José David Rangel-Medrano[1] ![]() and Edna Judith Márquez[1]
and Edna Judith Márquez[1] ![]()
![]()
PDF: EN XML: EN | Cite this article
Abstract
The Neotropical freshwater catfish Pseudopimelodus atricaudus and Pseudopimelodus magnus are two recently discovered species endemic to the Colombian Magdalena-Cauca River basin. In this study, a set of 13 microsatellite loci were developed by using next generation sequence technology to assess the genetic diversity and population structure in P. atricaudus and test for cross-species amplification in P. magnus. Both species exhibited high genetic diversity (P. atricaudus: Na: 9.000 – 9.769 alleles/locus, Ho: 0.760 – 0.804, HE: 0.804 – 0.840; P. magnus:Na: 12.8 – 5.4 alleles/locus, Ho: 0.638 – 0.683, HE: 0.747 – 0.755) compared to the mean levels of genetic diversity reported for Neotropical Siluriformes, and lack of genetic differentiation among sampling sites within the Cauca River (P. atricaudus: F’ST=0.013 – 0.017, P > 0.05, D’est= -0.004 – 0.023, P > 0.05; P. magnus: F’ST= 0.031, P= 0.055; D’est= 0.045, P= 0.058). This work is the first insight on the diversity and the population genetics of species of the family Pseudopimelodidae and provides a framework to further population genetic and conservation analyses needed in this poorly studied family at the microevolutionary level.
Keywords: Colombia, Freshwater fish, Gene flow, Genetic diversity, Magdalena-Cauca basin.
Los bagres neotropicales Pseudopimelodus atricaudus y Pseudopimelodus magnus son dos especies recientemente descubiertas, endémicas de la cuenca Magdalena-Cauca en Colombia. En este estudio, se desarrollaron 13 loci microsatélites usando tecnología de secuenciación de próxima generación para evaluar la diversidad genética y la estructura poblacional de P. atricaudus y evaluar su amplificación cruzada en P. magnus. Ambas especies exhibieron altos valores de diversidad genética (P. atricaudus: Na: 9.000 – 9.769 alelos/locus, HO: 0.760 – 0.804, HE: 0.804 – 0.840; P. magnus:Na: 12.8 – 5.4 alelos/locus, HO: 0.638 – 0.683, HE: 0.747 – 0.755) comparados con los valores promedios de diversidad genética reportados para Siluriformes neotropicales, y ausencia de estructura genética entre los sitios analizados (P. atricaudus: F’ST= 0.013 – 0.017, P > 0.05, D’est= -0.004 – 0.023, P > 0.05; P. magnus: F’ST= 0.031, P= 0.055; D’est= 0.045, P= 0.058). Este trabajo representa la primera aproximación a la diversidad y genética poblacional de especies de la familia Pseudopimelodidae y proporciona un marco de referencia para futuros estudios genético-poblacionales y de conservación, requeridos en esta familia de bagres poco estudiada en el nivel microevolutivo.
Palavras-chave: Colombia, Cuenca Magdalena-Cauca, Diversidad genética, Flujo génico, Pez dulceacuícola.
Introduction
The family Pseudopimelodidae (formerly assigned to Pimelodidae) is a Neotropical family that comprises seven genera and 50 valid species with a wide distribution in South America (Fricke et al., 2020). Pseudopimelodus Bleeker, 1858, one of the six genera recorded in Colombia (DoNascimiento et al., 2017), contains large and medium-sized species in a distribution range from the Atrato River in Colombia to the La Plata River in Argentina (Shibatta, 2003). Only two members of this genus have been recorded in commercial and artisanal fisheries from different Colombian basins: Pseudopimelodus bufonius (Valenciennes, 1840) from Atrato, Orinoco and Amazon river basins and Pseudopimelodus schultzi (Dahl, 1955) from Atrato-Sinú and Magdalena-Cauca river basins (DoNascimiento et al., 2017). However, a recent phylogeographic study based on the mitochondrial cox1 and the nuclear rag2 genes showed that Pseudopimelodus comprises at least five divergent lineages in Colombia (Rangel-Medrano et al., 2020): lineage 1 (Magdalena River and Cauca River lower sectors and the San Jorge River); lineage 2 (Amazon River hydrographic zone); lineage 3 (Orinoco River hydrographic zone); lineage 4 (Atrato River and Sinú River); lineage 5 (Magdalena River upper sector and, Cauca River upper and middle sectors). Based on these results, further osteological and external morphological analyses allowed the description of two new Pseudopimelodus species in Colombia: Pseudopimelodus atricaudus (lineage 1) Restrepo-Gómez, Rangel-Medrano, Márquez & Ortega-Lara, 2020 and Pseudopimelodus magnus (lineage 5) Restrepo-Gómez, Rangel-Medrano, Márquez & Ortega-Lara, 2020.
Currently, there is a gap in our knowledge of the biology, diversity, and genetic structure of members of the genus Pseudopimelodus, especially for the recently discovered species, which could hinder the development and implementation of effective management and conservation programs. In this context, prior knowledge about genetic variability is crucial for the proper population management of these two species (Hilsdorf, Hallerman, 2017). In the current literature, P. atricaudus and P. magnus have been identified as P. schultzi and categorized as least concern in the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; Mesa-Salazar, Sanchez-Duarte, 2016), mainly because it was thought that nominal P. schultzi had a wide distribution range. However, this species has been categorized as Near Threatened (NT) in the Colombian red list of freshwater fishes because of habitat degradation and pollution (Mojica et al., 2012). In addition, P. magnus can reach a large size, and plausibly could be harvested as an alternative fishery resource and consequently, might bear higher fishing pressure than the other species previously identified as P. schultzi. Therefore, it is important to characterize the genetic diversity of natural populations of these two species to reassess its current conservation status.
Population genetic studies are absent for all members of Pseudopimelodidae. Indeed, only one study developed and characterized a set of 13 microsatellite loci in Microglanis cottoides (Boulenger, 1891) and most of them were successfully cross-amplified in other Pseudopimelodid species (Souza-Shibatta et al., 2013). In contrast, some genetic studies have been conducted in the phylogenetically related family Pimelodidae showing gene flow and genetic structure. For example, using polymorphic microsatellite analyses, Ribolli et al. (2012) detected lack of genetic structure in Pimelodus maculatus Lacepède, 1803, from the upper Uruguay River. In addition, lack of genetic differentiation was observed in wild populations of Pseudoplatystoma corruscans (Spix & Agassiz, 1829) from the São Francisco River (Dantas et al., 2013) and within the Paraguay basins in Brazil (Prado et al., 2018). This later study also detected gene flow in Pseudoplatystoma reticulatum Eigenmann & Eigenmann, 1889from Paraguay basin in Brazil. Additionally, other studies in the genus Pseudoplatystoma Bleeker, 1862, and Steindachneridion Eigenmann & Eigenmann, 1919, have shown genetic structure likely explained by homing behavior, geographical accidents, disconnections among rivers, isolation by distance, or short-distance migration range (Sekine et al., 2002; Abreu et al., 2009; Pereira et al., 2009; Carvalho et al., 2012; Telles et al., 2014; Fonseca et al., 2017; Prado et al., 2018). Considering the geomorphological and topographic characteristics of the middle sector of the Cauca canyon in Colombia (Landínez-García, Márquez, 2016), Pseudopimelodus speciesinhabiting upstream and downstream of the rapids may exhibit population genetic structure. To tests this hypothesis, we developed a set of 13 microsatellite loci using next generation sequence technology in P. atricaudus and tested for cross-amplification in two populations of P. magnus from Magdalena-Cauca basin. The information obtained in this study, regarding genetic variability and extent of genetic structure, will help providing useful information for conservation and management programs aiming to protect natural populations of these bumblebee catfish species in Colombia.
Material and methods
Sampling. A total of 123 ethanol preserved muscle tissue samples of Pseudopimelodus (P. atricaudus:58; P. magnus: 65)previously studied by Rangel-Medrano et al. (2020) were submitted to microsatellite analyses. Collection information, voucher numbers and GenBank accessions for all sequences obtained were previously provided by Rangel-Medrano et al. (2020). Samples came from the middle and lower sections of the Cauca River (S1-S8) previously described by Landínez-García, Márquez (2016) (Fig. 1). Collection sites were located upstream and downstream in the Cauca River canyon, which is the steepest margin of the Antioqueño Plateau in the northern portion of the Central Cordillera (Restrepo-Moreno et al., 2009) and marks the boundary between the middle and lower sectors of the Cauca River. This landform has been considered a geographic barrier for many fish species (Dahl, 1971) and is the site of the largest hydropower project in Colombia (Ituango). Collection sites downstream the Cauca River canyon, are in sectors where the Cauca River flows through lower slopes and meanders through a more ample floodplain.
Microsatellite loci development. Total genomic DNA was extracted from muscle tissue of one individual of P. atricaudus collected in the Cauca River lower sector by using the DNeasy Blood & Tissue (Qiagen, Germany) and ~5 ng/µL of the extracted DNA was employed to prepare a genomic library that was subsequently sequenced by using the pyrosequencing process in a 454 FLX next generation sequence platform (Roche Applied Science) following Landínez-García et al. (2016). Then, we used the methodology described by Landínez-García, Márquez (2018) for cleaning of nuclear genome reads, primer design, electronic polymerase chain reaction (ePCR), polymorphism analysis, validation and amplification condition optimization for 30 microsatellite loci.

FIGURE 1| Sampling sites of Pseudopimelodus magnus and P. atricaudus in the middle and lower sectors of the Cauca River.
In brief, the software PAL_FINDER v0.02.03 (Castoe et al., 2010) was employed to detect and extract potential amplifiable loci (PAL) corresponding to reads containing perfect tri-, tetra- and pentanucleotide microsatellites loci. Next, the software PRIMER3 v.2.0 (Rozen, Skaletsky, 2000) was used for primer design in the flanking regions of each PAL setting a set of criteria as described in Castoe et al. (2010) such as GC content >30%; melting temperatures 58–65ºC with a maximum 2ºC difference between paired primers, the last two 3’ nucleotides were G or C and an average amplicon size of at least 100 bp. All other software settings were left at their default values. Finally, an electronic PCR (ePCR; Rotmistrovsky et al., 2004) was conducted to verify the correct alignment of each pair of primers with the corresponding DNA template.
A total of 30 microsatellite loci were selected to assess the level polymorphism in 15 samples of P. atricaudus from Cauca River based on standard PCR conditions (Sambrook, Russell, 2001). Amplicons were separated in 8% polyacrylamide gels, ran at 120 volts for 40 min using a Mini Protean® Tetra vertical electrophoresis cell (Bio-Rad Laboratories, Inc.) and visualized by silver staining. Furthermore, primers that showed consistent amplification, PCR products of 100–400 bp and ability to detect heterozygotes were selected for subsequent diversity and population structure analyses in P. atricaudus and tested for cross-amplification in P. magnus.
A set of 13 of 15 polymorphic loci were selected based on the above-mentioned criteria and used to assess the levels of genetic diversity and population structure in all collected samples of P. atricaudus and P. magnus.For this purpose, the 5’ end of each forward primer was labeled with either fluorescent dye FAM6, VIC, NED or PET (Applied Biosystems). Polymerase chain reactions were conducted using a final volume of 10 µL with ~30 ng/µL of genomic DNA, 1X of PCR buffer, 2.5 mM of MgCl2, 0.4 mM of dNTPs mix (Invitrogen™), 1 pmol/µL of each primer and 0.04 U/µL of Platinum Taq DNA Polymerase (Invitrogen™). Thermal conditions consisted of an initial denaturation step of 95ºC during 3 min, followed by 30 cycles of 94°C for 30 s, an annealing temperature of 54–56°C for 20 s and an extension step of 72°C for 18 s. An automated DNA sequencer ABI 3730 XL were used to separate all PCR products using the GeneScan™ 500 LIZ® as internal size standard. Finally, allelic scoring was conducted with the program GeneMapper v4.0 (Applied Biosystems). Each dataset was checked for potential genotyping errors including null alleles, allele dropout and scoring of stutter peaks by using the software Micro-Checker v2.2.052 (Van Oosterhout et al., 2004). The software CERVUS v3.0.7 (www.fieldgenetics.com) was employed to determine the polymorphic information content for each locus.
Genetic diversity and population structure analyses. Before performing diversity and genetic structure intraspecific analyses, all 123 samples of Pseudopimelodus and 10 loci that showed consistent amplification in both species were used to explore patterns of genotype distribution across all river sections analyzed. Structuring pattern was compared with barcoding results previously found by Rangel-Medrano et al. (2020) for confidently identifying both species. To this end, a Bayesian clustering method was implemented in software STRUCTURE v2.3.4 (Pritchard et al., 2000). The number of populations (K) was estimated using the admixture ancestry model, the LOCPRIOR option and correlated allele frequencies. Chain parameters included 20 independent runs using a burn-in of 50000 iterations followed by 500000 Markov chain Monte-Carlo (MCMC) chains assuming K=1 to K=7. The web-based software STRUCTURESELECTOR (Li, Liu, 2018) was used to calculate the most likely number of K based on the ∆K and LnP(K) methods previously described by Evanno et al. (2005) and the choose K algorithm (Raj et al., 2014). In addition, since uneven sampling often leads to wrong inferences on hierarchical structure and biased estimates of the true number of subpopulations, STRUCTURESELECTOR was also used to calculate the estimators MedMedK, MedMeaK, MaxMedK and MaxMeaK (Puechmaille, 2016). Finally, the integrated software CLUMPAK (Kopelman et al., 2015) was used to generate a graphical representation of the results.
Furthermore, STRUCTURE analyses were also conducted to examine intraspecific population structure in samples of P. atricaudus and P. magnus, using 13 and 10 loci, respectively. In addition, the degree of population genetic differentiation was calculated by the standardized statistics D’est (Meirmans, Hedrick, 2011) and F’st (Meirmans, 2006) and Analysis of Molecular Variance (AMOVA) using the software GenAlex v6.503. Finally, in order to explore alternative grouping patterns, the ADEGENET package (Jombart, 2008) implemented in R (www.r-project.org) was used to perform a Discriminant Analysis of Principal Components using all genotypes of both species.
Once all genotypes of P. atricaudus and P. magnus were confidently identified, genetic diversity analyses were conducted independently for both species. Genetic diversity indices such as number of alleles per locus (Na), observed (HO) and expected (HE) heterozygosity were calculated using the software GenAlex v6.502 (Peakall, Smouse, 2012). Estimation of population specific inbreeding coefficient (FIS), linkage disequilibrium among the pairs of loci and departures from Hardy-Weinberg expectations was performed by using a Markov chain-based approximations method in the software Arlequin v3.5.2.2 (Excoffier, Lischer, 2010). Finally, we adjusted the statistical significance for multiple comparisons by applying sequential Bonferroni correction (Holm, 1979; Rice, 1989).
Results
Microsatellite development. A total of 13 of 30 microsatellite loci evaluated were polymorphic, showed consistent amplification and absence of null alleles or genotyping errors in Pseudopimelodus atricaudus (Tab. 1). The number of alleles ranged from 5 (Patr03 and Patr11) to 17 (Patr05) with a mean of 8.769 alleles/locus and an average HO of 0.750. In addition, all loci met the Hardy-Weinberg and linkage assumptions. The PIC values ranged from 0.525 (Patr12) and 0.895 (Patr14) with an average of 0.803 (Tab. 1). Cross-amplification of these 13 microsatellite loci in Pseudopimelodus magnus showed successful amplification in 10 loci.
TABLE 1 | List of microsatellite loci developed for Pseudopimelodus atricaudus. The asterisk indicates lack of amplification in samples of Pseudopimelodus magnus. Na: number alleles per locus; HO and HE: observed and expected heterozygosity, respectively; PIC: polymorphism information content; P: statistical significance for tests of departure from Hardy-Weinberg equilibrium; FIS: inbreeding coefficient; P(FIS): statistical significance for FIS.
Locus | Repeat motif | Forward (F) and reverse (R) sequence 5’-3’ | Size Range (bp) | Na | Ho | He | PIC | P | FIS | P(FIS) |
Patr01 | (ATATC)n | F: TCTCTATTGCCCTATGGTGGG | 273-353 | 9 | 0.933 | 0.867 | 0.847 | 0.570 | -0.080 | 0.889 |
R: CATACATGCAGCACACTGGC |
|
|
|
|
|
|
|
| ||
Patr02 | (TCTCC)n | F: TCACTTTAAGAGAGGAAGACCCG | 95-140 | 7 | 0.667 | 0.738 | 0.714 | 0.090 | 0.100 | 0.342 |
R: GTGACAGTGGTTGGGTGAGC |
|
|
|
|
|
|
|
| ||
Patr03* | (ATCTT)n | F: TTGTGTTATCCCCTTGCTGG | 144-179 | 5 | 0.714 | 0.786 | 0.759 | 0.678 | 0.094 | 0.363 |
R: TCCCAGTTACCAGGTCATTTCC |
|
|
|
|
|
|
|
| ||
Patr04* | (ATTTT)n | F: GAAGTGTTACGGGCACTGAGC | 140-215 | 10 | 0.800 | 0.915 | 0.877 | 0.548 | 0.130 | 0.149 |
R: TCACTGAATTAACTGTGCAGCC |
|
|
|
|
|
|
|
| ||
Patr05 | (ATCT)n | F: GGAACAGCTGTGTAGGTGAAGC | 183-311 | 17 | 1.000 | 0.940 | 0.892 | 0.825 | -0.066 | 1.000 |
R: CCTTAGTGTCCTCTAAAGAGATCGG |
|
|
|
|
|
|
|
| ||
Patr06 | (ATCT)n | F: GAATGGCACAGACAGAAAGGC | 129-213 | 10 | 0.800 | 0.897 | 0.884 | 0.204 | 0.111 | 0.197 |
R: TGCACGAATTTCTCTTGGGG |
|
|
|
|
|
|
|
| ||
Patr07 | (ATCT)n | F: GCATCTCATCCCATACTTGAGC | 216-268 | 10 | 0.800 | 0.908 | 0.881 | 0.324 | 0.080 | 0.308 |
R: GCATAACCATTTCGCTCTGG |
|
|
|
|
|
|
|
| ||
Patr08 | (ATCT)n | F: TGAATGAGTAATGCCACTGCC | 220-268 | 8 | 0.786 | 0.852 | 0.851 | 0.862 | 0.123 | 0.156 |
R: ACTGGTAACCTTCCGGTCCC |
|
|
|
|
|
|
|
| ||
Patr09 | (ATCT)n | F: AAGGTGAGGTGAGATCTGTCAGG | 125-189 | 7 | 0.733 | 0.782 | 0.772 | 0.406 | 0.064 | 0.415 |
R: CCTTCACCTTCAGATAAACATTCC |
|
|
|
|
|
|
|
| ||
Patr10 | (AAAG)n | F: GAGACCATGCTGACACTCATCC | 157-269 | 9 | 0.600 | 0.674 | 0.713 | 0.592 | 0.113 | 0.264 |
R: AGAGAACTGACGTTTCCGCC |
|
|
|
|
|
|
|
| ||
Patr11 | (AATG)n | F: ATTCCTCCGTTGCTCTCAGC | 93-149 | 5 | 0.467 | 0.409 | 0.825 | 1.000 | -0.146 | 1.000 |
R: CACTGCTTCAGTCTCATGTTCG |
|
|
|
|
|
|
|
| ||
Patr12* | (ATCT)n | F: CCCCAGAGAACACAGAATAAACC | 141-197 | 8 | 0.733 | 0.832 | 0.525 | 0.716 | 0.123 | 0.211 |
R: AAAATATTTGCAGAAATAAGAGGGG |
|
|
|
|
|
|
|
| ||
Patr13 | (ATCT)n | F: AGCGTAATGGTGCCATAGCC | 199-251 | 9 | 0.733 | 0.878 | 0.850 | 0.195 | 0.170 | 0.081 |
R: CGCACTTAGAGGCGATTTACC |
|
|
|
|
|
|
|
|
Genetic structure. Genetic structure analysis using pooled samples of Pseudopimelodus atricaudus and P. magnus showed K=2 as the most likely number of clusters as indicated by ∆K (Mean LnP(K): -4991.64; Fig. 2A) and Puechmaille statistics (MedMedK=2, MedMeaK=2). Geographical distribution of genotypes revealed that P. atricaudus occurs predominantly in Cauca River lower sector, although one genotype of this species was found in the Cauca River middle sector (Fig. 2A). Conversely, P. magnus was present predominantly in Cauca River middle sector, although some genotypes were found coexisting with P. atricaudus in Cauca River S4 (Fig. 2A).
Genetic structure analysis at intraspecific level in both species (Figs. 2B–C), showed K=2 as the most likely number of populations, however, all samples of P. atricaudus (Mean LnP(K): -2489.26) and P. magnus (Mean LnP(K): -2489.26) were assigned to the same specific genetic cluster, suggesting that they correspond to a single genetic stock as confirmed by Puechmaille estimators (MedMedK=1; MedMeanK=1). For P. atricaudus, this result is concordant with the pairwise comparisons using the standardized F’ST and D’est statistics and Discriminant analysis of Principal Components, which detected lack of significant genetic differentiation among the sampling sites (Fig. 2E): S4-S5 (F’ST=0.017, P=0.308; D’est=0.023, P=0.305), S4-S8 (F’ST=0.013, P=0.856; D’est=-0.004, P=0.857) and S5-S8 (F’ST=0.015, P=0.861; D’est=0.011, P=0.862). Similarly, for P. magnus, STRUCTURE analysis is consistent with the results of standardized F’ST and D’est statistics (F’ST=0.031, P=0.055; D’est=0.045, P=0.058) and Discriminant Analysis of Principal Components (Fig. 2D).
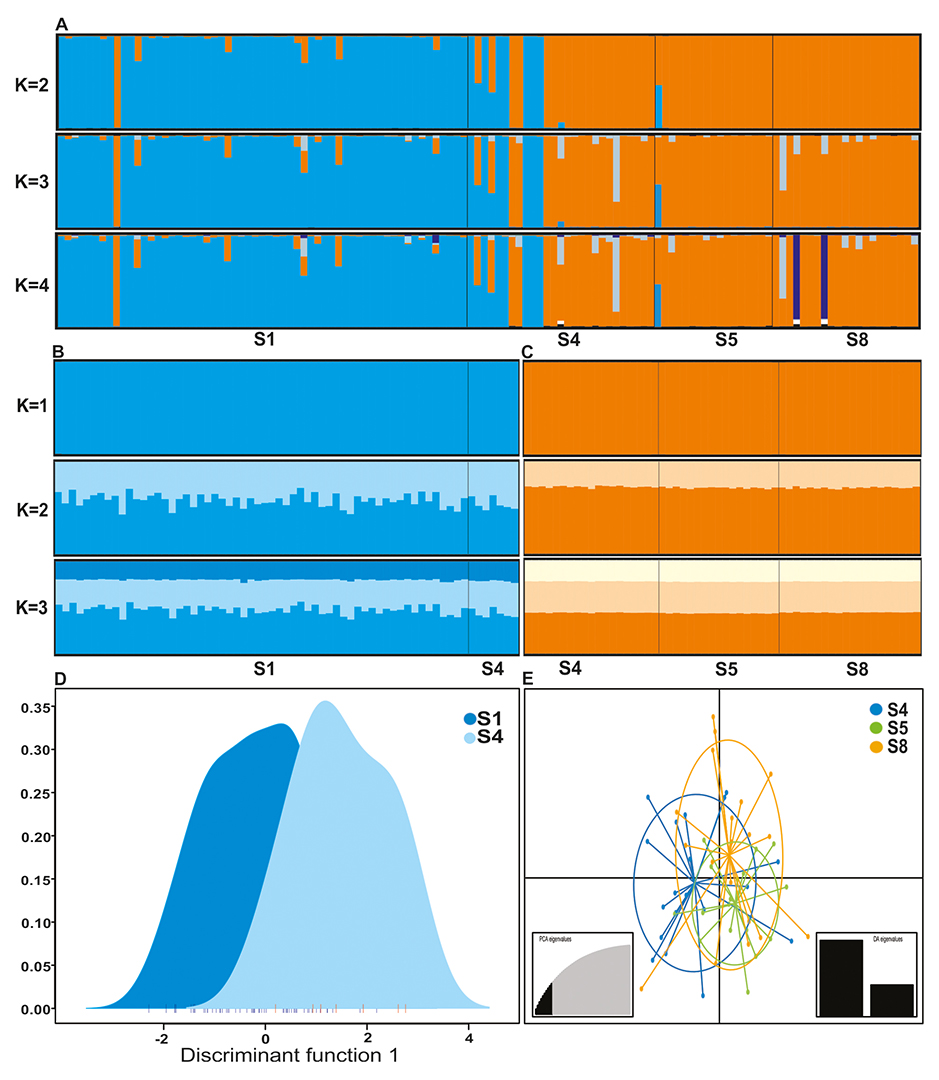
FIGURE 2 | Population structure suggested by STRUCTURE (A–C) and the Discriminant Analysis of the Principal Components (D, E) for Pseudopimelodus magnus (A, B, D), and Pseudopimelodus atricaudus (A, C, E). S1: sector 1 of the middle Cauca River; S4, S5, and S8: sectors 4, 5 and 8 of the lower Cauca River, respectively.
Genetic diversity. For Pseudopimelodus atricaudus (Tab. 2), all 13 microsatellite loci produced a total of 373 different alleles ranging from 4 (Patr11) to 18 (Patr05) alleles per locus. Levels of genetic diversity were higher in S4 (Na: 9.923; HO: 0.804, HE=0.840) followed by S8 (Na: 9.769; HO=0.798; He=0.825) and S5 (Na: 9.000; HO=0.760; HE=0.804). All loci satisfied Hardy-Weinberg and Linkage assumptions after sequential Bonferroni correction. Furthermore, values of population specific inbreeding coefficient were not significant in all sections evaluated for this species in the Cauca River (FIS: S4=0.023, P=0.245; S5=0.035; P=0.158, S8=0.011, P=0.355).
In Pseudopimelodus magnus (Tab. 3), a total of 182 different alleles were detected ranging from 25 (Patr05) to 3 (Patr11). Levels of genetic diversity were higher in Cauca River S1 (Na: 12.800; HO=0.683; HE=0.755) than in S4 (Na: 5.400; HO=0.638; HE=0.747). Furthermore, all loci satisfied Linkage and Hardy-Weinberg equilibria although 6 of 10 loci analyzed in Cauca River S1 showed departures from Hardy-Weinberg expectations after sequential Bonferroni correction. Finally, samples of both sites analyzed exhibited significant values of FIS (Cauca River S1 FIS=0.064, P=0.000; Cauca River S4 FIS=0.132; P=0.033).
TABLE 2 | Genetic diversity per locus and across loci of Pseudopimelodus atricaudus in three sites of the Cauca River. S4, S5 and S8: sectors 4, 5 and 8 of the lower Cauca River, respectively; N: Sampling size; Na: number alleles per locus; HO and HE: observed and expected heterozygosity, respectively; P: statistical significance for tests of departure from Hardy-Weinberg equilibrium; FIS: inbreeding coefficient; P(FIS): statistical significance for FIS.
Locus | S4 (N= 19) | S5 (N= 17) | S8 (N= 20) | |||||||||||||||
Na | HO | HE | P | FIS | P(FIS) | Na | HO | HE | P | FIS | P(FIS) | Na | HO | HE | P | FIS | P(FIS) | |
Patr01 | 10 | 0.895 | 0.852 | 0.233 | -0.052 | 0.826 | 9 | 0.941 | 0.884 | 0.578 | -0.067 | 0.860 | 9 | 0.900 | 0.887 | 0.324 | -0.015 | 0.692 |
Patr02 | 8 | 0.500 | 0.827 | 0.001 | 0.402 | 0.000 | 7 | 0.688 | 0.730 | 0.057 | 0.060 | 0.441 | 7 | 0.650 | 0.688 | 0.009 | 0.057 | 0.424 |
Patr03 | 6 | 0.611 | 0.754 | 0.060 | 0.194 | 0.108 | 5 | 0.625 | 0.796 | 0.193 | 0.221 | 0.082 | 8 | 0.706 | 0.859 | 0.012 | 0.183 | 0.058 |
Patr04 | 8 | 0.842 | 0.879 | 0.190 | 0.043 | 0.414 | 10 | 0.824 | 0.913 | 0.707 | 0.100 | 0.168 | 12 | 0.850 | 0.890 | 0.103 | 0.046 | 0.368 |
Patr05 | 16 | 0.947 | 0.882 | 0.295 | -0.076 | 0.939 | 18 | 1.000 | 0.925 | 0.961 | -0.084 | 1.000 | 17 | 0.842 | 0.905 | 0.356 | 0.071 | 0.278 |
Patr06 | 13 | 0.944 | 0.927 | 0.281 | -0.019 | 0.752 | 10 | 0.824 | 0.898 | 0.378 | 0.086 | 0.267 | 14 | 0.947 | 0.899 | 0.064 | -0.055 | 0.888 |
Patr07 | 11 | 0.842 | 0.876 | 0.632 | 0.040 | 0.451 | 11 | 0.824 | 0.916 | 0.379 | 0.104 | 0.172 | 12 | 0.850 | 0.919 | 0.562 | 0.077 | 0.244 |
Patr08 | 10 | 0.941 | 0.888 | 0.782 | -0.062 | 0.879 | 8 | 0.800 | 0.857 | 0.916 | 0.069 | 0.359 | 8 | 0.889 | 0.873 | 0.347 | -0.019 | 0.679 |
Patr09 | 10 | 0.684 | 0.838 | 0.129 | 0.188 | 0.050 | 8 | 0.765 | 0.783 | 0.581 | 0.023 | 0.550 | 9 | 0.750 | 0.795 | 0.142 | 0.058 | 0.363 |
Patr10 | 10 | 0.737 | 0.784 | 0.069 | 0.061 | 0.388 | 9 | 0.647 | 0.679 | 0.670 | 0.049 | 0.502 | 8 | 0.789 | 0.700 | 0.924 | -0.132 | 0.954 |
Patr11 | 7 | 0.778 | 0.667 | 0.529 | -0.172 | 0.926 | 5 | 0.412 | 0.367 | 1.000 | -0.126 | 1.000 | 4 | 0.550 | 0.586 | 0.252 | 0.063 | 0.453 |
Patr12 | 10 | 0.833 | 0.867 | 0.725 | 0.040 | 0.442 | 8 | 0.765 | 0.841 | 0.868 | 0.094 | 0.265 | 9 | 0.800 | 0.862 | 0.084 | 0.073 | 0.296 |
Patr13 | 10 | 0.895 | 0.883 | 0.246 | -0.013 | 0.698 | 9 | 0.765 | 0.863 | 0.302 | 0.117 | 0.195 | 10 | 0.850 | 0.867 | 0.053 | 0.020 | 0.536 |
Across loci | 9.923 | 0.804 | 0.840 | 0.012 | 0.023 | 0.245 | 9.000 | 0.760 | 0.804 | 0.817 | 0.035 | 0.158 | 9.769 | 0.798 | 0.825 | 0.001 | 0.011 | 0.355 |
TABLE 3 | Genetic diversity per locus and across loci of Pseudopimelodus magnus in two sitesof the Cauca River. S1 and S4: sectors 1 and 4 of the middle and lower Cauca River, respectively; N: Sampling size; Na: number alleles per locus; HO and HE: observed and expected heterozygosity, respectively; P: statistical significance for tests of departure from Hardy-Weinberg equilibrium; FIS: inbreeding coefficient; P(FIS): statistical significance for FIS.
Locus | S1 (N= 58) | S4 (N= 7) | ||||||||||
Na | HO | HE | P | FIS | P(FIS) | Na | HO | HE | P | FIS | P(FIS) | |
Patr01 | 14 | 0.828 | 0.891 | 0.147 | 0.072 | 0.087 | 6 | 0.714 | 0.857 | 0.194 | 0.178 | 0.288 |
Patr02 | 9 | 0.776 | 0.656 | 0.000 | -0.185 | 0.988 | 5 | 0.571 | 0.670 | 0.117 | 0.158 | 0.361 |
Patr05 | 25 | 0.839 | 0.897 | 0.023 | 0.065 | 0.096 | 7 | 0.714 | 0.868 | 0.102 | 0.189 | 0.234 |
Patr06 | 17 | 0.860 | 0.911 | 0.068 | 0.057 | 0.142 | 7 | 1.000 | 0.879 | 0.758 | -0.154 | 1.000 |
Patr07 | 9 | 0.707 | 0.827 | 0.029 | 0.146 | 0.024 | 5 | 0.571 | 0.769 | 0.296 | 0.273 | 0.160 |
Patr08 | 14 | 0.839 | 0.868 | 0.311 | 0.033 | 0.277 | 6 | 0.857 | 0.791 | 0.874 | -0.091 | 0.870 |
Patr09 | 10 | 0.839 | 0.800 | 0.889 | -0.050 | 0.839 | 5 | 0.571 | 0.725 | 0.548 | 0.226 | 0.274 |
Patr10 | 11 | 0.370 | 0.546 | 0.000 | 0.324 | 0.000 | 4 | 0.429 | 0.495 | 0.435 | 0.143 | 0.446 |
Patr11 | 6 | 0.259 | 0.492 | 0.000 | 0.477 | 0.000 | 3 | 0.286 | 0.615 | 0.104 | 0.556 | 0.055 |
Patr13 | 13 | 0.517 | 0.658 | 0.008 | 0.216 | 0.002 | 6 | 0.667 | 0.803 | 0.165 | 0.184 | 0.309 |
Across loci | 12.800 | 0.683 | 0.755 | < 0.001 | 0.064 | 0.000 | 5.400 | 0.638 | 0.747 | 0.164 | 0.132 | 0.033 |
Discussion
Microsatellite development. The present study developed a set of 13 polymorphic microsatellite loci to assess genetic diversity and population structure of the endemic bumblebee catfish species Pseudopimelodus atricaudus. This is the first study reporting the development of microsatellite markers in the genus Pseudopimelodus and the second in the family Pseudopimelodidae. PIC values ranged from 0.525 to 0.895 with an average of 0.803 which indicates that these loci are highly informative (Botstein et al., 1980). Although two additional loci showed good PIC values in the analyzed specimens, they were not considered in further analysis because of inconsistencies in the amplifications. However, 13 loci were used to confidently assess levels of genetic diversity in P. atricaudus and are, therefore, appropriate for further population genetic analyses in this species.
Furthermore, only 10 out of 13 microsatellite loci developed for P. atricaudus were successfully cross-amplified in P. magnus, since three loci failed to amplify in several individuals of this species, producing a high proportion of missing data. As described for the use of heterologous microsatellite loci in other taxa with distant phylogenetic relationships (Dakin, Avise, 2004), this differential amplification success may result from the high genetic divergence previously detected by nuclear and mitochondrial gene sequences (Rangel-Medrano et al., 2020). Despite this limitation, these 10 loci allowed us to estimate the levels of gene flow and diversity of P. magnus in the sampled sites.
Genetic structure and diversity. In concordance with previous phylogeographic studies (Rangel-Medrano et al., 2020), the Bayesian Structure analysis revealed two genetic clusters corroborating the predominance of Pseudopimelodus magnus and P. atricaudus in the middle and lower sections of Cauca River, respectively. This finding supports the hypothesis that the Cauca River canyon represents a significant barrier to dispersion. However, this study revealed the presence of genotypes of P. atricaudus and P. magnus in S4, suggesting the coexistence of both species in that sector of the river. Although not documented so far, the coexistence of these species may be facilitated by occasional downstream leakage of larvae of P. magnus from upper and middle sectors of the Cauca River. In addition, we also detected one genotype of P. atricaudus in Cauca River S1, however, the events that allowed the presence of this individual in the Cauca River middle sector are not yet clear. Moreover, our results showed that P. atricaudus and P. magnus are reproductively isolated as we found no sign of genetic admixture between them. However, to make a more robust assessment of this result, additional studies are encouraged using more genetic markers and a more exhaustive sampling across the Magdalena-Cauca River basin. Furthermore, additional studies are needed to determine if these species are reproductively isolated through mechanical or behavioral barriers, which are key components of speciation.
Specifically, for Pseudopimelodus atricaudus, all three methods to assess levels of population genetic structure revealed genetic homogeneity across sampling sites, not supporting our hypothesis of genetic structure. Similar results were found in P. magnus using the software STRUCTURE and the statistics D’est and F’st, although Discriminant Analysis of Principal Components showed a subtle genetic differentiation in samples from S1 and S4, likely caused by uneven sampling between these two localities. These findings are not surprising considering the middle-distance migratory range described for P. magnus and P. atricaudus, however, we encourage further genetic structure analyses including additional sampling sites of these species withing the Magdalena-Cauca River to have a better understanding of the genetic structure of these species. In addition, genetic structure results obtained in this study may not be comparable with those obtained in members of the phylogenetically related family Pimelodidae, mainly due to marked differences in their life cycles. For instance, it has been reported that members of Pseudopimelodidae are solitary botton-dwelling species with low vagility or sedentary habits (Esguícero, Arcifa, 2010) while some pimelodids such us Pimelodus maculatus and species of Pseudoplatystoma exhibit gregarious behavior and high migratory capacity (Carolsfeld et al., 2003; Sabinson et al., 2014).
Moreover, since all markers used in this study corresponded to tetra and pentanucleotide loci, average number of alleles per locus identified for P. atricaudus and P. magnus are not comparable with values detected in M. cottoides in which most of the loci analyzed included dinucleotide motifs in 24 individuals (Souza-Shibatta et al., 2013). However, average values per site of observed and expected heterozygosities detected (see Tabs. 2–3) were higher than those found in that cis-Andean species (Souza-Shibatta et al., 2013) and other phylogenetically related species of the family Pimelodidae, such as Brachyplatystoma rousseauxii (Castelnau, 1855)(Batista, Alves-Gomes, 2006), P. corruscans and P. reticulatum (Vaini et al., 2016; Prado et al., 2017). Nevertheless, these values of genetic diversity were similar to those found in the pimelodid species P. maculatus (Paiva, Kalapothakis, 2008; Ribolli et al., 2012). Overall, the results obtained in the present study showed that P. atricaudus and P. magnus exhibit a high genetic diversity compared to the global mean levels of genetic diversity reported for Neotropical Siluriformes (HE:0.609±0.210; Hilsdorf, Hallerman, 2017).
Since P. atricaudus is not considered an important fishery resource in the Magdalena-Cauca River basin, we suggest that the high genetic diversity detected in this species may be related to the low fishing pressure across its distribution range. Moreover, although we found good levels of genetic diversity in P. magnus, technical problems with the analysis suggest that species-specific primers must be designed to make a more precise assessment and for future monitoring of the genetic diversity of this species. This is important because P. magnus is currently being exploited for fish markets and local consumption given its large size, and so will be more likely to suffer a decrease in its genetic variability. In addition to fishing pressure, it is well known that river damming, as in the case of Ituango hydropower project in the Cauca River middle sector, is an important factor of freshwater habitat alteration due to the magnitude of freshwater ecosystem disturbances during and after dam construction (Nilsson et al., 2005; Liermann et al., 2012). The information obtained in this study constitute an important starting point for the formulation of future conservation and management policies and can be used to supplement conventional conservation assessments for these bumblebee catfish species in the Magdalena-Cauca River basin.
This study is the first report of a set of microsatellite loci for species of the genus Pseudopimelodus and the second within the family Pseudopimelodidae. Results obtained here showed that populations of the two new species P. atricaudus and P. magnus exhibit lack of genetic differentiation among sampling sites within the Cauca River and high levels of genetic diversity. However, given the deep genetic divergence detected between these two species, new diversity and genetic differentiation analyses are encouraged using species specific markers for P. magnus to avoid potential technical problems associated with genotyping.
Acknowledgments
This work was funded by the Universidad Nacional de Colombia, Sede Medellín and Empresas Públicas de Medellín, Grant CT–2013–002443–R1 “Variación genotípica y fenotípica de poblaciones de especies reófilas presentes en el área de influencia del proyecto hidroeléctrico Ituango”, Grant Convenio CT–2019–000661 “Variabilidad genética de un banco de peces de los sectores medio y bajo del Río Cauca”. We thank Dr. Donald Charles Taphorn for revising the English language and for providing some useful suggestions to the manuscript.
References
Abreu MM, Pereira LHG, Vila VB, Foresti F, Oliveira C. Genetic variability of two populations of Pseudoplatystoma reticulatum from the Upper Paraguay River Basin. Genet Mol Biol. 2009; 32(4):868–73. http://dx.doi.org/10.1590/S1415-47572009005000075
Batista JDS, Alves-Gomes JA. Phylogeography of Brachyplatystoma rousseauxii (Siluriformes – Pimelodidae) in the Amazon Basin offers preliminary evidence for the first case of “homing” for an Amazonian migratory catfish. Genet Mol Res. 2006; 5(4):723–40. Available from: http://www.funpecrp.com.br/gmr/year2006/vol4-5/pdf/gmr0231.pdf
Botstein D, White RL, Skolnick M, Davis RW. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am J Hum Genet.1980; 32:314–31. Available from: http://njc.rockefeller.edu/pdf3/BotsteinDavisAmJHumGenet1980.pdf
Carolsfeld J, Harvey B, Ross C, Baer A. Migratory fishes of South America: biology, fisheries and conservation status. Ottawa: World Bank; 2003. https://doi.org/10.1596/1-5525-0114-0
Carvalho DC, Oliveira DAA, Beheregaray LB, Torres RA. Hidden genetic diversity and distinct evolutionarily significant units in a commercially important Neotropical apex predator, the catfish Pseudoplatystoma corruscans. Conserv Genet. 2012; 13(6):1671–75. https://doi.org/10.1007/s10592-012-0402-6
Castoe TA, Poole AW, Gu W, Jason de Koning AP, Daza JM, Smith EN, Pollock DD. Rapid identification of thousands of copperhead snake (Agkistrodon contortrix) microsatellite loci from modest amounts of 454 shotgun genome sequence. Mol Ecol Resour. 2010; 10(2):341–47. https://doi.org/10.1111/j.1755-0998.2009.02750.x
Dahl G. Peces del Norte de Colombia. Bogotá: INDERENA. 1971.
Dakin EE, Avise JC. Microsatellite null alleles in parentage analysis. Heredity. 2004; 93(5):504–09. https://doi.org/10.1038/sj.hdy.6800545
Dantas HL, Neto MAS, Oliveira KKC, Severi W, Diniz FM, Coimbra MRM. Genetic diversity of captive and wild threatened catfish Pseudoplatystoma corruscans in the São Francisco River. Rev Fish Sci. 2013; 21(3–4):237–46. https://doi.org/10.1080/10641262.2013.800787
DoNascimiento C, Herrera-Collazos EE, Herrera-R GA, Ortega-Lara A, Villa-Navarro FA, Oviedo JSU, Maldonado-Ocampo JA. Checklist of the freshwater fishes of Colombia: a Darwin Core alternative to the updating problem. Zookeys.2017;708:25–138. http://doi.org/10.3897/zookeys.708.13897
Esguícero ALH, Arcifa MS. Biology and population features of a rare species of Pseudopimelodidae from the upper Paraná river basin. Biota Neotrop. 2010; 10(3):161–67. https://doi.org/10.1590/s1676-06032010000300018
Evanno G, Regnaut S, Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol. 2005; 14(8):2611–20. https://doi.org/10.1111/j.1365-294X.2005.02553.x
Excoffier L, Lischer HEL. Arlequin suite v3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour. 2010; 10(3):564–67. https://doi.org/10.1111/j.1755-0998.2010.02847.x
Fonseca FS, Domingues RR, Hallerman EM, Hilsdorf AWS. Genetic diversity of an imperiled neotropical catfish and recommendations for its restoration. Front Genet. 2017; 8(196):1–12. https://doi.org/10.3389/fgene.2017.00196
Fricke R, Eschmeyer WN, Van der Laan R. Eschmeyer’s catalog of fishes: genera, species, references [Internet]. San Francisco: California Academy of Science; 2020. Available from: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Hilsdorf AWS, Hallerman EM. Genetic Resources of Neotropical Fishes. New York; Springer International Publishing; 2017.
Holm S. A simple sequentially rejective multiple test procedure. Scand J Stat. 1979; 6(2):65–70. Available from: http://www.jstor.org/stable/4615733
Jombart T. ADEGENET: A R package for the multivariate analysis of genetic markers. Bioinformatics. 2008; 24(11):1403–05. https://doi.org/10.1093/bioinformatics/btn129
Kopelman NM, Mayzel J, Jakobsson M, Rosenberg NA, Mayrose I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol Ecol Resour. 2015; 15(5):1179–91. https://doi.org/10.1111/1755-0998.12387
Landínez-García RM, Alzate JF, Márquez EJ. Mitochondrial genome of the Neotropical detritivore fish Curimata mivartii Steindachner 1878 (Characiformes, Curimatidae). Mitochondrial DNA A DNA Mapp Seq Anal. 2016; 27(3):2196–97. https://doi.org/10.3109/19401736.2014.982623
Landínez-García RM, Márquez EJ. Development and characterization of 24 polymorphic microsatellite loci for the freshwater fish Ichthyoelephas longirostris (Characiformes: Prochilodontidae). PeerJ. 2016; 4:e2419. https://doi.org/10.7717/peerj.2419
Landínez-García RM, Márquez EJ. Microsatellite loci development and population genetics in Neotropical fish Curimata mivartii (Characiformes: Curimatidae). PeerJ. 2018; 6:e5959. https://doi.org/10.7717/peerj.5959
Li YL, Liu JX. StructureSelector: A web-based software to select and visualize the optimal number of clusters using multiple methods. Mol Ecol Resour. 2018; 18(1): 176–77. https://doi.org/10.1111/1755-0998.12719
Liermann CR, Nilsson C, Robertson J, Ng RY. Implications of dam obstruction for global freshwater fish diversity. BioScience. 2012; 62(6):539–48. https://doi.org/10.1525/bio.2012.62.6.5
Meirmans PG. Using the AMOVA framework to estimate a standardized genetic differentiation measure. Evolution.2006; 60(11):2399–402. https://doi.org/10.1111/j.0014-3820.2006.tb01874.x
Meirmans PG, Hedrick PW. Assessing population structure: FST and related measures. Mol Ecol Resour.2011; 11(1):5–18. https://doi.org/10.1111/j.1755-0998.2010.02927.x
Mesa-Salazar L, Sanchez-Duarte P. Pseudopimelodus schultzi. The IUCN Red List of Threatened Species [Internet]. Gland; 2016. Available from: https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T64792658A64890605.en
Mojica JI, Usma-Oviedo JS, Álvarez-León R, Lasso CA, editors. Libro rojo de peces dulceacuícolas de Colombia 2012. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt; 2012.
Nilsson C, Reidy CA, Dynesius M, Revenga C. Fragmentation and flow regulation of the world’s large river systems. Science. 2005; 308(5720):405–08. https://dx.doi.org/10.1126/science.1107887
Paiva LB, Kalapothakis E. Isolation and characterization of microsatellite loci in Pimelodus maculatus (Siluriformes: Pimelodidae). Mol Ecol Resour. 2008; 8(5):1078–80. https://dx.doi.org/10.1111/j.1755-0998.2008.02160.x
Peakall R, Smouse PE. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research — an update. Bioinformatics. 2012; 28(19):2537–39. https://doi.org/10.1093/bioinformatics/bts460
Pereira LHG, Foresti F, Oliveira C. Genetic structure of the migratory catfish Pseudoplatystoma corruscans (Siluriformes: Pimelodidae) suggests homing behaviour. Ecol Freshw Fish. 2009; 18(2):215–25. https://doi.org/10.1111/j.1600-0633.2008.00338.x
Prado FD, Fernandez-Cebrián R, Foresti F, Oliveira C, Martínez P, Porto-Foresti F. Genetic structure and evidence of anthropogenic effects on wild populations of two neotropical catfishes: baselines for conservation. J Fish Biol. 2018; 92(1):55–72. https://doi.org/10.1111/jfb.13486
Pritchard JK, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data. Genetics. 2000; 155:945–59. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1461096/pdf/10835412.pdf
Puechmaille SJ. The program STRUCTURE does not reliably recover the correct population structure when sampling is uneven: Subsampling and new estimators alleviate the problem. Mol Ecol Resour. 2016; 16(3):608–27. https://doi.org/10.1111/1755-0998.12512
Raj A, Stephens M, Pritchard JK. FastSTRUCTURE: Variational inference of population structure in large SNP data sets. Genetics. 2014;197(2):573–89. https://doi.org/10.1534/genetics.114.164350
Rangel-Medrano JD, Ortega-Lara A, Márquez EJ. Ancient genetic divergence in bumblebee catfish of the genus Pseudopimelodus (Pseudopimelodidae: Siluriformes) from northwestern South America. PeerJ. 2020; 8:e9028. http://doi.org/10.7717/peerj.9028
Restrepo-Moreno SA, Foster DA, Stockli DF, Parra-Sánchez LN. Long-term erosion and exhumation of the “Altiplano Antioqueño”, Northern Andes (Colombia) from apatite (U-Th)/He thermochronology. Earth Planet Sci Lett. 2009; 278(1–2):1–12. https://doi.org/10.1016/j.epsl.2008.09.037
Ribolli J, Melo CMR, Zaniboni-Filho E. Genetic characterization of the Neotropical catfish Pimelodus maculatus (Pimelodidae, Siluriformes) in the Upper Uruguay River. Genet Mol Biol. 2012; 35(4):761–69. https://doi.org/10.1590/S1415-47572012005000060
Rice WR. Analyzing tables of statistical tests. Evolution. 1989; 43(1):223–25. https://doi.org/10.2307/2409177
Rotmistrovsky K, Jang W, Schuler GD. A web server for performing electronic PCR. Nucleic Acids Res. 2004; 32(2):W108-W112. https://doi.org/10.1093/nar/gkh450
Rozen S, Skaletsky H. Primer3 on the WWW for general users and for biologist programmers. In: Misener S, Krawetz S, editors. Bioinformatics methods and protocols: methods in molecular biology. New Jersey: Humana Press; 2000. p.365–86.
Sabinson LM, Rodrigues-Filho JL, Peret AC, Verani JR. Growth and reproduction aspects of Pimelodus maculatus Lacépède, 1803 (Siluriformes, Pimelodidae) of the Cachoeira Dourada reservoir, state of Goiás and Minas Gerais, Brazil. Braz J Biol. 2014; 74(2):450–59. https://doi.org/10.1590/1519-6984.09012
Sambrook J, Russel DW. Molecular cloning: a laboratory manual. New York: Cold Spring Harbor Laboratory Press; 2001.
Sekine ES, Prioli A, Prioli S, Júlio H. Genetic differentiation among populations of Pseudoplatystoma corruscans (Agassiz, 1829) (Osteichthyes, Pimelodidae) isolated by the Guaíra Falls in the Paraná River. Acta Sci. 2002; 24(2):507–12. Available from: http://repositorio.uem.br:8080/jspui/bitstream/1/5222/1/375.pdf
Shibatta OA. Family Pseudopimelodidae (Bumblebee catfishes, dwarf marbled catfishes). In: Reis RE, Kullander SO, Ferraris CJ, Jr., editors. Check list of the freshwater fishes of South and Central America. Porto Alegre: Edipucrs; 2003. p.401–05.
Souza-Shibatta L, Ferreira DG, Oliveira C, Almeida FS, Shibatta OA, Sofia SH. Development and characterization of microsatellite loci of Microglanis cottoides (Siluriformes: Pseudopimelodidae) and cross-species amplification. Neotrop Ichthyol. 2013; 11(3):581–85. http://dx.doi.org/10.1590/S1679-62252013000300011
Telles MPC, Collevatti RG, Braga RS, Castro TG, Costa MC, Silva-Júnior NJD, Barthem R, Diniz-Filho JAF. Geographical genetics of Pseudoplatystoma punctifer (Castelnau, 1855) (Siluriformes, Pimelodidae) in the Amazon Basin. Genet Mol Res. 2014; 13(2):3656–66.
Vaini JO, Crispim BA, Silva DBS, Benites C, Russo MR, Grisolia AB. Genetic variability of pure Pseudoplatystoma corruscans and Pseudoplatystoma reticulatum individuals in the Paraná and Paraguay River basins. Fish Sci. 2016; 82(4):605–11. https://doi.org/10.1007/s12562-016-0999-3
Van Oosterhout C, Hutchinson WF, Wills DPM, Shipley P. Micro-Checker: software for identifying and correcting genotyping errors in microsatellite data. Mol Ecol Notes. 2004; 4(3):535–38. https://doi.org/10.1111/j.1471-8286.2004.00684.x
Authors

José David Rangel-Medrano[1] ![]() and Edna Judith Márquez[1]
and Edna Judith Márquez[1] ![]()
![]()
[1] Universidad Nacional de Colombia, Sede Medellín, Facultad de Ciencias, Escuela de Biociencias, Laboratorio de Biología Molecular y Celular Carrera 65 No 59A-110 Bloque 19A Laboratorio 310, 050034 Medellín, Antioquia, Colombia. (JDR) jdrangelm@unal.edu.co; (EJM) ejmarque@unal.edu.co, ejmarque@gmail.com (corresponding autor).
Authors Contribution
José David Rangel-Medrano: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing-original draft, Writing-review and editing.
Edna Judith Márquez: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing-original draft, Writing-review and editing.
Ethical Statement
Sample collections were performed framed under the environmental license # 0155 (January 30, 2009) from Ministerio de Ambiente, Vivienda y Desarrollo Territorial.
Competing Interests
The authors declare no competing interests.
How to cite this article
Rangel-Medrano JD, Márquez EJ. Development of microsatellite loci and population genetics in the bumblebee catfish species Pseudopimelodus atricaudus and Pseudopimelodus magnus (Siluriformes: Pseudopimelodidae). Neotrop Ichthyol. 2021; 19(1):e200053. https://doi.org/10.1590/1982-0224-2020-0053
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2021 The Authors.
Diversity and Distributions Published by SBI
![]() Submitted June 22, 2020
Submitted June 22, 2020
![]() Accepted December 16, 2020
Accepted December 16, 2020
by Claudio Oliveira
![]() Epub 08 March, 2021
Epub 08 March, 2021