth.2109
Edgar RC. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics. 2004; 5:1–19. https://doi.org/10.1186/1471-2105-5-113
Excoffier L, Lischer HEL. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour. 2010; 10(3):564–67. https://doi.org/10.1111/j.1755-0998.2010.02847.x
Excoffier L, Smouse PE, Quattro JM. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics. 1992; 131:479–91.
Farias IP, Torrico JP, García-Dávila C, Santos M da CF, Hrbek T, Renno JF. Are rapids a barrier for floodplain fishes of the Amazon basin? A demographic study of the keystone floodplain species Colossoma macropomum (Teleostei: Characiformes). Mol Phylogenet Evol. 2010; 56(3):1129–35. https://doi.org/10.1016/j.ympev.2010.03.028
Forsberg BR, Melack JM, Dunne T, Barthem RB, Goulding M, Paiva RCD et al. The potential impact of new Andean dams on Amazon fluvial ecosystems. PLoS One. 2017; 12(8):e0182254. https://doi.org/10.1371/journal.pone.0182254
Frédou FL, Almeida OT, Rivero S, Mourão K, Barbosa C, Thompson R. Aspectos econômicos da pesca industrial do Pará: entraves e perspectivas. Papers do NAEA. 2010; (265):1–25.
Fu YX. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics. 1997; 147(2):915–25. Available from: https://www.genetics.org/content/genetics/147/2/915.full.pdf
García-Dávila C, Duponchelle F, Castro-Ruiz D, Villacorta J, Quérouil S, Chota-Macuyama W et al. Molecular identification of a cryptic species in the Amazonian predatory catfish genus Pseudoplatystoma (Bleeker, 1962) from Peru. Genetica. 2013; 141(7–9):347–58. https://doi.org/10.1007/s10709-013-9734-5
Hall T. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. 1999; 41:95–98.
Hegg JC, Giarrizzo T, Kennedy BP. Diverse early life-history strategies in migratory Amazonian catfish: Implications for conservation and management. PLoS One. 2015; 10(7):1–19. https://doi.org/10.1371/journal.pone.0129697
Hrbek T, Farias IP, Crossa M, Sampaio I, Porto JIR, Meyer A. Population genetic analysis of Arapaima gigas, one of the largest freshwater fishes of the Amazon basin: Implications for its conservation. Anim Conserv. 2005; 8(3):297–308. https://doi.org/10.1017/S1367943005002210
Hubert N, Duponchelle F, Nuñez J, Rivera R, Bonhomme F, Renno JF. Isolation by distance and Pleistocene expansion of the lowland populations of the white piranha Serrasalmus rhombeus. Mol Ecol. 2007; 16(12):2488–503. https://doi.org/10.1111/j.1365-294X.2007.03338.x
Huergo GM, Filgueiras-Souza RJ, Batista JS, Formiga-Aquino K, Alves-Gomes JA. Molecular genetics as a tool for fisheries management in the Brazilian Amazon: Piraíba (Brachyplatystoma filamentosum and Brachyplatystoma capapretum) (Siluriformes: Pimelodidae) in white-water rivers. Panam J Aquat Sci. 2011; 6(4):280–9. Available from: https://panamjas.org/pdf_artigos/PANAMJAS_6(4)_280-289.pdf
Klautau AGCDM, Cordeiro APB, Cintra IHA, Silva LEO, Bastos CEMC, Carvalho HRL, Ito LS. Analysis of the industrial fishing of piramutaba catfish, Brachyplatystoma vaillantii (Valenciennes 1840), in two estuarine areas of the brazilian Amazon. Panam J Aquat Sci. 2016; 11(2):143–50. Available from: https://panamjas.org/pdf_artigos/PANAMJAS_11(2)_143-150.pdf
Kozlov AM, Darriba D, Flouri T, Morel B, Stamatakis A. RAxML-NG: a fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics. 2019; 35(21):4453–5. https://doi.org/10.1093/bioinformatics/btz305
Leite RG, Cañas C, Forsberg B, Barthem RB, Goulding M. Larvas dos grandes bagres migradores. Manaus, Lima: Amazônia, Asociación para la Conservación de la Cuenca Amazónica; 2007.
Lira-Cordeiro A. Estimativa da variabilidade genética da piraíba negra Brachyplatystoma capapretum (Siluriformes: Pimelodidae), por meio de marcadores mitocondriais e microssatélites. [Master dissertation]. Manaus: Universidade Federal do Amazonas; 2013.
Machado VN, Willis SC, Teixeira AS, Hrbek T, Farias IP. Population genetic structure of the Amazonian black flannelmouth characin (Characiformes, Prochilodontidae: Prochilodus nigricans Spix and Agassiz, 1829): contemporary and historical gene flow of a migratory and abundant fishery species. Environ Biol Fishes. 2017; 100:1–16. https://doi.org/10.1007/s10641-016-0547-0
Maggia ME, Vigouroux Y, Renno JF, Duponchelle F, Desmarais E, Nunez J et al. DNA metabarcoding of Amazonian ichthyoplankton swarms. PLoS One. 2017; 12(1):e0170009. https://doi.org/doi:10.1371/journal.pone.0170009
Mori-Marin J, Renno JF, Carvajal-Vallejos F, Alcantara-Bocanera F, Duponchelle F, García-Davila CR. Estudio comparativo de la variabilidad genética del zúngaro tigrinus – Brachyplatystoma tigrinum (Britski, 1981) y dos especies relacionadas de la familia pimelodidae. Folia Amazônica. 2012; 21(1–2):87–95. https://doi.org/10.24841/fa.v21i1-2.36
Nei M. Molecular Evolutionary Genetics. New York: Columbia University Press; 1987. https://doi.org/10.7312/nei-92038
Nei M, Tajima F. DNA polymorphism detectable by restriction endonucleases. Genetics. 1981; 97(1):145–63.
Ochoa LE, Pereira LHG, Costa-Silva GJ, Roxo FF, Batista JS, Formiga K et al. Genetic structure and historical diversification of catfish Brachyplatystoma platynemum (Siluriformes: Pimelodidae) in the Amazon basin with implications for its conservation. Ecol Evol. 2015; 5(10):2005–20. https://doi.org/10.1002/ece3.1486
Ota RP, Nogueira Machado V, Andrade MC, Collins RA, Farias IP, Hrbek T et al. Integrative taxonomy reveals a new species of pacu (Characiformes: Serrasalmidae: Myloplus) from the Brazilian Amazon. Neotrop Ichthyol. 2020; 18(1):e190112. https://doi.org/10.1590/1982-0224-20190112
Pelicice Júnior FM, Pompeu PS, Agostinho AA. Large reservoirs as ecological barriers to downstream movements of Neotropical migratory fish. Fish Fish. 2015; 16(4):697–715. https://doi.org/10.1111/faf.12089
Petrere M, Barthem RB, Cordoba EA, Gomez BC. Review of the large catfish fisheries in the upper Amazon and the stock depletion of piraíba (Brachyplatystoma filamentosum Lichtenstein). Rev Fish Biol Fish. 2004; 14:403–14. https://doi.org/10.1007/s11160-004-8362-7
R Development Core Team. R: A language and environment for statistical computing. [Computer software manual – Internet]. R Foundation for Statistical Computing; 2019. Available from: https://www.rproject.org
Rambaut A. FigTree: tree figure drawing tool, version 1.4.4. 2018. http://tree.bio.ed.ac.uk/software/figtree
Ramos-Onsins SE, Rozas J. Statistical properties of new neutrality tests against population growth. Mol Biol Evol. 2002; 19(12):2092–100. https://doi.org/10.1093/oxfordjournals.molbev.a004034
Reis RE, Albert JS, Di Dario F, Mincarone MM, Petry P, Rocha LA. Fish biodiversity and conservation in South America. J Fish Biol. 2016; 89(1):12–47. https://doi.org/10.1111/jfb.13016
Ríos-Villamizar EA, Adeney JM, Junk WJ, Piedade MTF. Physicochemical features of Amazonian water typologies for water resources management. IOP Conference Series, Earth and Environ Scince. 2020; 427:012003. https://doi.org/10.1088/1755-1315/427/1/012003
Rozas J, Ferrer-Mata A, Sánchez-DelBarrio JC, Guirao-Rico S, Librado P, Ramos-Onsins SE, Sánchez-Gracia A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol Biol Evol. 2017; 34(12):3299–302. https://doi.org/10.1093/molbev/msx248
Ruffino ML, Barthem RB, Fischer CFA. Perspectivas do manejo dos bagres migradores na Amazônia. In: Fisher CFA, editor. Recursos pesqueiros do médio Amazonas: biologia e estatística pesqueira. Brasília, DF, Brazil: Coleção Meio Ambiente; 2000. p.141–52.
Ruffino ML, Isaac VJ. Ciclo de vida e parâmetros biólogicos de algumas espécies de peixes da Amazônia brasileira. In: Fisher CFA, editor. Recursos pesqueiros do médio Amazonas: biologia e estatística pesqueira. Brasília, DF, Brazil: Coleção Meio Ambiente; 2000. p.11–30.
Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. vol. 1. 2nd editio. New York: Cold Springs Harbor Laboratory Press; 1989.
Santos JCM, Ferreira ES, Oliveira C, Oliveira TB, Costa ASL, de Queiroz AKO et al. Phylogeny of the genus Hypophthalmus Cuvier, 1829 (Pimelodidae – Siluriformes), based on a multilocus analysis, indicates diversification and introgression in the Amazon basin. Mol Phylogenet Evol. 2019; 137:285–92. https://doi.org/10.1016/j.ympev.2019.05.017
Santos MCF, Hrbek T, Farias IP. A multilocus approach to understanding historical and contemporary demography of the keystone floodplain species Colossoma macropomum (Teleostei: Characiformes). Front Genet. 2018; 263. https://doi.org/10.3389/fgene.2018.00263
Santos MCF, Ruffino ML, Farias IP. High levels of genetic variability and panmixia of the tambaqui Colossoma macropomum (Cuvier, 1816) in the main channel of the Amazon River. J Fish Biol. 2007; 71(SUPPL. A):33–44. https://doi.org/10.1111/j.1095-8649.2007.01514.x
Silva-Júnior UL, Raseira MB, Ruffino ML, Batista V da S, Leite RG. Estimativas do tamanho do estoque de algumas espécies de peixes comerciais da Amazônia a partir de dados de captura e esforço. Biodiversidade Brasileira. 2017; 7(1):105–21.
Sivasundar A, Bermingham E, Ortí G. Population structure and biogeography of migratory freshwater fishes (Prochilodus: Characiformes)in major South American rivers. Mol Ecol. 2001; 10(2):407–17. https://doi.org/10.1046/j.1365-294X.2001.01194.x
Telles MPC, Collevatti RG, Costa MC, Barthem RB, Silva Júnior NJ, Neto ACS et al. A geographical genetics framework for inferring homing reproductive behavior in fishes. Genetica. 2011; 139(2):243–53. https://doi.org/10.1007/s10709-010-9541-1
Terencio ML, Schneider CH, Porto JIR. Molecular signature of the D-loop in the brown pencilfish Nannostomus eques (Characiformes, Lebiasinidae) reveals at least two evolutionary units in the Rio Negro basin, Brazil. J Fish Biol. 2012; 81(1):110–24. https://doi.org/10.1111/j.1095-8649.2012.03320.x
Tonkin-Hill G, Lees JA, Bentley SD, Frost SDW, Corander J. Fast hierarchical Bayesian analysis of population structure. Nucleic Acids Res. 2019; 47(11):5539–49. https://doi.org/10.1093/nar/gkz361.
Weir BS, Cockerham CC. Estimating F -Statistics for the analysis of population structure. Evolution (NY). 1984; 38(6):1358–70. https://doi.org/10.1111/j.1558-5646.1984.tb05657.x
Willis SC, Winemiller KO, Montaña CG, Macrander J, Reiss P, Farias IP, Ortí F. Population genetics of the speckled peacock bass (Cichla temensis), South America’s most important inland sport fishery. Conserv Genet. 2015; 16(6):1345–57. https://doi.org/10.1007/s10592-015-0744-y
Winemiller KO. Fish ecology. In: Nierenberg WA, editor. Encyclopedia of environmental biology. vol. 2. San Diego: Academic Press; 1995. p.49–65.
Winemiller KO. Patterns of variation in life history among South American fishes in seasonal environments. Oecologia. 1989; 81(2):225–41. https://doi.org/10.1007/BF00379810
Authors

Kyara Martins Formiga[1][2][4] ![]()
![]() , Jacqueline da Silva Batista[1][2][4]
, Jacqueline da Silva Batista[1][2][4] ![]() and José Antônio Alves-Gomes[1][3][4]
and José Antônio Alves-Gomes[1][3][4] ![]()
[1] Instituto Nacional de Pesquisas da Amazônia (INPA), Coordenação de Biodiversidade (COBIO), Av. André Araújo, 2936, 69067-375 Manaus, AM, Brazil. (KMF) formiga@inpa.gov.br (corresponding author); (JSB) jac@inpa.gov.br; (JAAG) puraque@inpa.gov.br.
[2] Instituto Nacional de Pesquisas da Amazônia (INPA), Laboratório Temático de Biologia Molecular (LTBM), Av. André Araújo, 2936, 69067-375 Manaus, AM, Brazil.
[3] Instituto Nacional de Pesquisas da Amazônia (INPA), Laboratório de Fisiologia Comportamental e Evolução (LFCE), Av. André Araújo, 2936, 69067-375 Manaus, AM, Brazil.
[4] Instituto Nacional de Pesquisas da Amazônia (INPA), Programa de Pós-Graduação em Genética, Conservação e Biologia Evolutiva (PPG GCBEv/INPA), Av. André Araújo, 2936, 69067-375 Manaus, AM, Brazil.
Authors Contribution
Kyara Martins Formiga: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Jacqueline da Silva Batista: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
José Antônio Alves-Gomes: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Ethical Statement
All specimens were collected in accordance with Brazilian laws, using a collection license approved by the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA) via permit numbers 130 and 14278–1.
Competing Interests
The authors declare no competing interests.
How to cite this article
Formiga KM, Batista JS, Alves-Gomes JA. The most important fishery resource in the Amazon, the migratory catfish Brachyplatystoma vaillantii (Siluriformes: Pimelodidae), is composed by an unique and genetically diverse population in the Solimões-Amazonas River System. Neotrop Ichthyol. 2021; 19(1):e200082. https://doi.org/10.1590/1982-0224-2020-0082
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2021 The Authors.
Diversity and Distributions Published by SBI
![]() Submitted August 22, 2020
Submitted August 22, 2020
![]() Accepted February 23, 2021
Accepted February 23, 2021
by Alexandre Hilsdorf
![]() Epub March 31, 2021
Epub March 31, 2021