![]() Violeta R. Perini2,
Violeta R. Perini2, ![]() Alessandro L. Paschoalini1,
Alessandro L. Paschoalini1, ![]() Nilo Bazzoli1,
Nilo Bazzoli1, ![]() Elizete Rizzo2 and
Elizete Rizzo2 and ![]() Daniel C. Carvalho1
Daniel C. Carvalho1 ![]()
PDF: EN XML: EN | Supplementary: S1 S2 | Cite this article
Abstract
River impoundments for electricity generation lead to environmental changes which severely affect fish migration and species richness. However, little is known about their effect on the genetic structure and population dynamics downstream from the reservoir. Here, we analyzed a set of ten microsatellite loci of Prochilodus lineatus, an important South American migratory fish. Specimens (n = 150) were sampled from five sites in a remnant lotic system that includes sections of the Grande, Pardo and Mogi Guaçu rivers, southeastern Brazil. The data showed that all microsatellites were polymorphic with the allele number per locus ranging from 5 to 32, and genetic diversity (He) varied from 0.74 to 0.80. Indices of genetic differentiation and Bayesian analysis showed a significant genetic structure and three genetic clusters inhabiting this river system. An asymmetric gene flow suggests source-sink metapopulation dynamics from tributaries (genetic source) to the main river (genetic sink). A genetic cluster that was not detected in the upper Mogi and Pardo rivers tributaries may indicate there is a “trapped gene pool” downstream from the Porto Colômbia dam. Thus, here we provide new insights into the genetic structure and population dynamics of a migratory fish species in a highly dammed river basin.
Keywords: Anthropogenic environmental impact, Metapopulation dynamics, Microsatellites, Migratory species, Neotropical.
Represamento de rios para geração de eletricidade levam a mudanças ambientais que afetam severamente a migração de peixes e riqueza de espécies. No entanto, pouco se sabe sobre seu efeito na estrutura genética e dinâmica populacional a jusante de reservatórios. Aqui, analisamos um conjunto de dez loci de microssatélites de Prochilodus lineatus, um importante peixe migratório sul-americano. Os espécimes (n = 150) foram amostrados em cinco locais de um sistema lótico remanescente que inclui seções dos rios Grande, Pardo e Mogi Guaçu, sudeste do Brasil. Os dados mostraram que todos microssatélites eram polimórficos com o número de alelos por locus variando de 5 a 32 e diversidade genética (He) variou de 0,74 a 0,80. Índices de diferenciação genética e análise de agrupamento baseada em modelo bayesiano indicou a presença de três agrupamentos genéticos habitando este sistema fluvial. Um fluxo gênico assimétrico sugere dinâmica metapopulacional de fonte-sumidouro dos tributários (fonte genética) para o rio principal (sumidouro genético). Um agrupamento genético que não foi detectado nos tributários rio Mogi e rio Pardo parecem indicar que há um “trapped gene pool” a jusante da represa de Porto Colômbia. Assim, nós provemos aqui novos conhecimentos sobre a estrutura genética e dinâmica populacional de uma espécie de peixe migratório em um rio altamente fragmentado por barramentos.
Palavras-chave: Dinâmica metapopulacional, Espécies migratórias, Impactos ambientais antropogênicos, Microssatétlites, Neotropical.
Introduction
Rivers are amongst the most heavily impacted ecosystems worldwide (Dudgeon et al., 2006; Geist, 2011). In South America, river impoundment is a major source of electricity generation, with Brazil having more than 700 large reservoirs (Agostinho et al., 2007). Due to a growing demand for electricity, it is expected that more dams are still to be built, and this is a cause of great environmental concern because dams act as barriers that interrupt migration routes used by several fish species (Petrere, 1996; Agostinho et al., 2005). Along the Grande River’s 1,300 km length (Upper Paraná River basin, Brazil), there are 13 hydroelectric power plants that have fragmented the river course and transformed it into a series of artificial reservoirs (Casali et al., 2010). One of these reservoirs is formed by the Porto Colômbia dam, which was constructed in 1973 and is located about 100 km from the next downstream reservoir, the Marimbondo dam (Fig. 1). The Grande, Pardo and Mogi Guaçu rivers in southeastern Brazil form a three-rivers system located downstream from Porto Colômbia dam, which is considered an important lotic remnant of the Grande River basin, essential to the reproduction and survival of member of the local freshwater fish assemblage, such as the Prochilodus lineatus (Valenciennes, 1837), Salminus brasiliensis (Cuvier, 1816), and Pimelodus maculatus Lacepède, 1803 (Godoy, 1972; Ham, Pearsons, 2001).
Theoretical simulation experiments have shown that river fragmentation by dams have the potential to affect population size and connectivity which may reduce genetic diversity and increase genetic differences among isolated riverine fish populations (Jager et al., 2001). Reduced genetic diversity has been detected in headwater populations of white-spotted char (Salvelinus leucomaenis (Pallas, 1814)) (Yamamoto et al., 2004) and river sculpin (Cottus gobio Linnaeus, 1758) isolated by dams within a river system located in Northeast England (Hänfling, Weetman, 2006). On the other hand, when there is a relatively short period of isolation (< 45 years) and relatively large amount of habitat still available to isolated upstream populations, no genetic differences are expected to be detected (Deiner et al., 2007).
The Neotropical fish Prochilodus lineatus is an interesting model to investigate human impacts on fish diversity since this species performs long distance reproductive migration to reach suitable places for spawning, as it is able to migrate more than 1,200 km during its breeding season (Godoy, 1972). Moreover, this species can attain a total length of up to 75 cm and is important for commercial fisheries, accounting for around 50% of the total fish biomass in the Grande River (Castro, Vari, 2003). Prochilodus lineatus usually migrate upriver during the reproductive season (November–February), leaving their feeding areas to find more appropriate places to reproduce. After reproduction, they migrate downstream back to feeding areas, where the schools remain until the next reproductive season (Godoy, 1972).
Several studies conducted on Neotropical dammed rivers have assessed the impact of impoundments on the genetic differentiation and diversity of fish from the Prochilodus Agassiz, 1829 genus (Wasko, Galetti Jr., 2002; Hatanaka, Galetti Jr., 2003; Ramella et al., 2006; Garcez et al., 2011; Ferreira et al., 2017). For instance, Garcez et al. (2011) tested how P. lineatus genetic diversity is allocated within the Grande and Mogi-Guaçu Rivers and possible genetic impacts due to river fragmentation by damming using polymerase chain reaction-restriction fragment length polymorphism (PCR‐RFLP). They sampled P. lineatus from river stretches between eight large hydroelectric dams and also tested if fish ladders would positively affect population connectivity and genetic diversity. They found a panmictic population and no genetic structure due to fragmentation by damming or influence of fish ladders on the genetic diversity of P. lineatus. Similarly, Ferreira et al. (2017), estimated genetic diversity, population differentiation and gene flow, in populations of P. lineatus, from six locations in the Upper and Middle Paraná River using microsatellites markers and mitochondrial haplotypes (D-loop). Since the authors observed high genetic diversity in all sample sites and no evidence of spatial structuring was correlated with habitat fragmentation, no negative influence of two large dams (Porto Primavera and Itaipu) was detected on the genetic diversity of P. lineatus.
However, population structuring related to seasonal stocks (winter and summer) has been detected for P. lineatus in the Lower Uruguay River, suggesting that the Lower Uruguay River may act as a resting area, where different genetic stocks use it as a feeding area (Rueda et al., 2013). Moreover, for the congeneric species Prochilodus costatus Valenciennes, 1850, from the São Francisco River (South-eastern Brazil), temporal structuring within a reproductive season was reported (Braga-Silva et al., 2016), suggesting isolation by time for this species. Therefore, different adaptation strategies can be present in the genus that may generate population structuring over time, but with no evidence for geographic isolation or isolation by distance.
Identifying the effects of barriers to gene flow and population dynamics are some of the major goals of conservation genetics, since the number of anthropogenic barriers, such as roads and dams, rises each day thus affecting population structure, dynamics, diversity, and ecosystem functions. Therefore, we used a set of ten microsatellite molecular markers to investigate the spatial-genetic structure, gene flow, and population dynamics of the Neotropical fish species P. lineatus from five sites of a remnant lotic system that includes sections of the Grande, Pardo and Mogi Guaçu Rivers in the context of anthropogenic impacts. Genetic diversity, inbreeding and bottleneck effect were also estimated to evaluate possible anthropogenic impacts, particularly downstream from the Porto Colômbia dam. The spatial-genetic population structure detected using two indices of genetic differentiation (FST and DEST) and Bayesian clustering approach provide evidence for a source-sink population dynamic for Prochilodus lineatus.
Material and methods
Fish sampling. A total of 150 adult specimens were caught at five sampling sites (Fig. 1) using gillnets placed overnight from December 2010 to February 2011: (S1) located in the Grande River immediately downstream from the Porto Colômbia dam (20º08’04”S 48º35’35”W); (S2) in the Grande River downstream from the confluence with the Pardo River (20º09’47”S 48º39’30”W); (S3) in the Pardo River, 110 km upstream from S1 (20º48’43”S 48º14’32”W); (S4) in the Mogi Guaçu River, in the region of Emas’ fall, approximately 200 km upstream from site S3 (21º55’34”S 47º22’03”W); (S5) in the Pardo River upstream from the confluence with the Mogi Guaçu River (20º53’27”S 48º08’20”W) (Fig. 1). Fresh adipose fins of P. lineatus specimens were collected and preserved in 95% ethanol for molecular analysis. Voucher specimens were deposited at the Laboratory of Conservation Genetics at Pontifícia Universidade Católica de Minas Gerais (PUC Minas) (LGC 4800, 6089, 6174, 6224, 6225, 6226, 6231).
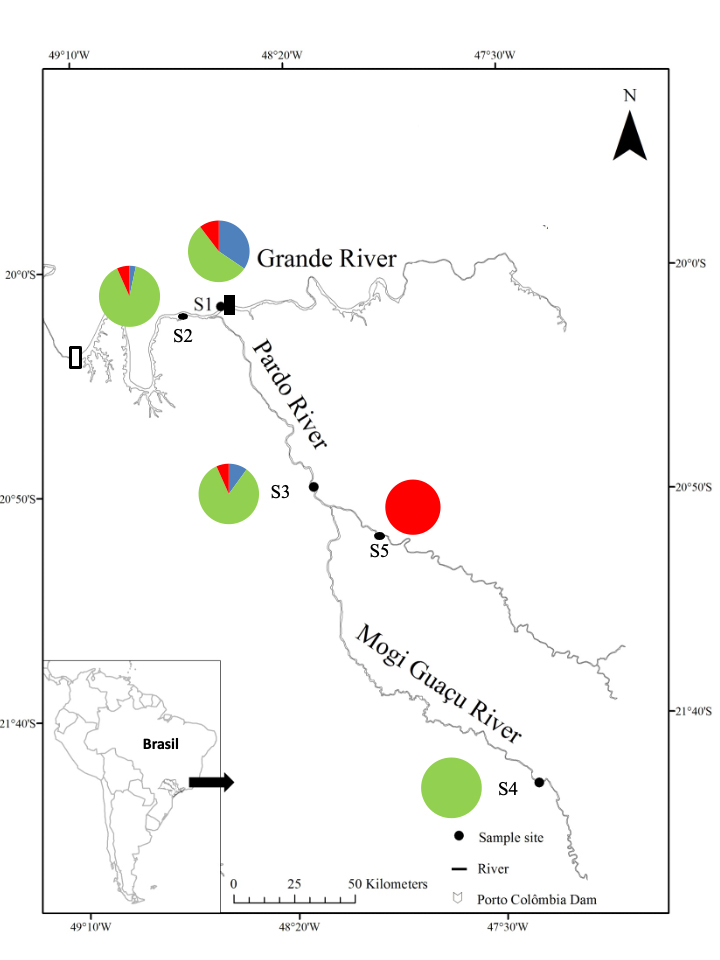
FIGURE 1 | Location of the sampling sites in the Grande River, southeastern Brazil and proposed population structuring model of Prochilodus lineatus from the Grande River basin. Pie graphs show the distribution of three genetic clusters assigned by Structure in the five geographic sampling sites: S1, Grande River immediately downstream from the Porto Colômbia dam; S2, Grande River downstream from the confluence with the Pardo River; S3, Pardo River downstream from the confluence with the Mogi Guaçu River; S4, Mogi Guaçu River, in the region of Emas’ fall; S5, Pardo River upstream from the confluence with the Mogi Guaçu River. Black rectangle = Porto Colômbia dam; White rectangle = Marimbondo dam. Genetic assigned clusters: Cluster A in blue; Cluster B in green, Cluster C in red.
DNA extraction and microsatellite analysis. Genomic DNA was extracted using a salt-based protocol according to Aljanabi, Martinez (1997). Five species-specific loci for P. lineatus (Yazbeck, Kalapothakis, 2007) and five heterologous loci for the genus Prochilodus (Barbosa et al., 2008) (Tab. 1) were used to design a multiplex reaction using the Multiplex Manager 1.0 software (Holleley, Geerts 2009). A polymerase chain reaction (PCR) was performed in three separated runs, following procedures described in Carvalho et al. (2009). Formamide and size standard LIZ (Applied Biosystems, USA) were combined with the amplified products and submitted to fragment analysis using an automated Applied Biosystems ABI 3130 sequencer. Profiles were manually scored using the Genemapper 4.0 software (Applied Biosystems, USA).
Data analysis. Microchecker 2.2 software (Van Oosterhout et al., 2004) was used to detect the presence of null alleles and possible scoring errors. Genepop 4 (Rousset, 2008) was utilized to estimate deviations from the Hardy-Weinberg equilibrium (HWE) by Markov’s method, to obtain the inbreeding coefficient (FIS), and to estimate deviations from the genotypic linkage disequilibrium (LD) across all sampled sites. Tests for deviations from HWE and LD were performed following Bonferroni’s correction (Rice, 1989). Genalex 6.5 (Peakall, Smouse, 2012) was used for estimating genetic parameters of each sampled site, such as: expected and observed heterozygosity (He, Ho), number of alleles in each population (Na), number of effective alleles (Ne), and number of private alleles (Tab. 2).
TABLE 1 | Microsatellite loci of Prochilodus lineatus (Pli) and P. argenteus (Par) used in the study and multiplexed in three distinct reactions (plex 1, plex 2, and plex 3). ªBarbosa et al., 2008; b Yazbeck, Kalapotakis, 2007.
Plex / Locus | Sequency (5’ ?
3’) | Range (pb) |
Plex 1 | ||
Par 86a | F: ATCCGTCTCTATGTGTGTC VIC R: TCTACAGTTACTTGGAGGAC | 91–137 |
Pli
61b | F: ACCAAGGTGGGATTCCTTGGTC 6-FAM R: TTCTCCATCTCTGGCTGCGCA | 208–218 |
Pli
30b | F: GATGTCGGTTCTTGTACAGTGGTG NED R: AGCTGCTGAGGATTCTGGGTCAC | 194–366 |
Pli
43b | F: AGTCCACTCCTTAGGCGAGTGAG PET R: ATAGACGGGCATGTGTCACAGCT | 190–246 |
Plex 2 | ||
Par 83ª | F: CATTTTCTAACAGCACTCC NED R: TTCTTGTTCTCCTGTGTAAC | 228–292 |
Par 85ª | F: CCACTTAATGAGACCACAC PET R: TTTCATTAGACTCGGTGAG | 195–261 |
Pli
60b | F: GCTAGGACGGTTAGCGTCCACTG VIC R: CGACACGTACATCATTACCTCGG | 174–226 |
Pli
34b | F: TGTTGGATGTAAAAAGTGC 6-FAM R: GCTCGCTGGCATGTTAAGATCC | 180–212 |
Plex 3 | ||
Par 80ª | F: CTAACCTACAAACCTCATTC VIC R: CTGTAAAAGCTCCACTTATC | 193–267 |
Par 66ª | F: TCTATAACTGTGGTCGTATG NED R: GAGGTTTTGAGATCAGTTG | 133–165 |
TABLE 2 | Genetic indexes in five sampling sites in the Grande River basin. Different letters indicate significant abiotic differences among sites (P < 0.05). Number of individuals (N), alleles (Na) and effective alleles (Ne), expected and observed heterozygosity (He, Ho).
| S1 – Grande River | S2 – Grande River | S3 – Pardo River | S4 – Mogi Guaçu
River | S5 – Upper Pardo River |
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
Genetic indexes | |||||
N | 28.9 ± 0.1 | 29.00 ± 0.47 | 29.80 ± 0.13 | 29.50 ± 0.40 | 29.40 ± 0.31 |
Na | 11.9 ± 2.07 | 12.90 ± 2.17 | 13.50 ± 2.09 | 12.80 ± 1.77 | 9.30 ± 1.37 |
Ne | 6.66 ± 1.41 | 8.11 ± 1.74 | 7.66 ± 1.58 | 7.66 ± 1.37 | 5.71 ± 1.05 |
Ho | 0.62 ± 0.05 | 0.69 ± 0.05 | 0.71 ± 0.06 | 0.73 ± 0.04 | 0.68 ± 0.07 |
He | 0.77 ± 0.05 | 0.80 ± 0.04 | 0.80 ± 0.04 | 0.81 ± 0.04 | 0.74 ± 0.05 |
FIS | 0.20629 ± 0.132 | 0.16141 ± 0.110 | 0.12445 ± 0.177 | 0.11905 ± 0.093 | 0.11975 ± 0.151 |
Spatial-genetic structure. To investigate the occurrence of genetic structure among the five populations of P. lineatus, the FST and DEST indices of genetic differentiation were calculated using Genalex 6.5. In order to test if FST and DEST values were correlated to the geographic distance between groups, the Mantel test (Mantel, 1967) was performed using geographic distances, along the rivers, between localities obtained from Google Earth.
To assess population structure as indicated by the number of genetically distinct populations (clusters), the Bayesian clustering approach implemented in Structure 2.2 (Pritchard et al., 2000) was used. This software calculates posterior probabilities for K clusters, assuming Hardy-Weinberg equilibrium conditions within clusters. The number of clusters was estimated using the “admixture” ancestral model with correlated alleles, allowing maximal population resolution, with K ranging from 1 to 10, to determine the optimal value of K. Five independent runs of 10,000 Markov chain Monte Carlo (MCMC) generations and 1,000 generations of burn-in were used for each value of K. The most likely number of clusters (K) was estimated using the Structure Havester software (Earl, vonHoldt, 2012).
Gene flow. To estimate the migration rate between each sampling site, we used the migration rate scaled by mutation rate estimated as single parameter (M) in Migrate 3.5.1 software (Beerli, Felsenstein, 2001). For the analyses, we used a maximum-likelihood method based on a coalescent approach (Beerli, Felsenstein, 1999), and the default settings of Migrate, with ten short Markov chains sampling 10,000 trees and three long Markov chains sampling 100,000 trees, following an initial burn-in period of 10,000 trees. Analyses were performed under the Brownian motion model as an approximation of the stepwise mutation model. All runs were repeated five times and the data presented are the mean ± standard error of the five replicate runs.
Testing for genetic bottlenecks. Heterozygote excess may be detected after a recent bottleneck event because initially the allelic diversity declines faster than heterozygosity due to the loss of rare alleles (Barson et al., 2009).Thus, population bottleneck was tested in the P. lineatus populations by performing a Wilcoxon’s signed rank test to calculate probabilities for heterozygote excess for two mutation models, Infinite Alleles Model (IAM) and Stepwise Mutation Model (SMM), using the Bottleneck 1.2.02 software (Piry et al., 1999), which was run for 10,000 iterations.
Results
A total of 150 P. lineatus specimens from five sampling sites (namely S1 to S5) were genotyped using a panel of ten microsatellite loci (Tab. 1). Genotypic linkage equilibrium tests across all populations were not significant (P < 0.05). All microsatellites were polymorphic with allele numbers per locus ranging from 5 (Pli61) to 32 (Pli30) (Tab. S1). Across all specimens, 179 alleles were detected, and 32 of them were private to their sampling site and in low frequency (P < 0.05). The two most frequent private alleles were detected in the S5 site: allele 346 (0.050 – Pli30) and allele 224 (0.054 – Par83) (Tab. S2).
The number of alleles (Na) per locus in each population ranged from 9.30 ± 1.37 (site S5) to 13.50 ± 2.09 (site S3), and the number of effective alleles (Ne) ranged from 5.71 ± 1.05 (site S5) to 8.11 ± 1.74 (site S2) (Tab. S1). The expected and observed heterozygosity (He and Ho) ranged from 0.74 ± 0.05 (site S5) to 0.81 ± 0.04 (site S4) and from 0.62 ± 0.05 (site S1) to 0.73 ± 0.04 (site S4), respectively (Tab. S1).
Deviations from the Hardy-Weinberg equilibrium were identified for three loci in site S1, two loci in site S2, three loci in site S3, one locus in site S4, and eight loci in site S5. No locus presented consistent deviations from the HWE in all the five sites studied, except for Par 86 (Tab. S1). Since the exclusion of Par 86 from the dataset did not alter our analysis, we opted to keep all markers in all further analysis. No evidence of scoring errors due to stuttering or allele dropout was found by Microchecker. Although null alleles were detected, they were not consistent across all populations for the same locus.
We used Bottleneck software to test for the heterozygote excess in each population as evidence of a possible recent bottleneck event. However, no significant heterozygote excess was recovered by the Wilcoxon’s test for both IAM and SMM mutation models (P < 0.05) (Tab. 3).
TABLE 3 | Wilcoxon test for detection of recent bottleneck events in the studied sites. IAM = Infinite Alleles Model; SMM = Stepwise Mutation Model.
Wilcoxon test | ||
| IAM | SSM |
Geographic Population | P values | |
S1 | 0.37 | 0.23 |
S2 | 0.08 | 1.00 |
S3 | 0.27 | 0.13 |
S4 | 0.08 | 0.27 |
S5 | 0.10 | 0.85 |
Spatial-genetic structure. The genetic differentiation among groups of P. lineatus ranged from 0.009 to 0.022 (FST), with significant differences (P < 0.05) observed for 5 of 10 pairwise comparisons, and the DEST indices values ranged from 0.009 to 0.095, with significant differences (P < 0.05) observed for the same pairwise comparisons (Tab. 4). These indices suggested site S5 (Pardo River) is the sampled site with fish most genetically differentiated from the other four sites (Tab. 4). Both FST and DEST indices also showed a significant difference between the specimens from sites S1 and S4.
No significant correlation between geographical distance among sites and genetic divergence estimated by FST (r² = 0.1454, P > 0.05) and DEST (r² = 0.1437, P > 0.05) was recovered, thus our Mantel test rejected the hypothesis of isolation by distance. The Bayesian model-based clustering analysis indicated the presence of three genetic clusters (K = 3), namely: Cluster A (blue), B (green) and C (red) (Figs. 2A,B). Only cluster C was clearly differentiated from the others, but, on the other hand, clusters A and B appear in admixed form. Consistent with the pairwise FST values reported above, Structure results also suggested that the population from site S5 was the most differentiated, composed entirely of individuals assigned to Cluster C. Fish from sites S2, S3 and S4 showed a higher probability of assignment to Cluster B (Fig. 2B).
TABLE 4 | FST (above diagonal) and DEST (below diagonal) values between pairs are shown for Prochilodus lineatus populations. *Significant values (P < 0.05).
| S1 | S2 | S3 | S4 | S5 |
S1 | 0.00000 | 0.01221 | 0.00892 | 0.01439* | 0.01603* |
S2 | 0.01524 | 0.00000 | 0.00854 | 0.01005 | 0.01367* |
S3 | -0.00869 | -0.01089 | 0.00000 | 0.00849 | 0.01415* |
S4 | 0.03642* | 0.00197 | -0.00976 | 0.00000 | 0.02176* |
S5 | 0.04082* | 0.02869* | 0.03410* | 0.09502* | 0.00000 |
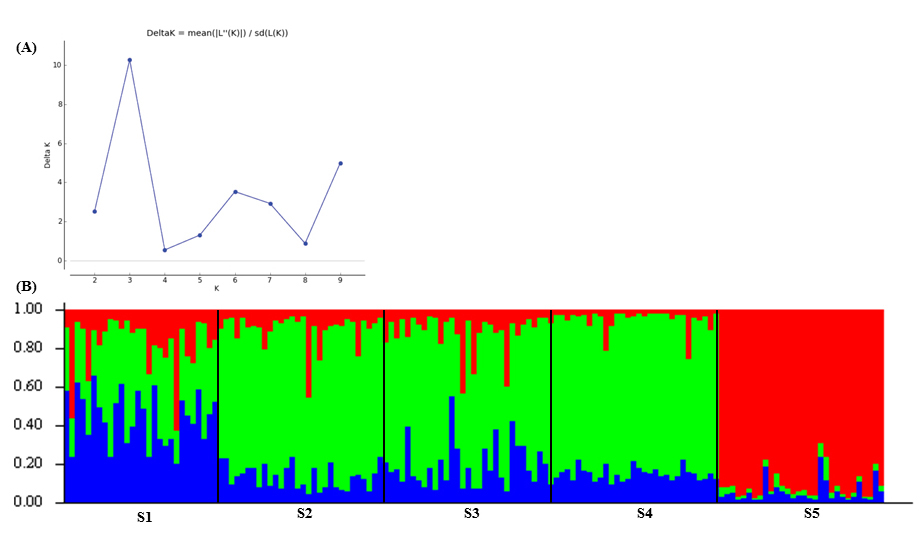
FIGURE 2 | Results of the Structure 2.2 analysis for Prochilodus lineatus populations from the Grande River basin. A. Estimated Delta K (= mean (|L”(K)|)/sd(L(K))) versus the number of populations, K. Each point is the mean of five independent runs of 10000 Markov chain Montecarlo generations and 1000 generations of ‘burn-in’. B. Barplot showing posterior probability of membership of P. lineatus in the three clusters identified by Structure. Each individual is represented by a single vertical bar in which the color indicates the individual’s estimated membership to alternative clusters. Black vertical lines separate the five sites, and the name of each locality of origin is indicated below. Cluster A in blue; Cluster B in green, Cluster C in red. See the names of the sampling sites in Fig. 1.
Migration rates. Evidence for source-sink population structure was detected from the asymmetric immigration–emigration patterns, a measure of the degree to which a population is a donor or a recipient of migrants. The main Grande River (sites S1 and S2) tended to be recipients of fish specimens (sink population), presenting positive scaled migration rates (2.80 ± 0.75 and 4.29 ± 0.84, respectively), which indicates that immigration rates are greater than emigration rates in these areas. On the other hand, sampled sites S3, S4 and S5 from tributaries tended to be donors (source population) and showed negative migration rates (-1.65 ± 0.06, -1.29 ± 0.04 and -4.16 ± 0.23, respectively) (Fig. 3).
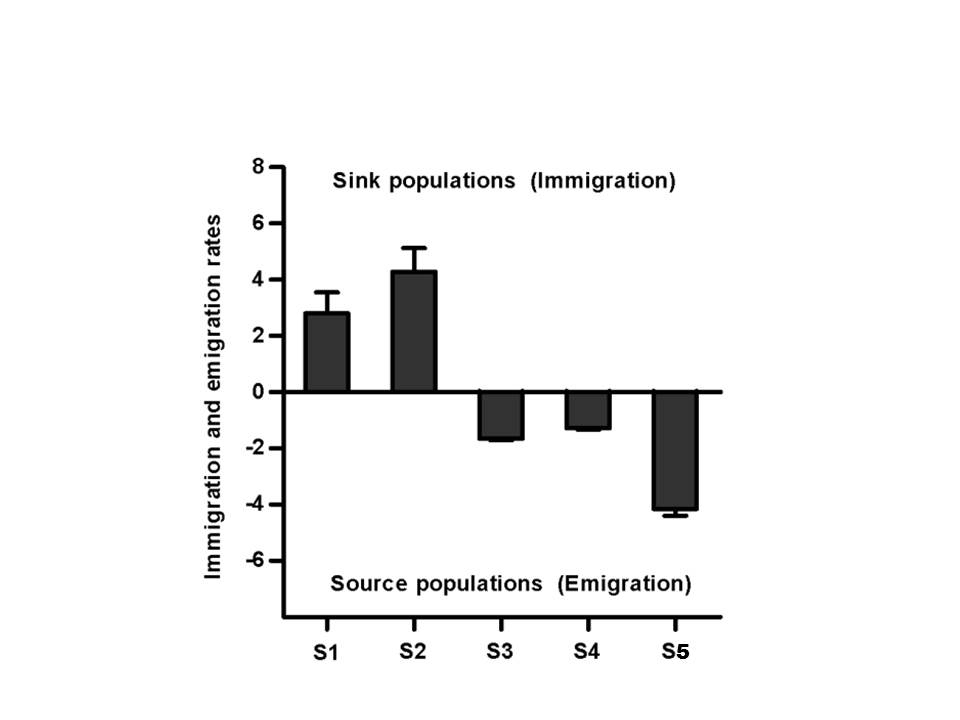
FIGURE 3 | Asymmetry in average proportions of immigrant and emigrant gene flow in five populations of Prochilodus lineatus from the Grande River basin. Bars show the mean difference between scaled immigration and emigration rate per population ± standard error. Positive values represent sink populations, with a relative excess of immigrants; and negative values represent source populations with a relative excess of emigrants.
Discussion
Considering the continuous river system with no physical barriers among the five sampling sites of the present study, and a previous report on mtDNA (Sivasundar et al., 2001) and PCR-RFLP analyses (Garcez et al., 2011) showing no population structuring for P. lineatus in the Grande River, a single panmictic population of P. lineatus would be expected within the Grande-Pardo-Mogi Guaçu River system. Interestingly, the FST and DEST pairwise values of genetic differentiation and Bayesian analysis showed a significant genetic structure and three genetic clusters inhabiting this river system. Despite the absence of barriers, genetic structuring within fish populations can occur across very short geographical distances (Angers, Bernatchez, 1998; Koskinen et al., 2001, 2002; Primmer et al., 2006). Genetic structuring with no evident physical barrier has also been demonstrated for several migratory neotropical fish species (Wasko, Galetti Jr., 2002; Hatanaka et al., 2006; Sanches, Galetti Jr, 2007; Piorski et al., 2008; Pereira et al., 2009; Sanches et al., 2012). Although different migratory populations occupy and explore the same river, they segregate during the spawning season, and consequently, population structuring is maintained (Pereira et al., 2009; Sanches et al., 2012), as has also been shown for P. lineatus in the lower Uruguay River (Rueda et al., 2013).
Such genetic structure in a continuous river system with no physical barriers among sampling sites could be due to source-sink effect, homing behavior, environmental factors (Gaggiotti et al., 2009; Sanches et al., 2012), due to distinct seasonal stocks (Rueda et al., 2013) or spawning waves consisting of different genetic populations (Braga-Silva et al., 2016).
Here, we have evidence for source-sink population dynamics within P. lineatus provided by asymmetric immigration-emigration patterns. Populations from the tributaries (sites S3, S4, and S5) were considered as source populations (donor of migrants) while sites S1 and S2 located in the main river were considered sink populations (recipient of migrants – Fig. 3). These results may be explained by a migratory route between the main river and the tributary, since mark-recapture studies have shown that P. lineatus uses tributaries for reproduction and the main river for feeding purposes (Godoy, 1972).
The genetic structure observed in our study (FST ranging from 0.9 to 2.2% and DEST ranging from 0.9 to 9.5%, (P < 0.05) indicates site S5 (Pardo River) as a distinct population. Sites S1, S2, S3, and S4 (Grande and Mogi Guaçu rivers) showed greater similarity between populations, composed mainly of individuals from Cluster B. On the other hand, the upper Pardo River (S5), not yet investigated by mark-recapture studies, was entirely composed of individuals assigned to Cluster C, showing the most differentiation of all the five sampling sites. Thus, our results showed that the P. lineatus populations from the lower and middle sections of the Grande-Pardo-Mogi Guaçu rivers system are composed of at least three genetic stocks that may be using it as a feeding area, but in the upper sections, these stocks segregate to each tributary.
Interestingly, within all three genetic pools identified for the Grande-Pardo-Mogi River system, one gene pool (Fig. 1, site S1 – Cluster A) was not found in the upper sections of this river system and were mainly detected downstream from the Porto Colômbia dam. This gene pool (Cluster A) may represent an ancient population that is not able to reproduce in this modified environment and could be related to an upstream population that lost connectivity after the Porto Colômbia dam closure. Thus, we speculate that the downstream Porto Colômbia population is partially composed of a “trapped gene pool”, which were not detected at the upper Mogi and Pardo tributaries (Fig. 1, Cluster A). Thus, fish from Cluster A which once performed an upstream Grande River migration to reproduce, before the construction of the Porto Colômbia dam use other tributaries, but now cannot perform such migration and were not able to adapt to the new environmental conditions. This may also explain the higher endogamic indices (FIS) and the lack of this genetic pool (Cluster A) in the upper sections of the Grande-Pardo-Mogi Guaçu rivers system. Moreover, sites S1 and S2, located in the area influenced by the Porto Colômbia dam presented the highest FIS inbreeding coefficient. Moreover, fish from the site located downstream from the dam (S1) had more fat deposits (P < 0.05) (Perini et al., 2013), lower heterozygosity and higher inbreeding coefficient (Tab. 2), suggesting a “resident population” in this region that do not perform upstream reproductive migration to the available tributaries. Such “resident population” was found to be a sink population in our metapopulation dynamics model, supporting our hypothesis of a “trapped gene pool” downstream from the Porto Colômbia dam.
Theoretically, inbreeding reduces heterozygosity and exposes deleterious recessive alleles, consequently, recessive deleterious homozygotes are more frequent than in a random mating population (Frankham et al., 2010). Inbreeding is a conservation concern as it reduces reproductive fitness, increasing the risk of extinction. However, considering that inbreeding increases more rapidly in small populations, this would not be a conservation problem for P. lineatus because it possesses one of the largest fish populations in the Grande River basin, with no signal of bottlenecks (Tab. 3). The data presented here provide new insights into the genetic structure and population dynamics of a migratory fish species in a highly dammed river basin. Moreover, more extensive sampling is needed for conclusive results, and therefore, we suggest analyzing a broader range of P. lineatus populations in the Grande River, including populations above and below dams, to provide a more complete picture of the population structure and population dynamics of species affected by intense habitat fragmentation.
Acknowledgments
The authors are thankful to Dr. Dirceu Marzulo, Dr. José A. Senhorini and the fish team of the Hydrobiology and Hatchery Station of Furnas and the Centro Nacional de Pesquisa e Conservação de Peixes Continentais/CEPTA, for supporting the fish collections. Thanks also to Stephen Latham for suggestions on the English version and to the editor Dr. Fernando Pelicice and anonymous reviewer for invaluable suggestions during the review process. The research was supported by grants from the Brazilian agencies: CNPq, FAPEMIG and CAPES. The authors are grateful to CNPq felowships (310283/2020-5; 306946/2016-5; 306155/2018–4).
References
Agostinho AA, Gomes LC, Pelicice FM. Ecologia e manejo de recursos pesqueiros em reservatórios do Brasil. Maringá: EDUEM; 2007.
Agostinho AA, Thomaz SM, Gomes LC. Conservação da biodiversidade em águas continentais do Brasil. Megadiversidade. 2005; 1(1):70–78.
Aljanabi SM, Martinez I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997; 25(22):4692–93. https://doi.org/10.1093/nar/25.22.4692
Angers B, Bernatchez L. Combined use of SMM and non-SMM methods to infer fine structure and evolutionary history of closely related brook charr (Salvelinus fontinalis, Salmonidae) populations from microsatellites. Mol Biol Evol. 1998; 15(2):143. https://doi.org/10.1093/oxfordjournals.molbev.a025911
Barbosa ACDR, Galzerani F, Corrêa TC, Galetti Jr. PM, Hatanaka T. Description of novel microsatellite loci in the Neotropical fish Prochilodus argenteus and cross-amplification in P. costatus and P. lineatus. Genet Mol Biol. 2008; 31(1 Suppl.):357–60. https://doi.org/10.1590/S1415-47572008000200032
Barson NJ, Cable J, Van Oosterhout C. Population genetic analysis of microsatellite variation of guppies (Poecilia reticulata) in Trinidad and Tobago: Evidence for a dynamic source–sink metapopulation structure, founder events and population bottlenecks. J Evol Biol 2009; 22(3):485–97. https://doi.org/10.1111/j.1420-9101.2008.01675.x
Beerli P, Felsenstein J. Maximum-Likelihood estimation of migration rates and effective population numbers in two populations using a coalescent approach. Genetics. 1999; 152(2):763–73. https://doi.org/10.1093/genetics/152.2.763
Beerli P, Felsenstein J. Maximum likelihood estimation of a migration matrix and effective population sizes in n subpopulations by using a coalescent approach. PNAS USA. 2001; 98(8):4563–68. https://doi.org/10.1073/pnas.081068098
Braga-Silva A, Galetti Jr., PM. Evidence of isolation by time in freshwater migratory fish Prochilodus costatus (Characiformes, Prochilodontidae). Hydrobiologia. 2016; 765(1):159–67. https://doi.org/10.1007/s10750-015-2409-8
Carvalho DC, Oliveira DAA, Sampaio I, Beheregaray LB. Microsatellite markers for the Amazon peacock bass (Cichla piquiti). Mol Ecol Resour. 2009; 9(1):239–41. https://doi.org/10.1111/j.1755-0998.2008.02425.x
Casali RCV, Vono V, Godinho HP, Luz RK, Bazzoli N. Passage and reproductive activity of fishes in the Igarapava fish ladder, Grande River, Southeastern Brazil. River Res Appl. 2010; 26(2):157–65. https://doi.org/10.1002/rra.1242
Castro RMC, Vari RP. Family Prochilodontidae. In: Reis RE, Kullander SO, Ferraris CJ, Jr., editors. Check list of the freshwater fishes of South and Central America. Porto Alegre: Edipucrs; 2003. p.65–70.
Deiner K, Garza JC, Coey R, Girman DJ. Population structure and genetic diversity of trout (Onchorynchus mykiss) above and below natural and man-made barriers in the Russian River, California. Conserv Genet. 2007; 8(2):437–54. https://doi.org/10.1007/s10592-006-9183-0
Dudgeon D, Arthington AH, Gessner MO, Kawabata Z-I, Knowler DJ, Lévêque C, Naiman RJ, Prieur-Richard A-H, Soto D, Stiassny MLJ, Sullivan CA. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol Rev. 2006; 81(2):163–82. https://doi.org/10.1017/S1464793105006950
Earl DA, vonHoldt BM. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour. 2012; 4(2):359–61. https://doi.org/10.1007/s12686-011-9548-7
Ferreira DG, Souza-Shibatta L, Shibatta OA, Sofia SH, Carlsson J, Dias JHP, Makrakis S, Makrakis MC. Genetic structure and diversity of migratory freshwater fish in a fragmented Neotropical river system. Rev Fish Biol Fish. 2017; 27(1):209–31. https://doi.org/10.1007/s11160-016-9441-2
Frankham R, Ballou JD, Briscoe DA. Introduction to conservation genetics. 2nd ed. Cambridge: Cambridge University Press; 2010. https://doi.org/10.1017/CBO9780511809002
Gaggiotti OE, Bekkevold D, Jørgensen HBH, Foll M, Carvalho GR, Andre C, Ruzzante DE. Disentangling the effects of evolutionary, demographic, and environmental factors influencing genetic structure of natural populations: Atlantic herring as a case study. Evolution. 2009; 63(11):2939–51. https://doi.org/10.1111/j.1558-5646.2009.00779.x
Garcez R, Calcagnotto D, Almeida-Toledo LF. Population structure of the migratory fish Prochilodus lineatus (Characiformes) from rio Grande basin (Brazil), an area fragmented by dams. Aquat Conserv. 2011; 21(3):268–75. https://doi.org/10.1002/aqc.1176
Geist J. Integrative freshwater ecology and biodiversity conservation. Ecol Indic. 2011; 11(6):1507–16. https://doi.org/10.1016/j.ecolind.2011.04.002
Godoy MP. Brazilian tagging experiments, fishes migration, and upper Paraná River basin eco-system. Rev Bras Biol. 1972; 32(4):473–84.
Ham KD, Pearsons TN. A practical approach for containing ecological risks associated with fish stocking programs. Fisheries. 2001; 26(4):15–23. https://doi.org/10.1577/1548-8446(2001)026%3C0015:APAFCE%3E2.0.CO;2
Hänfling B, Weetman D. Concordant genetic estimators of migration reveal anthropogenically enhanced source-sink population structure in the river sculpin, Cottus gobio. Genetics. 2006; 173(3):1487–501. https://doi.org/10.1534/genetics.105.054296
Hatanaka T, Galetti Jr. PM. RAPD markers indicate the occurrence of structured populations in a migratory freshwater fish species. Genet Mol Biol. 2003; 26(1):19–25. https://doi.org/10.1590/S1415-47572003000100004
Hatanaka T, Henrique-Silva F, Galetti Jr. PM. Population substructuring in a migratory freshwater fish Prochilodus argenteus (Characiformes, Prochilodontidae) from the São Francisco River. Genetica. 2006; 126(1):153–59. https://doi.org/10.1007/s10709-005-1445-0
Holleley CE, Geerts PG. Multiplex Manager 1.0: A cross-platform computer program that plans and optimizes multiplex PCR. Biotechniques. 2009; 46(7):511–17. https://doi.org/10.2144/000113156
Jager HI, Chandler JA, Lepla KB, Van Winkle W. A theoretical study of river fragmentation by dams and its effects on White sturgeon populations. Environ Biol Fishes. 2001; 60(4):347–61. https://doi.org/10.1023/A:1011036127663
Koskinen MT, Piironen J, Primmer CR. Interpopulation genetic divergence in European grayling (Thymallus thymallus, Salmonidae) at a microgeographic scale: Implications for conservation. Conserv Genet. 2001; 2(2):133–43. https://doi.org/10.1023/A:1011814528664
Koskinen MT, Sundell P, Piironen J, Primmer CR. Spatiotemporal evolutionary relationships and genetic assessment of stocking effects in grayling (Thymallus thymallus, Salmonidae). Ecol Lett. 2002; 5(2):193–205. https://doi.org/10.1046/j.1461-0248.2002.00302.x
Mantel N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967; 27(2):209–20.
Peakall R, Smouse PE. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research – An update. Bioinformatics. 2012; 28(19):2537–39. https://doi.org/10.1093/bioinformatics/bts460
Pereira LHG, Foresti F, Oliveira C. Genetic structure of the migratory catfish Pseudoplatystoma corruscans (Siluriformes: Pimelodidae) suggests homing behaviour. Ecol Freshw Fish. 2009; 18(2):215–25. https://doi.org/10.1111/j.1600-0633.2008.00338.x
Perini VR, Paschoalini AL, Cruz CKF, Rocha RCGA, Senhorini JA, Ribeiro DM, Formagio PS, Bazzoli N, Rizzo E. Profiles of sex steroids, fecundity and spawning of a migratory characiform fish from the Paraguay–Paraná basin: A comparative study in a three-river system. Fish Physiol Biochem. 2013; 39(6):1473–84. https://doi.org/10.1007/s10695-013-9800-z
Petrere Jr M. Fisheries in large tropical reservoirs in South America. Lakes Reserv. 1996; 2(1–2):111–33. https://doi.org/10.1111/j.1440-1770.1996.tb00054.x
Piorski NM, Sanches A, Carvalho-Costa LF, Hatanaka T, Carrillo-Avila M, Freitas PD, Galetti Jr. PM. Contribution of conservation genetics in assessing neotropical freshwater fish biodiversity. Braz J Biol. 2008; 68(4 Suppl.):1039–50. https://doi.org/10.1590/S1519-69842008000500011
Piry S, Luikart G, Cornuet J-M. Computer note. BOTTLENECK: A computer program for detecting recent reductions in the effective size using allele frequency data. J Hered. 1999; 90(4):502–03. https://doi.org/10.1093/jhered/90.4.502
Primmer CR, Veselov AJ, Zubchenko A, Poututkin A, Bakhmet I, Koskinen MT. Isolation by distance within a river system: Genetic population structuring of Atlantic salmon, Salmo salar, in tributaries of the Varzuga River in northwest Russia. Mol Ecol. 2006; 15(3):653–66. https://doi.org/10.1111/j.1365-294X.2005.02844.x
Pritchard JK, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data. Genetics. 2000; 155(2):945–59. https://doi.org/10.1093/genetics/155.2.945
Ramella MS, Kroth MA, Meurer S, Nuñer APO, Zaniboni Filho E, Arisi ACM. Genetic variability in four fish species (Pimelodus maculatus, Prochilodus lineatus, Salminus brasiliensis and Steindachneridion scripta) from Uruguay River basin. Braz Arch Biol Technol. 2006; 49(4):589–98. https://doi.org/10.1590/S1516-89132006000500008
Rice WR. Analyzing tables of statistical tests. Evolution. 1989; 43(1):223–25. https://doi.org/10.2307/2409177
Rousset F. Genepop’007: A complete re-implementation of the Genepop software for Windows and Linux. Mol Ecol Resour. 2008; 8(1):103–06. https://doi.org/10.1111/j.1471-8286.2007.01931.x
Rueda EC, Carriquiriborde P, Monzón AM, Somoza GM, Ortí G. Seasonal variation in genetic population structure of sábalo (Prochilodus lineatus) in the Lower Uruguay River. Genetica. 2013; 141(7):401–07. https://doi.org/10.1007/s10709-013-9739-0
Sanches A, Galetti Jr. PM. Genetic evidence of population structuring in the neotropical freshwater fish Brycon hilarii (Valenciennes, 1850). Braz J Biol. 2007; 67(4 Suppl.):889–95. https://doi.org/10.1590/S1519-69842007000500012
Sanches A, Galetti Jr. PM, Galzerani F, Derazo J, Cutilak-Bianchi B, Hatanaka T. Genetic population structure of two migratory freshwater fish species (Brycon orthotaenia and Prochilodus argenteus) from the São Francisco River in Brazil and its significance for conservation. Lat Am J Aquat Res. 2012; 40(1):177–86. https://doi.org/10.3856/vol40-issue1-fulltext-17
Sivasundar A, Bermingham E, Ortí G. Population structure and biogeography of migratory freshwater fishes (Prochilodus: Characiformes) in major South American rivers. Mol Ecol. 2001; 10(2):407–17. https://doi.org/10.1046/j.1365-294X.2001.01194.x
Van Oosterhout C, Hutchinson WF, Wills DPM, Shipley P. Micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol Ecol Notes. 2004; 4(3):535–38. https://doi.org/10.1111/j.1471-8286.2004.00684.x
Wasko AP, Galetti Jr. PM. RAPD analysis in the Neotropical fish Brycon lundii: Genetic diversity and its implications for the conservation of the species. Hydrobiologia. 2002; 474(1):131–37. https://doi.org/10.1023/A:1016569919615
Yamamoto S, Morita K, Koizumi I, Maekawa K. Genetic differentiation of white-spotted charr (Salvelinus leucomaenis) populations after habitat fragmentation: Spatial-temporal changes in gene frequencies. Conserv Genet. 2004; 5(4):529–38. https://doi.org/10.1023/B:COGE.0000041029.38961.a0
Yazbeck GM, Kalapothakis E. Isolation and characterization of microsatellite DNA in the piracema fish Prochilodus lineatus (Characiformes). Genet Mol Res. 2007; 6(4):1026–34.
Authors

![]() Violeta R. Perini2,
Violeta R. Perini2, ![]() Alessandro L. Paschoalini1,
Alessandro L. Paschoalini1, ![]() Nilo Bazzoli1,
Nilo Bazzoli1, ![]() Elizete Rizzo2 and
Elizete Rizzo2 and ![]() Daniel C. Carvalho1
Daniel C. Carvalho1 ![]()
[1] Programa de Pós-graduação em Biologia de Vertebrados, Pontifícia Universidade Católica de Minas Gerais, Av. Dom José Gaspar, 500, 30535-610 Belo Horizonte, MG, Brazil. (NB) bazzoli@puc.minas.br, (ALP) alessandrobh@gmail.com, (DC)
carvalho.lgc@gmail.com (corresponding author).
[2] Departamento de Morfologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Av. Antônio Carlos, 6627, 30161-970 Belo Horizonte, MG, Brazil. (ER) ictio@icb.ufmg.br, (VRP) violetaperini@gmail.com.
Authors Contribution
Alessandro Loureiro Paschoalini: Investigation, Methodology.
Nilo Bazzoli: Project administration.
Elizete Rizzo: Conceptualization, Data curation, Project administration.
Daniel Carvalho: Conceptualization, Data curation, Formal analysis, Investigation, Project administration.
Ethical Statement
This research was approved by the Ethics Committee in Animal Experimentation from the Universidade Federal de Minas Gerais (Protocol number: 92/2010). Sampling was conducted with the authorization of the government agency SISBIO (License number: 22501–1/2009).
Competing Interests
The authors declare no competing interests.
How to cite this article
Perini VR, Paschoalini AL, Bazzoli N, Rizzo E, Carvalho D. Metapopulation dynamics of the migratory fish Prochilodus lineatus (Characiformes: Prochilodontidae) in a lotic remnant of the Grande River, Southeastern Brazil. Neotrop Ichthyol. 2021; 19(4):e200046. https://doi.org/10.1590/1982-0224-2020-0046
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2021 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted June 30, 2021 by Fernando Pelicice
Accepted June 30, 2021 by Fernando Pelicice
![]() Submitted August 27, 2020
Submitted August 27, 2020
![]() Epub November 01, 2021
Epub November 01, 2021