![]() Marina Tagliaferro1
Marina Tagliaferro1 ![]() ,
, ![]() Sean P. Kelly2 and
Sean P. Kelly2 and ![]() Miguel Pascual3
Miguel Pascual3
PDF: EN XML: EN | Supplementary: S1 | Cite this article
Abstract
The aim of this study was to determine the food webs structure of a large Patagonian river in two river sections (Upstream and Midstream) and to evaluate isotopic overlap between native and introduced species. We used stable isotope analyses of δ15N and δ13C and stomach content. The Upstream section had a more complex food webs structure with a greater richness of macroinvertebrates and fish species than Midstream. Upstream basal resources were dominated by filamentous algae. Lake Trout were found to have a higher trophic position than all other fish species in that area although, the most abundant fish species, were Rainbow Trout. Depending on the life stage, Rainbow Trout shifted from prey to competitor/predator. In the Midstream section, the base of the food webs was dominated by coarse particulate organic matter, and adult Rainbow Trout had the highest trophic level. Isotopic values changed among macroinvertebrates and fish for both areas. The two most abundant native and invasive species — Puyen and Rainbow Trout — showed an isotopic separation in Midstream but did not in Upstream areas. The presence of invasive fish that occupy top trophic levels can have a significant impact on native fish populations that have great ecological importance in the region.
Keywords: Diet; food webs; Mixing models; Salmonids; Stable isotopes analysis
El objetivo de este estudio fue determinar la estructura trófica de un gran río de la Patagonia en dos secciones (río arriba y medio) y evaluar la superposición isotópica entre especies nativas e introducidas. Utilizamos análisis de isótopos estables δ15N y δ13C y contenido estomacal. La sección río arriba tuvo una estructura de trama trófica más compleja, con mayor riqueza de macroinvertebrados y peces respecto de la sección media. Los recursos basales dominantes río arriba fueron las algas filamentosas. En esta área, la trucha de lago tuvo la posición trófica más alta entre los peces, aunque, las especies de peces más abundantes fueron las truchas arcoiris. Dependiendo del estadio, la trucha arcoiris cambió su rol de presa a competidor/depredador. En la sección media del río, la base de la trama trófica estuvo dominada por materia orgánica particulada gruesa y la trucha arcoíris adulta fue el depredador tope. Los valores isotópicos variaron entre zonas para invetebrados y peces. Las dos especies nativas e invasoras más abundantes, Puyen y trucha arcoiris, mostraron una separación isotópica en la sección media, pero no en secciones de río arriba. La presencia de peces invasores que ocupan una posición tope en los niveles tróficos puede tener un impacto significativo sobre las poblaciones de peces nativos de gran importancia ecológica en la región.
Palabras chave: Análisis de Isótopos estables; Dieta; Modelos de mezcla; Salmónidos; Trama trófica
Introduction
Species invasion and habitat degradation represent major threats to biodiversity (Vitousek, 1990; Clavero, García-Berthou, 2005; Gallardo et al., 2016). Invasive species can alter fundamental ecological characteristics, such as species composition, the cycling of nutrients and overall ecosystem productivity (Mack et al., 2000; Muñoz et al., 2009; Buoro et al., 2016). The introduction of non-native fish species is considered one of the most significantly negative anthropogenic impacts to aquatic ecosystems (Power et al., 1996; Fausch, 2007; Simberloff et al., 2013). As dominant predators, fish can have an important effect at the individual, population, community, and ecosystem level (Simon, Townsend, 2003; Tagliaferro et al., 2014a; Buoro et al., 2016).
At the community level, food webs can experience different effects from introduced species, both in structural and functional characteristics (Townsend, 2003). Changes in the ecological structure of communities by introducing species can directly modify the flow of energy and matter in the ecosystem (de Ruiter et al., 1995; Chapin III et al., 2000) or indirectly modify abundance or species traits (Power et al., 1996; Milardi et al., 2016). In particular, top predators such as salmonids can alter trophic relationships through cascading effects (Power, 1992; Milardi et al., 2016; Herrera-Martínez et al., 2017). A reduction in native top predators could generate an increase in prey populations and deplete basal resources, generating a cascade of ecological effects (Chapin III et al., 2000; Shelton et al., 2016). Furthermore, a reduction in native species or the presence of non-native assemblages can produce novel species interactions that have not co-evolved (Hobbs et al., 2006; Tagliaferro et al., 2014a) and could lead to a system with unstable trophic characteristics (Vander Zanden et al., 1999; Cross et al., 2013).
In the Southern Hemisphere, introductions of salmonids were promoted since the beginning of the 20th century by the governments of Chile (Basulto del Campo, 2003), New Zealand (Flecker, Townsend, 1994; McDowall, 2003), and Argentina (Tulian, 1908). Most introductions included anadromous (Atlantic, Chinook, Coho and, Sockeye Salmon), partially migratory and resident species (Rainbow Trout), facultative anadromous species (Rainbow and Brown Trout), and freshwater resident species (Brook and Lake Trout) (Pascual et al., 2002; Pascual, Ciancio, 2007). These species were introduced in ecosystems where the fish fauna had originally consisted of small and unique assemblages of native species (Dyer, 2000; López et al., 2003) without prior evaluation of ecological and trophic consequences. Many studies have shown that the introduction of salmonids has been widely implicated in the reduction of different prey species and native biota (Crowl et al., 1992; Greig, McIntosh, 2006; Cussac et al., 2020), reduction of niche breadth in native Galaxiid fish (Townsend, 2003; McHugh et al., 2012), habitat segregation of Galaxiid populations (Penaluna et al., 2009; Cussac et al., 2020), and dietary overlap between native fish species and introduced salmonids (Di Prinzio, Casaux, 2012; Tagliaferro et al., 2014a). However, variations in the impacts on the system would depend on the particular characteristics of the invasive taxa (Arismendi et al., 2014).
One of the main problems of these introductions was the establishment of natural populations of several of these species across freshwater ecosystems in South America (Arismendi et al., 2019). Rainbow Trout Oncorhynchus mykiss (Walbaum, 1792) and Brown Trout Salmo trutta Linnaeus, 1758 were widely spread and became the most abundant species (Pascual et al., 2002), followed by Chinook Salmon Oncorhynchus tshawytscha (Walbaum, 1792) (Ciancio et al., 2005; Becker et al., 2007; Correa, Gross, 2008) and Lake Trout Salvelinus namaycush (Walbaum, 1792) (Arismendi et al., 2014; Tagliaferro, 2014). Although salmonids have been classified as visual and opportunistic predators (Elliott, 1973; Hansen et al., 2013), their diet widely changes between species and ontogeny (De Crespin De Billy, Usseglio-Polatera, 2002). For instance, studies in adults indicated that salmonids feed heavily on Galaxiids and silversides (Vila et al., 1999; Macchi, 2004; Alvear et al., 2007), with large Brown Trout being more piscivorous than Rainbow Trout (Pascual et al., 2007). However, both Rainbow and Brown Trout feed on macroinvertebrates during the first year of life (Tagliaferro et al., 2014a). Meanwhile, Chinook Salmon are primarily piscivorous with Galaxiids as the most common prey (Soto et al., 2003; Arismendi et al., 2009) as well as Lake Trout, that although they might feed on macroinvertebrates, it is still considered an apex piscivore (Post et al., 2000; Tronstad et al., 2010; Syslo et al., 2016).
Traditionally, food webs interactions have been studied utilizing stomach content analyses (SCA) and exclusion/forced interaction experiments. Currently, stable isotope analyses (SIA) complement these methodologies because it provides continuous measurements of trophic position and energy flow (DeNiro, Epstein, 1978; Caut et al., 2009; Nielsen et al., 2015). Therefore, SIA provides a robust tool to test theories of trophic connections (Post et al., 2000; Post, 2002) and to evaluate effects of species invasions on trophic structures (Vander Zanden et al., 1999; Collins et al., 2016). This is especially useful for estimating the trophic position of species with diets difficult to quantify (Kling et al., 1992; Bowes, Thorp, 2015). Recently isotopic ratios of carbon (δ13C) and nitrogen (δ15N) have been utilized in determining the marine diet of introduced salmonids (Ciancio et al., 2008[), characterizing food webs of shallow lakes (Lancelotti et al., 2010) and documenting trophic shifts between invasive salmonid and native Galaxiid species in lakes (Correa et al., 2012).
The aim of this study was to reconstruct the trophic relationships within aquatic food webs of the Santa Cruz River using SIA and SCA. This is especially important not only because of the need to identify the impacts of invasive species but also because of imminent changes associated with the construction of dams along glacial rivers in Patagonia, which could also impact aquatic food webs in the region. This research will be the first study of food webs in the Santa Cruz River (the second largest river of Patagonia). This river is a large glacial river, with low human impact, that has a predictable flood pulse with a stable discharge, distinct seasonal cycles, and a high sediment load (Tagliaferro et al., 2013). Besides the interest of knowing how communities are formed in this understudied system, there were two main objectives: 1) determine differences in the food webs structure in two river sections with different habitat structure, 2) evaluate if there are overlaps of isotopic signatures among native species and introduced salmonids. Our hypotheses were: (H1) a more complex food webs will be suitable in upstream sections, (H2) the two most abundant species, Rainbow Trout and the native Galaxiid will experience a different diet and isotopic overlap between the two sections. Since upstream sections represent more suitable environment for Lake Trout and native Perch (Otturi et al., 2016; Arismendi et al., 2019), are widely used for anadromous Rainbow Trout (Liberoff et al., 2015), and have a greater amount of biomass of macroinvertebrates and producers (Tagliaferro et al., 2013). We predict a more diverse food webs in this section; moreover, we predict that diet and isotopic overlap between the two sections we selected will depend on the presence of other salmonids and macroinvertebrate abundance. This study provides evidence for how introduced fish species can significantly alter food webs interactions. Understanding the impacts of introduced species should lead to better management practices that result in greater conservation efforts for native fish populations in these understudied ecosystems.
Material and methods
Study area. The Santa Cruz River (50°14’S, 71°58’W to 50°07’S, 68°20’W) is in one of the least studied areas of Argentina. It originates in two oligotrophic to ultra-oligotrophic large glacial lakes, Viedma and Argentino, and flows uninterrupted for 382 km across the Patagonian plateau to drain into the Atlantic Ocean (Fig. 1; Brunet et al., 2005). The river has an average discharge of 691 m3 s-1 (min. 278.1 m3 s-1 in September and max. 1,278 m3 s-1 in March), which is highly predictable due to a glacially dominated regime (Tagliaferro et al., 2013). The mean water temperature is 9 °C with a maximum registered in January (15° C) and a minimum in July (3° C). The sampling sites were located in two river sections: Upstream (50°10’S, 69°55’W, an area which contains gravel bars and sediment deposits) and Midstream (50°09’S, 69°59’W, where the river runs through a natural canyon). Downstream areas were not included to avoid the marine influence in trophic webs and possible urban effects. Whereas temperature, slope, dissolved oxygen were homogeneous at large scales, the two studied sections present different characteristics at the local scale in chlorophyll-a concentrations, inorganic matter, particles substrate size, and depth (S1; Tagliaferro et al., 2013).
In relation to biological characteristics, the Upstream section was previously characterized as areas with higher macroinvertebrate abundance, richness and higher chlorophyll-a biomass. Whereas the Midstream section was associated with lower macroinvertebrate richness and abundance (Tagliaferro et al., 2013; Tagliaferro, Pascual, 2017). Fish assemblages in the Santa Cruz River contain populations of native Perch Percichthys trucha (Valenciennes, 1833) (Percichthydae), Large or Big Puyen Galaxias platei Steindachner, 1898 and Puyen G. maculatus (Jenyns, 1842) (Galaxiidae), the latter being the most abundant native species (Tagliaferro et al., 2014b). Among the exotic species, the most abundant are Rainbow Trout Oncorhynchus mykiss. Other introduced salmonids include Brown Trout Salmo trutta, Lake Trout Salvelinus namaycush and Chinook Salmon O. tshawytscha.

FIGURE 1| Sampling areas in the Santa Cruz River, Argentina. Upstream area corresponds to the locally known “Labyrinth”, and Midstream area correspond to “Estancia San Ramon”. Map created by the authors, upper picture taken from Google Earth (R).
Sampling design. Sampling was done in April 2010 (during average discharge condition of the Santa Cruz River) since (1) large glacial rivers in general experience a high flow during the summer (January-February in Southern hemisphere) due to ice melting, (2) to avoid the spawning period for Rainbow and Brown Trout (around September) in the Santa Cruz River (Riva Rossi et al., 2003). It is important to avoid taking samples for SIA between August-March since during the first month these two adult species are not feeding, and there would be a bias in the stomach content of adults. On the other hand, young of the year (YOY) juveniles can use maternal resources for few months (Liberoff et al., 2013), and the isotopic signal might get confusing results due to maternal effects. Finally, macroinvertebrates tend to experience changes in distribution due to temporal effects. Thus, we selected a mid-flow period which is the most representative scenario with YOY and adult trout feeding, and mid to high macroinvertebrate abundance.
Sampling in the Santa Cruz River included different components of the aquatic community: fish, macroinvertebrates, and basal resources in two distinct areas related to river morphology. Benthic producers (i.e., macrophyte and algae) were estimated by the mean value of three individual visual evaluations of a 10m long transect along the river. Benthic algae were obtained by scraping rocks (n=9 and n=3 for Upstream and Midstream sections, respectively), whereas planktonic algae (n=3 for each section) were collected by filtering river water using a plankton net (15 μm pore-size). Both samples of algae were filtered using sterile glass fiber filters. Macrophytes were cut from the riverside and packed in airtight plastic bags (n=3 for each section). Debris samples were taken from macrophytes cover areas. Four to nine benthic macroinvertebrate samples were obtained at each river section with a kick-net of 450 μm mesh covering 0.25 m2. Algae, macrophytes, and macroinvertebrate samples were stored in a portable cooler at -18°C in the field. Algae samples were stored in glass fiber filters inside individual aluminum envelopes. Macroinvertebrates were stored in plastic 500ml containers and once in the laboratory were separated and identified to the lowest possible taxonomic level following descriptions from Domínguez, Fernández (2009). Macroinvertebrates were then grouped according to functional feeding group (FFG) (Merrit, Cummins, 1996). Both macroinvertebrates and aquatic plants were dried for 24 h at 60° C. The most abundant macroinvertebrates, along those with sufficient biomass were used for SIA.
Small fish (i.e., total length range: 50 to 140 mm) were caught by using standard three-pass electrofishing methods along 100 m transects at each site from the littoral zone to depths of 0.6 m (Jones, Stockwell, 1995; Meador et al., 2003) using a Smith-Root LR-24 electrofisher set to a frequency of 90 Hz and a pulse width of 3 m/s. This data was then used as an indirect measurement of abundance (CPUE). Due to the morphology of the river and water velocity at the time of the study, as well as following work safety protocols, the sampling was restricted to a narrow width of the main stem of the river. Larger fish (length range > 180 mm) were captured by using gillnets of 15, 20, 30, 50, 60 mm. Captures were related to gillnet effort (CPUE). All fish were measured for total length with a digital caliper (0.01mm nearest unit) and weighed on a Mettler PC 440 Delta Range balance (0.003 g nearest unit). A portion of the posterior dorsal muscle was excised and dried at 60°C. Fish stomach contents were removed and stored in 70° ethanol for further separation and identification using the same procedure previously mentioned for macroinvertebrate samples.
Once dried, all samples were ground into a homogeneous powder using a hand mortar and pestle. Three replicates of macroinvertebrate and aquatic plants were used for stable isotopes analyses. In each stream area (i.e., Upstream and Midstream), we used replicate samples for Puyen (n=8-10), Chinook Salmon (n=4), and Rainbow Trout (n=18, n=6 for each life stage). In Upstream areas, we analyzed replicate samples for Brown Trout (n=3), Perch (n=4), Big Puyen (n=3) and Lake Trout (n=3). A subsample of each individual or group of individuals in case of very small species (e.g., chironomids) was weighed on a precision balance Shimadzu (error 0.001 mg), placed in a tin capsule for further analysis at the Stable Isotopes laboratory at the University of California, Davis: 2-3 mg in the case of plants and 1 ± 0.2 mg samples for animal tissue. Samples were analyzed for 13C and 15N isotopes using an elemental analyzer PDZ Europa ANCA-GSL interface with a mass spectrometer PDZ Europa 20-20 isotope ratio (Sercon Ltd., Cheshire, UK). The long-term standard deviation of these analyses was 0.2 ‰ to 0.3 ‰ for 13C to 15N. The stable isotope ratios are expressed as δ values of ‰: δX = 103 [(Rsample Rstandard -1)-1], where X is 13C or 15N and R is the corresponding ratio 13C:12C or 15N:14N. The values of final “δX” were expressed relative to international standards V-PDB (Vienna PeeDee Belemnite) and N2 from air for carbon and nitrogen, respectively.
Data Analysis. A two-way PERMANOVA test was performed using the statistical program PAST (version 3.14.) to evaluate possible differences in isotopic values of carbon and nitrogen between the two selected areas of the river for the two most abundant fish species (Rainbow Trout and Puyen) and dominant macroinvertebrate FFGs. For the most abundant species, a one-way PERMANOVA was performed to evaluate possible local differences.

Isotopic fractionation values for Rainbow Trout were ∆13C 1.9 ± 0.5 and ∆ 15N 3.2 ± 0.2 (McCutchan Jr et al., 2003), and ∆13C 1.6 ± 0.5 and ∆15N 3.5±0.7 were applied for macroinvertebrates and plants (DeNiro, Epstein, 1980; Rounick, Hicks, 1985; McCutchan Jr et al., 2003). The trophic position was calculated for fish and macroinvertebrates using the isotopic variation in nitrogen (Post, 2002) and possible variants of fractionation as follow:
where TP indicates the trophic position, λ represents the trophic position of the prey (possible prey items from diet), δ15Nconsumer, are the stable isotope ratios of the organism of which the trophic position is being calculated and δ15Nbase is the ratio for primary producers. Finally, ∆n indicates the fractionation in 15N between the consumer and its diet. The baseline for each trophic position in each stream zone was estimated using mean value of possible primary producers considering the fractionation factors (DeNiro, Epstein, 1980; Rounick, Hicks, 1985; McCutchan Jr et al., 2003). In the Midstream section only Debris and Debris associated to Myriophyllum sp. were used to calculate trophic positions of preys since the fractionation did not exceed the ∆13C 1.6 ± 0.5 and ∆15N 3.5±0.7; algae were not used since there was a ∆13C >15. Similarly, in Upstream sections, the macrophytes and the planktonic algae and Nostoc sp. were excluded from the estimation.
A total of 432 stomach contents of fish were analyzed in terms of biomass to evaluate the contribution of prey to diet. After the selection of possible isotopic sources according to SCA, Bayesian isotopic mixing models were applied by using V4.0 SIAR (Stable Isotope Analysis in R) (Parnell et al., 2010) using R software (R -version 3.2.5 2016) to assess the relative contributions of prey to the diet of fish. SIAR mixing model results were calculated with credibility intervals of 5, 25, 75 and 95%.
Results
General Pattern. Basal resources were represented by macrophytes (mainly Myriophyllum sp.) and algae (mainly Cladophora sp., but also Nostoc sp. and Batrachospermum sp.) (Tab. 1). Debris was constituted by dead macrophytes and Coiron sp. grasses. Both macrophytes and benthic visual algae cover were very low along the two sections (< 1.5-3% and < 4-5%, respectively), with algae patches being in the Upstream section and macrophytes in the Midstream section. Macroinvertebrate FFGs included scraper-grazers, shredders, filterer-collectors, collector-gatherers, and predators. Most abundant FFGs in Upstream areas were scraper-grazers (47.5 ± 22.9%), filterer-collectors (24.3 ± 33.0%), and shredders (19.7 ± 6.0%); in Midstream areas shredders were the most abundant FFG (41.0 ± 4.4%), followed by scraper-grazers (33.7 ± 12.6%) and collector-gatherers (17.4 ± 22.6%). Fish taxa in Upstream areas were dominated by top predators, including Lake Trout, Brown Trout, Rainbow Trout, Chinook Salmon, Perch, Puyen and Big Puyen, with Rainbow Trout being the most abundant species (Tab. 2). Moreover, different ontogenetic stages of Rainbow Trout were captured (yearling, juveniles and adults). In Midstream areas only four fish species were captured: Rainbow Trout (different ontogenetic stages), Chinook Salmon (ocean type), Perch and Puyen (Tab. 2), with Puyen being the most abundant species.
TABLE 1 | Primary producer cover (%) along a sampling line of 10 m and macroinvertebrates abundance in each kick-net sample (0.25 m2).
Upstream | Midstream | |
Primary
producers (% cover) | ||
Filamentous algae
(fil) | 0.8 ± 0.8 | < 0.1 |
Cladophora algae (Ch), | 17.1 ± 6.9 | < 0.1 |
Nostoc sp. | < 0.1 | < 0.1 |
Myriophyllum (Plant-My) | 1.2 ± 0.9 | 5.7 ± 4.9 |
Debris | < 0.1 | 1.4 ± 1.0 |
Macroinvertebrates | ||
Shredder | 3.7 ± 1.0 | 0.9 ± 0.9 |
Scraper/grazer | 15.5 ± 1.8 | 9.3 ± 2.7 |
Collector-Gatherer | 6.3 ± 1.3 | 15.8 ± 11.2 |
Collector-filterer | 3.8 ± 1.3 | 2.6 ± 1.7 |
Predator | 5.8 ± 1.0 | 2.1 ± 0.9 |
TABLE 2 | Fish captures along the studied areas in the Santa Cruz River. YRT refers to Yearling Rainbow Trout, JRT to juvenile Rainbow Trout, and ART to Adult Rainbow Trout. Puyen and YRT captures were related to the three pass electrofishing method and the rest of the fish species and stages were related to the use of gillnets.
Proportion
of captures (%) | ||
Upstream | Midstream | |
Lake Trout | 1.4 | 0 |
Brown Trout | 1.9 | 0 |
YRT | 42.1 | 13.9 |
JRT | 21.5 | 21.3 |
ART | 15.8 | 1.9 |
Chinook | 2.4 | 1.9 |
Big Puyen | 1.4 | 0 |
Puyen | 10.0 | 58.3 |
Perch | 3.3 | 2.8 |
Stomach Content Analyses. Stomach contents for small fish (Puyen and yearling Rainbow Trout) were composed nearly entirely of benthic macroinvertebrates, mainly shredders and collector-filterers, with less than 2% being attributed to terrestrial inputs (Tab. 3). Juvenile and adult Rainbow Trout (in both river areas) were found to consume Puyen, along with macroinvertebrates of different FFGs (Tab. 3). Brown Trout, Chinook Salmon, and Perch consumed juvenile Trout. Lake Trout fed exclusively on fish of any size including both Trout and Puyen species (Tab. 3).
TABLE 3 | Stomach content of fish species found along the Santa Cruz River. Relative contribution (% range) of collector-filterer, grazer, shredder and scraper benthic macroinvertebrates (functional feeding groups), predator invertebrates and fish items.
Lake Trout | Brown Trout | Rainbow yearling trout | Rainbow juvenile trout | Rainbow adult trout | Chinook Salmon | Puyen | Perch | Big/Large Puyen | |
Fish | 98-100 | 20-35 | 0-1 | 2-4 | 35-48 | 18-35 | 10-19 | 0-1 | |
Invertebrate
predator | 2-6 | 6-11 | 28-35 | 14-21 | 9-13 | 32-48 | 0-3 | 28-37 | |
Collector-filterer | 24-32 | 32-46 | 3-11 | 1-4 | 2-8 | 4-9 | 9-19 | 1-5 | |
Grazer | 4-7 | 16-22 | 27-39 | 7-14 | 0-1 | 0-2 | 46-65 | 47-69 | |
Shredder-scraper | 0-2 | 38-44 | 32-37 | 25-37 | 21-33 | 62-71 | 59-72 | 15-22 | 4-8 |
Stable Isotope Analyses. Biplots for carbon (δ13C) and nitrogen (δ15N) showed a clear pattern for an autotrophic base of δ13C values (Fig. 2). The Midstream area showed a narrower range at the base of the trophic web with debris (CPOM) and parts of Myriophyllum sp. being the primary basal sources. The Upstream area showed a wider range of basal sources with several different species of algae (mainly Cladophora sp. and Batrachospermum sp.) (Fig. 2A, B). Isotopic values also showed a grouping of herbivorous macroinvertebrates enriched in 15N, and a grouping of fish enriched in both 13C and 15N (Fig. 2A, B). Isotopic values in Midstream areas (Fig. 2A) tended to be enriched in 15N for all groups in comparison with Upstream areas (Fig. 2B). Although the general pattern of isotopic composition was similar for both study areas, there were statistically significant differences between sites in δ15N and δ13C values (Two-ways PERMANOVA, Friver area = 4.361; p= 0.007; Fspecies =41.329; p=0.0001). Among the most abundant fish species, Puyen showed significant differences in isotopic signature between Upstream and Midstream areas (p= 0.0001). Due to the presence of different ontogenetic stages of Rainbow Trout, the isotopic signature was analyzed separately, and differences were found depending on life stage. Only juvenile Rainbow Trout of the first year showed significant differences between Mid and Upstream areas (p=0.0001). Adult Rainbow Trout and older juveniles showed no significant differences (p=0.12, and p=0.834, respectively).
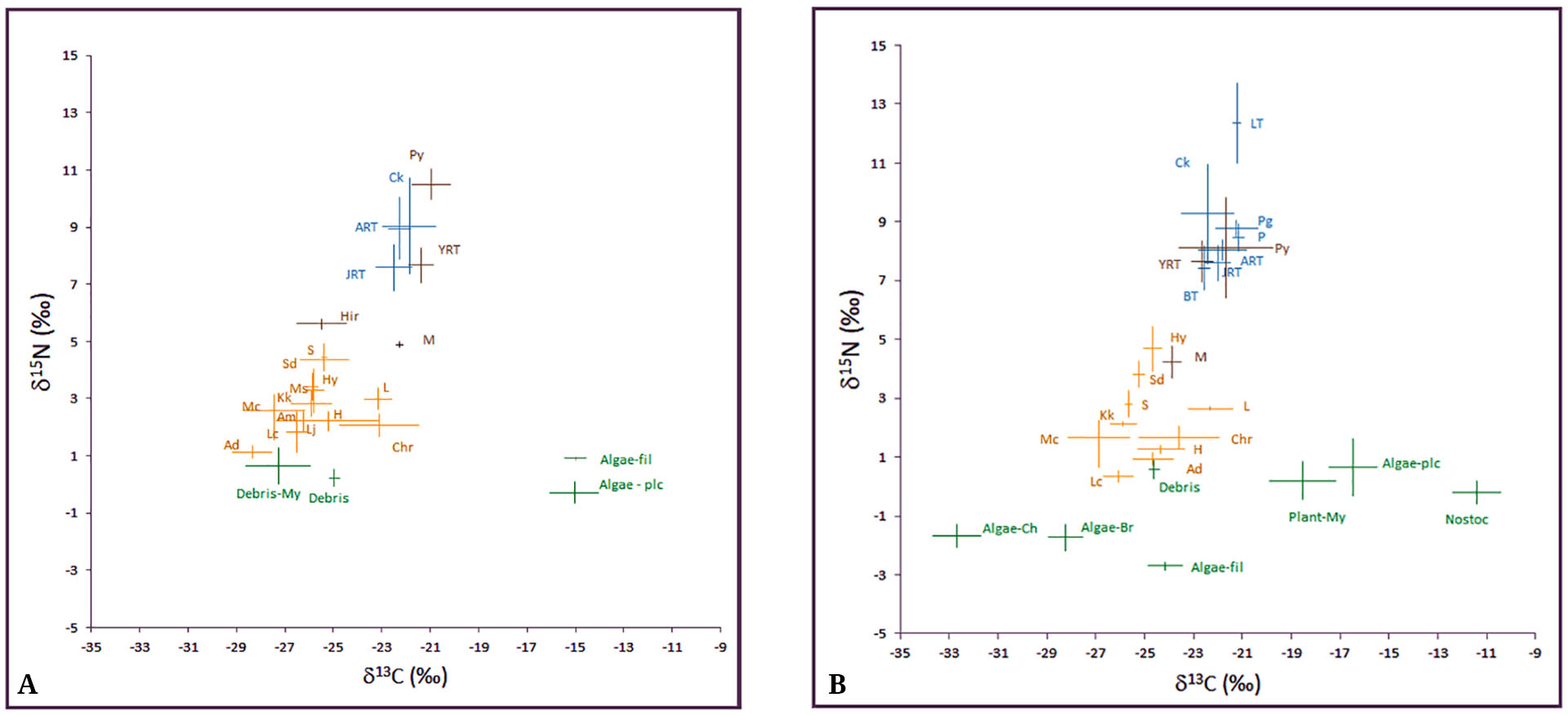
FIGURE 2| Values of δ15N and δ13C found in the Midstream areas (A) and Upstream areas (B). Error bars correspond to standard deviation. Abreviations: Hirudinea (Hir), Muscidae (M), Simuliidae (S), Smicridea dythira (Sd), Hydrobiosidae (Hy), Mastigoptila spp. (Ms), Klapopteryx kuscheli (Kk), Lymnaea (L), Meridialaris chiloeensis (Mc), Antarctoperla michaelseni (Am), Hyalella sp. (H), Luchoelmis cekalovici (Lc), Limnoperla jaffuelli (Lj), Andesiops sp. (Ad), Chironomidae (Chr), Filamentous algae (fil), Bratrachospermun sp. algae (Br), planktonic algae (plc), Debris associated to Myriophyllum sp. (My), Cladophora algae (Ch), macrophyta of genus Myriophyllum (plant-My). Colors indicate primary producers (green), herbivores (orange), non-piscivores predators (brown) and general predators (blue).
In the Upstream section, Lake Trout showed significantly higher δ15N values than the rest of the fish species. The most abundant species was Puyen, followed by Rainbow Trout. Although differences were found in isotopic signatures (F = 21.174, p=0.001), the “a-posteriori” comparisons showed no significant differences between Puyen and the rest of the fish species, except for Lake Trout and juvenile Rainbow Trout (Tab. 4). Rainbow Trout yearlings showed significant differences with Perch, Chinook Salmon, and Lake Trout. However, older juveniles (>1 year) were significantly different from Perch and Lake Trout, while adult specimens only differed from Lake Trout (Tab. 4). In Midstream areas significant differences in isotopic signature were found (F= 87.185, p=0.001). Puyen was the most abundant fish species and showed significant differences (a posteriori test) with all Rainbow Trout ontogenetic stages (Tab. 5). Among Rainbow Trout, ontogenetic stages differed between yearling Rainbow Trout and juveniles and adults, but no significant differences were found between juveniles and adult specimens (Tab. 5). Perch had low abundances and showed no significant differences in isotopic values with Puyen or Rainbow Trout (Tab. 5).
Juvenile and adult Rainbow Trout showed no significant differences in isotopic values between river sections, but there were significant differences with yearling stages between Mid and Upstream areas (F= 5.201, p=0.012). However, in the SCA, the contribution of different prey biomass in the diet of Rainbow Trout showed differences between ontogenetic stages and river areas for the period under study (Tab. 3). When comparing the contribution of prey to stomach contents and the possible sources for the mixing model, it was not possible to create a virtual polygon of resources that included the predator, and therefore mixing models were not employed for yearling Rainbow Trout.
TABLE 4 | PERMANOVA analysis results of Upstream area and pairwise comparisons. CF= collector-filterer, CG= collector-gatherer, SCR=scrapers, SHR= shredders, PRED= predator, L. Trout= Lake Trout, B. Trout= Brown Trout, R. Trout= Rainbow Trout.
Pairwise | CF | CG | SCR | SHR | PRED | Puyen | Perch | Chinook | Lake | Brown | A R. Trout | J R. Trout |
CG | 0.274 | |||||||||||
SCR | 0.009 | 0.001 | ||||||||||
SHR | 0.011 | 0.447 | 0.002 | |||||||||
PRED | 0.009 | 0.002 | 0.008 | 0.005 | ||||||||
Puyen | 0.000 | 0.000 | 0.001 | 0.000 | 0.002 | |||||||
Perch | 0.003 | 0.001 | 0.007 | 0.002 | 0.009 | 0.681 | ||||||
Chinook | 0.003 | 0.001 | 0.008 | 0.002 | 0.016 | 0.301 | 0.173 | |||||
L. Trout | 0.003 | 0.001 | 0.007 | 0.002 | 0.008 | 0.000 | 0.007 | 0.008 | ||||
B. Trout | 0.004 | 0.010 | 0.007 | 0.008 | 0.096 | 0.308 | 0.024 | 0.109 | 0.009 | |||
A R.
Trout | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.381 | 0.084 | 0.107 | 0.001 | 0.161 | ||
J R.
Trout | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.673 | 0.030 | 0.159 | 0.000 | 0.076 | 0.505 | |
Y R.
Trout | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.366 | 0.003 | 0.027 | 0.000 | 0.057 | 0.106 | 0.628 |
TABLE 5 | PERMANOVA analysis results of Midstream area and pairwise comparisons. CF= collector-filterer, CG= collector-gatherer, SCR=scrapers, SHR= shredders, PRED= predator, R. Trout= Rainbow Trout.
Pairwise | CF | CG | SCR | SHR | PRED | Puyen | Perch | A R. Trout | J R. Trout |
CG | 0.000 | ||||||||
SCR | 0.009 | 0.001 | |||||||
SHR | 0.030 | 0.009 | 0.007 | ||||||
PRED | 0.236 | 0.000 | 0.032 | 0.004 | |||||
Puyen | 0.000 | 0.000 | 0.001 | 0.000 | 0.000 | ||||
Perch | 0.029 | 0.011 | 0.006 | 0.011 | 0.044 | 0.180 | |||
A R.
Trout | 0.000 | 0.000 | 0.001 | 0.000 | 0.000 | 0.000 | 0.112 | ||
J R.
Trout | 0.000 | 0.000 | 0.001 | 0.000 | 0.000 | 0.001 | 0.082 | 0.056 | |
Y R.
Trout | 0.000 | 0.000 | 0.001 | 0.000 | 0.000 | 0.038 | 0.061 | 0.038 | 0.038 |
Trophic Position. Trophic positions for yearling Rainbow Trout and Puyen were not included due to large differences in isotopic values of their prey, which could cause an incorrect positioning. Herbivores, independently of their FFG, were placed in a lower secondary trophic position and close to one as would be expected for their feeding habits (Tab. 6). Collector-filterers showed a greater isotopic enrichment value for δ15N, which resulted in a higher trophic level. The trophic position for predatory macroinvertebrates was based on other macroinvertebrate isotopic signals and the trophic level varied between 2.5 and 3 (Tab. 6). Exclusively piscivorous fish, such as Lake Trout, showed a trophic level of 4, while fish that had mixed diets of fish and macroinvertebrates were 2.5 (Tab. 6).
TABLE 6 | Trophic position for selected taxa in Upstream and Midstream areas. FFG: Functional feeding groups. Rainbow Trout includes both juvenile and adult individuals.
Study Area | |||
FFG | Taxa | Upstream | Midstream |
Scraper-grazer | Lymnaea sp. | 1.50-1.65 | 1.55-1.71 |
Scraper-grazer | Luchoelmis cekalovici | 1.13-1.24 | 1.20-1.36 |
Scraper-grazer | Andesiops sp. | 1.30-1.43 | 1.00-1.14 |
Scraper-grazer | Meridialaris chiloeensis | 1.50-1.65 | 1.46-1.62 |
Collector-gatherer | Hyalella sp. | 1.38-1.53 | 1.30-1.48 |
Collector-gatherer | Limnoperla jaffueli | – | 1.22-1.35 |
Collector-gatherer | Chironomidae | 1.39-1.60 | 1.30-1.45 |
Shredder | Antarctoperla michaelseni | – | 1.34-1.38 |
Shredder | Klapopteryx kuschelli | 1.60-1.80 | 1.50-1.67 |
Collector-filterer | Mastigoptila sp. | – | 1.70-1.85 |
Collector-filterer | Smicridea dithira | 2.10-2.33 | 1.95-2.18 |
Collector-filterer | Simuliidae spp. | 1.82-2.02 | 1.97-2.11 |
Predator | Glossiphonidae spp. | – | 2.80-3.19 |
Predator | Hydrobiosidae spp. | 2.74-3.03 | 2.23-2.52 |
Predator | Muscidae spp. | 2.52-2.90 | 2.68-2.97 |
Predator | Lake Trout | 4.24-4.60 | – |
Predator | Brown Trout | 2.89-4.58 | – |
Predator | Rainbow Trout | 2.65-3.42 | 1.97-3.22 |
Predator | Chinook Salmon | 3.33-4.60 | 2.57-3.03 |
Predator | Big Puyen | 3.18-3.40 | – |
Predator | Puyen | 3.24-3.72 | 2.92-3.51 |
Predator | Perch | 3.11-4.12 | 2.70-3.70 |
Mixing Models. Only the most abundant prey items found during the SCA were selected to be included as possible sources in the subsequent mixing models for calculating the proportion of each group to the diet of native and introduced fish species. The results of Chinook Salmon mixing models showed a high contribution (95% Confidence Interval: 95%-CI) of Puyen (43.4 ± 19.7%), followed by simuliid larvae (35.4 ± 20.5%), and amphipod Hyalella sp. (21.2 ± 18.6%). The estimated mixing model for adult Brown Trout diet showed a major contribution from Hyalella sp. (5%-CI= 29.5 ± 11.3%), Klapopteryx kuscheli (5%-CI 33.4 ± 15%), Puyen (5%-CI 15.8 ± 13%) and juvenile Rainbow Trout (5%-CI 21.3 ± 15.1%). The fit of the data for Lake Trout was concordant with the results of stomach contents, showing a comparable contribution from Puyen (5%-CI 46.6 ± 16.8%) and Rainbow Trout (5%-CI 53.4 ± 13.6%). Perch showed a dominantly piscivorous diet, mainly composed of Puyen and juvenile Trout, followed by a variety of predatory (e.g., Hydrobiosidae, Lancetes sp.) and herbivorous (e.g., Hyalella sp., Chironomidae) macroinvertebrates. Prey that fit the diet model for Perch were amphipods (37.5 ± 10.1%), juvenile Rainbow Trout (5%-CI 32.1 ± 11.6 %), and Puyen (5%-CI 30.4 ± 14.9%).
In the Midstream section, due to differences in diets seen in SCA of juvenile and adult Rainbow Trout, mixing models were performed using Lymnaea sp., Simuliidae spp. larvae and Puyen as sources for juvenile Trout; and for adult Trout, we included Puyen and the stoneflies K. kuscheli and Antarctoperla michaelseni. Juvenile Rainbow Trout showed a low contribution of Puyen in their diet (5%-CI 10.5 ± 8.5%); while Simuliidae spp. and gastropods contributed 49.5 ± 12% and 40 ± 23.1% (5%-CI), respectively. The model for adult Trout showed, unlike the Upstream areas, an important contribution of Puyen (5%-CI 39.2 ± 10.1%), followed by A. michaelseni (5%-CI 30.7 ± 22.5%) and K. kuscheli (5%-CI 30.1 ± 19.8%) (Fig. 3B).
In Upstream section, mixing models for Rainbow Trout showed Puyen, the stonefly nymph K. kuscheli, the gastropod Lymnaea sp., and caddisfly larvae Hydrobiosidae as possible resources. Mixing models for juvenile Rainbow Trout showed the contribution of K. kuscheli (5%-CI 49.7 ± 16.4%), followed by Puyen (5%-CI 27.4 ± 8.3%) and Lymnaea sp. (5%-CI 22.9 ± 16%) (Fig. 4A). The model for adult Rainbow Trout showed a similar contribution of Lymnaea sp. (5%-CI 23.9 ± 13.1%), K. kuscheli (25.4 ± 17.8%), the Hydrobiosidae spp. (5%-CI 26.9 ± 15.7%), and Puyen (5%-CI 25.1 ± 7.9%) (Fig. 4B).

FIGURE 3| Mixing models adjusted for juvenile and adult Rainbow Trout in Midstream areas. Herbivores are in orange and predators in brown color.

FIGURE 4| Mixing models adjusted for juvenile and adult Rainbow Trout in Upstream areas. Herbivores are in orange and predators in brown color.
Discussion
The present research is the first study of food webs in the Santa Cruz River, a river that is about to change due to damming without information regarding the trophic structure and with poor information about the influence of introduced species on aquatic food webs. Findings in this study support that native Puyen is more abundant in Midstream areas and exotic salmonids are more abundant in Upstream areas, consistent with previous studies of juvenile fish distributions in this river (Tagliaferro et al., 2014a) and with seasonal studies on fish assemblages over three years (Tagliaferro, 2014).
This study supports the prediction of having a more complex food webs with a wider base and an extra trophic level (due to the presence of Lake Trout) in Upstream sections. This section presents a more heterogeneous habitat structure (Tagliaferro et al., 2014b; Quiroga et al., 2015) where the river runs through gravel bars and small gravel islands, which were associated with a more complex macroinvertebrate community structure with higher richness and biomass (Tagliaferro et al., 2013; Tagliaferro, Pascual, 2017). Moreover, using SIA, we noted that in Upstream sections, food webs are based on algae as the basal energy source; while in Midstream sections the main resource is fine debris (mainly parts of Myriophyllum sp.) that might have a lower energetic value and not be able to sustain complex food webs. Also, the two river sections differ in the trophic position and role of the most abundant invasive species, the Rainbow Trout. Previous research by Tagliaferro et al. (2014a) showed that 25% of fish captured in the lower part of the watershed were Rainbow Trout and 75% Puyen; in that case we expect Rainbow Trout to be the apex predator, without top down controls from larger fish (e.g., Lake Trout) feeding on yearlings, juveniles or adult Trout. More suitable interactions between small fish of the two species would be competition, and predation on Puyen by larger Rainbow Trout. However, in Upstream sections, the role of Rainbow Trout might change from prey and competitor to top predator depending on the abundance of other piscivorous fish taxa such as, Brown Trout, Perch, and Lake Trout.
Regarding our second hypothesis, where we proposed that Rainbow Trout and the native Galaxiid will experience a higher diet and isotopic overlap in Midstream sections we found that, although in Upstream sections a greater diversity of prey contributed to Rainbow Trout diet and in Midstream sections, larger stoneflies and fish had a greater contribution, similar isotopic signatures were found in both river sections. Thus, Rainbow Trout might have a stronger effect on Puyen populations in Upstream sections due to predatory effect and possible competition. Since Puyen and Rainbow Trout did not have significant differences in isotopic values in Upstream sections, but changes in feeding behavior of Galaxiids occur in the presence of Trout (Elgueta et al., 2013; Cussac et al., 2020), we propose that these species might be feeding in different areas (i.e., deeper or littoral areas) of the river to reduce possible competition.
There is wide support for all salmonids having certain degrees of piscivory (Pascual et al., 2007), with Lake Trout being a top predator (Post et al., 2000; Tronstad et al., 2010; Syslo et al., 2016). There is also evidence of the predation of Brown Trout, Chinook and Coho Salmon on Puyen (Vila et al., 1999; Penaluna et al., 2009). While in much of the work studying the diet of introduced salmonids, selectivity (Di Prinzio, Casaux, 2012; Tagliaferro et al., 2014a), size of prey (Di Prinzio et al., 2015), overlap with native species (Kusabs, Swales, 1991; Tagliaferro et al., 2014a; Horká et al., 2017) are evaluated, this work adds the interaction with other introduced species of salmonids. Thus, we could observe that in areas where several introduced species coexist, natural interactions such as competition and predation by other salmonids occur and less pressure could be exerted on native species. For instance, the presence of Lake Trout have been associated to the decline of both native and invasive fish species (Tronstad et al., 2010). In addition, this study highlights the differences in distribution of some native species such as Perch and Puyen. Thus, the interaction between yearling Rainbow Trout and Puyen feeding on macroinvertebrates, and juvenile and adult Rainbow Trout feeding also on Puyen in Midstream section, get more complex in the Upstream section. In Usptream section, yearling Rainbow and Brown Trout together with juvenile Chinook Salmon and Puyen are feeding on macroinvertebrates, and adults are preying on Puyen and also on yearling Trout. In conclusion, whereas one invasive salmonid species can generate negative effects on native species on a new environment, when new invasive species are established, the associated changes are much more complex; for instance, the establishment of other invasive species may have opposite effects on native fauna since they might release or increase pressure on native species, for example, controlling the population abundance of other introduced species.
Two main effects have been highlighted throughout studies of the ecology of salmonid invasion: (1) the use of habitat and timing (Glova et al., 1992; McIntosh et al., 1992; Stuart-Smith et al., 2008; Correa et al., 2012; Sowersby et al., 2016), and (2) the use of food resources and possible interactions with native species (Glova et al., 1992; Shelton et al., 2016; Milardi et al., 2020). Differential selection of habitat or time of the day using a certain space could help reducing unnatural interactions between species (Stuart-Smith et al., 2008; Otturi et al., 2016). food webs can be altered in their structure and function through top-down or bottom-up mechanisms (Gozlan et al., 2010). On the other hand, by reducing native species, introduced fish can also change the ecofunctional diversity of a community (Milardi et al., 2020). Recent studies found that invasive fish species can diminish the relative diversity of native fish communities (Milardi et al., 2016; 2020), and alter their functional traits (Shuai et al., 2018). Although most abundant juvenile fish species in the Santa Cruz River, independently from their origin are considered generalized benthic predators (Lattuca et al., 2008; Di Prinzio et al., 2013; Tagliaferro et al., 2014a; Hertz et al., 2017), they might feed on different functional feeding groups, changing food webs structures. For example, the replacement of native fish by non-native Trout has been shown to reduced top-down control over collector-gatherer (Shelton et al., 2017). On the other hand, predator pressure over Galaxiid by native Perch and adult salmonids might indirectly affect macroinvertebrate abundances. The reduction of Galaxiids due to salmonids was also associated with changes in insect behavior and algal standing crops (Flecker, Townsend, 1994; Herrera-Martínez et al., 2017).
In the present study, we were able to determine trophic interactions and identify differences in trophic structure depending on the river section by using the two alternative techniques of stomach content and stable isotope analyses (Fig. 5). While SCA results in partially or completely digested organisms creating difficulties in the identification process, stable isotope techniques allow an easier way of integrating information from all components of the food webs. Puyen was the most abundant native species in both river areas with similar roles in the food webs, but the stable isotopes analyses indicated a higher trophic position in midstream areas, which might be due to the presence of fewer fish species allowing it to have a broader diet. Also, SIA integrate information over a greater time span (months to years), which is especially important to assess the trophic role when organisms are slow-growing fish (Hesslein et al., 1993; McCarthy et al., 2004) or spend several days without feeding (e.g., spawning Steelhead Rainbow Trout or Lake Trout). The time span was important to take into consideration with Lake Trout, Chinook Salmon, and Big Puyen, since the number of stomach content samples were low and the integration of time in the SIA support the same diet over several months. On the other hand, SCA has the advantage of providing taxonomic information for food items, which is not possible with SIA (Power et al., 2002).

FIGURE 5| Midstream and Upstream areas food webs scheme done considering stable isotopes and stomach content analysis. Dark arrows indicate a higher contribution to diet. Rainbow Trout is placed above Perch, Chinook, and Brown Trout since when analyzing stomach content, it trophic role depends on the ontogenetic stage. CF= collector-filterer, CG= collector-gatherer, SCR=scrapers, SHR= shredders, PRED= predator.
Stable isotope analyses were found to be a useful tool in evaluating possible energy sources according to δ13C values and trophic positions with δ15N values. The fractionation of 15N, usually assumed to be 3.2-3.4‰, in an animal in relation to its diet (Peterson, Fry, 1987; Post, 2002; Baeta, 2018) depends on environmental and individual conditions (Minagawa, Wada, 1984; Peterson, Fry, 1987; Wiederhold, 2015). Some factors affecting the fractionation of nitrogen are tissue type (Pinnegar et al., 2000; Vanderklift, Ponsard, 2003), quality of the diet (McCutchan Jr et al., 2003; Cashman et al., 2016), species being studied (DeNiro, Epstein, 1980; Arcagni et al., 2015; Sánchez-Carrillo, Álvarez-Cobelas, 2018), and transgenerational effects (Liberoff et al., 2013). Moreover, methods for understanding the results of SIA are still developing (Phillips, Gregg, 2003; Moore, Semmens, 2008; Parnell et al., 2010) and therefore the sources of variability that contribute to these methods have not yet been fully explored (Bond, Diamond, 2011). In the case of mixing models, the fractionation factor (or isotopic enrichment) is cited as one of the weakest points for the reconstruction of diets (Wolf et al., 2009). Statistical programs developed for analyzing food webs and diets such as SIAR (Parnell et al., 2010) have the possibility of incorporating fractionation values for each species, the concentration of 13C and 15N and values of standard deviation; however, the absence of some of these estimates may give erroneous results. In their study, Bond, Diamond (2011) showed that in most studies diet reconstruction with no information on species-specific fractionation values, generates studies where these values are considered fixed following the widely cited work of Post (2002), or were selected from taxonomically similar groups. The results of these investigations had a bias in the estimation of the diet, which should be checked and corroborated by other methods.
Stable isotope analyses also showed that Rainbow Trout involved in the present study corresponded to the resident type. Even though we used non-selective fishing techniques, the isotopic ranges for the most abundant invasive species were concordant with previously published values: δ15N = 8.8 ± 1.1‰ and δ13C = 23.2 ± 2.5‰ (Ciancio et al., 2008). Even though it has been reported that the probability of capturing the offspring of anadromous mothers might increase towards Upstream sections due to the suitability of the environment (Liberoff et al., 2015), the isotopic signature of Rainbow Trout in Mid and Upstream sections were concordant with resident types. In the present research, all relevant prey present in fish diets were sampled (except for the rare contribution of terrestrial prey), though an inconsistency in the isotopic enrichment between the value of δ15N of prey and Puyen and yearling Rainbow Trout was found. The absence of the isotopic value of terrestrial prey, mainly arthropods, could be generating a deficiency in the necessary sources for the use of mixing models in Puyen and yearling Rainbow Trout; however, the enrichment of the latter species varied up to ~6 ‰ units in δ15N and we expect another source to be contributing to this variation. In the absence of experimental studies on the fractionation of Puyen, or other Galaxiids, many questions arise: is it possible that the terrestrial contribution accounts for this difference between diet and isotopic values in Puyen? Secondly, might Puyen have a higher isotopic fractionation to improve the utilization of their prey in the Santa Cruz River? Regarding yearling Rainbow Trout, might the fractionation change between different life stages?
In conclusion, the information presented in this study shows the importance of the spatial pattern in aquatic food webs and species distribution in the Santa Cruz River. This data will be relevant when considering possible dam management in each section of the river where recreational and economical activities related to salmonids will be affected.
Acknowledgments
Funded by Consejo Nacional de Investigaciones Científicas y Tecnológicas and Agencia Nacional para la Promoción de la Ciencia y la Tecnología (FONCyT). This study was part of M.T. PhD thesis directed by Dr. M. Pascual in Grupo de Estudio de Salmónidos Anádromos (GESA). M. T. was supported by CONICET Graduate Fellowship. Centro Nacional Patagónico (CENPAT-CONICET) provided support for the optic service. The author thanks P. Quiroga and A. Liberoff for the help with fish collections. Ea. Río Bote, Ea. La Martina, Ea. San Ramón, Ea. La Marina, Los Plateados provided logistic support. This manuscript was highly improved by two anonymous reviewer’s and editor’s (Dr. Teixeira de Mello) suggestions.
References
Alvear PA, Rechencq M, Macchi PJ, Alonso MF, Lippolt GE, Denegri MA et al. Composition, distribution and trophic relationships of the fish fauna of the Negro river, Patagonia Argentina. Ecol Austral. 2007; 17(2):231–46.
Arcagni M, Rizzo A, Campbell LM, Arribére MA, Juncos R, Reissig M et al. Stable isotope analysis of trophic structure, energy flow and spatial variability in a large ultraoligotrophic lake in Northwest Patagonia. J Great Lakes Res. 2015; 41(3):916–25. https://doi.org/10.1016/j.jglr.2015.05.008
Arismendi I, Soto D, Penaluna B, Jara C, Leal C, León Muñoz J. Aquaculture, nonnative salmonid invasions and associated declines of native fishes in Northern Patagonian lakes. Freshw Biol. 2009; 54(5):1135–47.
Arismendi I, Penaluna B, Gomez-uchida D, Di Prinzio C, Rodríguez-olarte D, Carvajal-vallejos FM et al. Trout and Char of South America. In: Kershner JL, WilliamsJE, Gresswell RE, Lobón-Cerviá J, editors.Trout and char of the world. Bethesda: American Fisheries Society; 2019. 33p.
Arismendi I, Penaluna BE, Dunham JB, Garcia de Leaniz C, Soto D, Fleming IA et al. Differential invasion success of salmonids in southern Chile: Patterns and hypotheses. Rev Fish Biol Fish. 2014; 24:919–41. https://doi.org/10.1007/s11160-014-9351-0
Baeta A. Stable isotope ecology. In: Fath, BD, editor. Encyclopedia of Ecology Volume 3. Amsterdam:Elsevier; 2018; p.606–15.
Basulto del Campo S. El largo viaje de los salmones: una crónica olvidada, Propagación y cultivo de especies acuáticas en Chile [The long salmon trip: a forgotten chronicle, propagation, and cultivation of aquatic species in Chile]. Santiago: Gobierno de Chile; 2003.
Becker LA, Pascual MA, Basso NG. Colonization of the southern Patagonia ocean by exotic Chinook Salmon. Conser Biol. 2007; 21(5):1347–52. https://doi.org/10.1111/j.1523-1739.2007.00761.x
Bond AL, Diamond AW. Recent Bayesian stable-isotope mixing models are highly sensitive to variation in discrimination factors. Ecol Appl. 2011; 21(4):1017–23. https://doi.org/10.1644/13-mamm-a-014.1
Bowes RE, Thorp JH. Consequences of employing amino acid vs. bulk-tissue, stable isotope analysis: a laboratory trophic position experiment. Ecosphere. 2015; 6(1)1–12. https://doi.org/10.1890/ES14-00423.1
Brunet F, Gaiero D, Probst JL, Depetris PJ, Lafaye FG, Stille P. δ13C tracing of dissolved inorganic carbon sources in patagonian rivers (Argentina). Hydrol Process. 2005; 19(17):3321–44. https://doi.org/10.1002/hyp.5973
Buoro M, Olden JD, Cucherousset J. Global Salmonidae introductions reveal stronger ecological effects of changing intraspecific compared to interspecific diversity. Ecol Lett. 2016; 19(11):1363–71. https://doi.org/10.1111/ele.12673
Cashman MJ, Pilotto F, Harvey GL, Wharton G, Pusch MT. Combined stable-isotope and fatty-acid analyses demonstrate that large wood increases the autochthonous trophic base of a macroinvertebrate assemblage. Freshw Biol. 2016; 61(4):549–64. https://doi.org/10.1111/fwb.12727
Caut S, Angulo E, Courchamp F. Variation in discrimination factors (Δ15N and Δ13C): The effect of diet isotopic values and applications for diet reconstruction. J Appl Ecol. 2009; 46(2):443–53. https://doi.org/10.1111/j.1365-2664.2009.01620.x
Chapin III FS, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM, Reynolds HL et al. Consequences of changing biodiversity. Nature. 2000; 405:234–42. https://doi.org/10.1038/35012241
Ciancio JE, Pascual MA, Botto F, Amaya-Santi M, O’Neal S, Riva-Rossi CM et al. Stable isotope profiles of partially migratory salmonid populations in Atlantic rivers of Patagonia. J Fish Biol. 2008; 72(7):1708–19. https://doi.org/10.1111/j.1095-8649.2008.01846.x
Ciancio JE, Pascual MA, Lancelotti JL, Riva Rossi C, Botto F. Natural colonization and establishment of a Chinook Salmon, Oncorhynchus tshawytscha, population in the Santa Cruz River, an Atlantic basin of Patagonia. Environ Biol Fish. 2005; 74:219–27. https://doi.org/10.1007/s10641-005-0208-1
Clavero M, García-Berthou E. Invasive species are a leading cause of animal extinctions. Trends Ecol Evol. 2005; 20(3):110–20. https://doi.org/10.1016/j.tree.2005.01.003
Collins SM, Thomas SA, Heatherly T, MacNeill KL, Leduc AOHC, López-Sepulcre A et al. Fish introductions and light modulate food webs fluxes in tropical streams: A whole-ecosystem experimental approach. Ecology. 2016; 97(11):3154–66. https://doi.org/10.1002/ecy.1530
Correa C, Bravo AP, Hendry AP. Reciprocal trophic niche shifts in native and invasive fish: Salmonids and Galaxiids in Patagonian lakes. Freshw Biol. 2012; 57(9):1769–81. https://doi.org/10.1111/j.1365-2427.2012.02837.x
Correa C, Gross MR. Chinook Salmon invade southern South America. Biol Invasions. 2008; 10:615–39. https://doi.org/10.1007/s10530-007-9157-2
De Crespin De Billy V, Usseglio-Polatera P. Traits of brown trout prey in relation to habitat characteristics and benthic invertebrate communities. J Fish Biol. 2002; 60(3):687–714. https://doi.org/10.1006/jfbi.2002.1887
Cross WF, Baxter CV, Rosi-Marshall EJ, Hall RO, Kennedy TA, Donner KC et al. Food-web dynamics in a large river discontinuum. Ecol Monogr. 2013; 83(3):311–37. https://doi.org/10.1890/12-1727.1
Crowl TA, Townsend CR, McIntosh AR. The impact of introduced brown and rainbow trout on native fish: the case of Australasia. Rev Fish Biol Fish. 1992; 2:217–41. https://doi.org/10.1007/BF00045038
Cussac VE, Barrantes ME, Boy CC, Górski K, Habit E, Lattuca ME et al. New insights into the distribution, physiology and life histories of South American Galaxiid fishes, and potential threats to this unique fauna. Diversity. 2020; 12(5):1–16.
DeNiro MJ, Epstein S. Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta. 1978; 42(5):495–506.
DeNiro MJ, Epstein S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Acta. 1980; 45(3):341–51. https://doi.org/10.1016/0016-7037(81)90244-1
Di Prinzio CY, Casaux R. Dietary overlap among native and non-native fish in Patagonian low-order streams. Ann Limnol- Int J Lim. 2012; 48(1):21–30. https://doi.org/10.1051/limn/2011055
Di Prinzio CY, Miserendino ML, Casaux R. Feeding strategy of the non-native rainbow trout, Oncorhynchus mykiss, in low-order Patagonian streams. Fisheries Manag and Ecology. 2013; 20(5):414–25. https://doi.org/10.1111/fme.12028
Di Prinzio CY, Omad G, Miserendino ML, Casaux R. Selective foraging by non-native rainbow trout on invertebrates in patagonian streams in Argentina. Zool Stud. 2015; 54:e29. https://doi.org/10.1186/s40555-015-0108-9
Domínguez E, Fernández HR. Macroinvertebrados bentónicos sudamericanos: Sistemática y biología. Tucumán:Fundación Miguel Lillo; 2009.
Dyer BS. Systematic review and biogeography of the freshwater fishes of Chile. Estud Oceanol. 2000; 19:77–98.
Elgueta A, González J, Ruzzante DE, Walde SJ, Habit E. Trophic interference by Salmo trutta on Aplochiton zebra and Aplochiton taeniatus in southern Patagonian lakes. J Fish Biol. 2013; 82(2):430–43. https://doi.org/10.1111/j.1095-8649.2012.03489.x
Elliott JM. The food of brown and rainbow trout (Salmo trutta and S. gairdneri) in relation to the abundance of drifting invertebrates in a mountain stream. Oecologia. 1973; 12:329–47. https://doi.org/10.1007/BF00345047
Fausch KD. Introduction, establishment and effects of non-native salmonids: Considering the risk of rainbow trout invasion in the United Kingdom. J Fish Biol. 2007; 71:1–32. https://doi.org/10.1111/j.1095-8649.2007.01682.x
Flecker AS, Townsend CR. Community-wide consequences of trout introduction in New Zealand streams. Ecol Appl. 1994; 4(4):798–807.
Gallardo B, Clavero M, Sánchez MI, Vilà M. Global ecological impacts of invasive species in aquatic ecosystems. Glob Change Biol. 2016; 22(1):151–63. https://doi.org/10.1111/gcb.13004
Glova GJ, Sagar PM, Näslund I. Interaction for food and space between populations of Galaxias vulgaris Stokell and juvenile Salmo trutta L. in a New Zealand stream. J Fish Biol. 1992; 41(6):909–25. https://doi.org/10.1111/j.1095-8649.1992.tb02719.x
Gozlan RE, Britton JR, Cowx I, Copp GH. Current knowledge on non-native freshwater fish introductions. J Fish Biol. 2010; 76(4):751–86. https://doi.org/10.1111/j.1095-8649.2010.02566.x
Greig H, McIntosh AR. Indirect effects of predatory trout on organic processing in detritus-based stream food webs. Oikos. 2006; 112(1):31–40.
Hansen AG, Beauchamp DA, Schoen ER. Visual prey detection responses of piscivorous trout and salmon: Effects of light, turbidity, and prey size. T Am Fish Soc. 2013; 142(3):854–67. https://doi.org/10.1080/00028487.2013.785978
Herrera-Martínez Y, Paggi JC, García CB. Cascading effect of exotic fish fry on plankton community in a tropical Andean high mountain lake: a mesocosm experiment. J Limnol. 2017; 76(2):397–408. https://doi.org/10.4081/jlimnol.2017.1488
Hertz E, Trudel M, Tucker S, Beacham TD, Mazumder A. Overwinter shifts in the feeding ecology of juvenile Chinook Salmon. ICES J Mar Sci. 2017; 74(1):226–33. https://doi.org/10.1093/icesjms/fsw140
Hesslein RH, Hallard KA, Ramlal P. Replacement of Sulfur, Carbon, and Nitrogen in Tissue of Growing Broad Whitefish (Coregonus nasus) in Response to a Change in Diet Traced by d34S, d13C, and d15N. Can J Fish Aquat Sci. 1993; 50(10):2071–76. https://doi.org/10.1139/f93-230
Hobbs RJ, Arico S, Aronson J, Baron JS, Cramer VA, Epstein PR et al. Novel ecosystems : theoretical and management aspects of the new ecological world order. Glob Ecol Biogeogr. 2006; 1591):1–7. https://doi.org/10.1111/j.1466-822x.2006.00212.x
Horká P, Sychrová O, Horký P, Slavík O, Švátora M, Petrusek A. Feeding habits of the alien brook trout Salvelinus fontinalis and the native brown trout Salmo trutta in Czech mountain streams. Knowl Manag Aquat Ecosyst. 2017; 418(6). https://doi.org/10.1051/kmae/2016038
Jones ML, Stockwell JD. A rapid assessment procedure for the enumeration of salmonine populations in streams. N Am J Fish Manag. 1995; 15(3):551–62. https://doi.org/10.1577/1548-8675(1995)015%3C0551:ARAPFT%3E2.3.CO;2
Kling GW, Fry B, O’Brien WJ. Stable Isotopes and Planktonic Trophic Structure in Arctic Lakes. Ecology. 1992; 73(2):561–66. https://doi.org/10.1890/0012-9658(2006)87[892:TEOMOU]2.0.CO;2
Kusabs IA, Swales S. Diet and food resource partitioning in koaro, Galaxias brevipinnis (Günther), and juvenile rainbow trout, Oncorhynchus mykiss (Richardson), in two taupo streams, New Zealand. N Z J Mar Freshwater Res. 1991; 25(3):317–25. https://doi.org/10.1080/00288330.1991.9516485
Lancelotti JL, Pozzi LM, Yorio PM, Diéguez MC, Pascual MA. Precautionary rules for exotic trout aquaculture in fishless shallow lakes of Patagonia: minimizing impacts on the threatened hooded grebe (Podiceps gallardoi). Aquatic Conserv. 2010; 20:1–8. https://doi.org/10.1002/aqc
Lattuca ME, Battini MA, Macchi PJ. Trophic interactions among native and introduced fishes in a northern Patagonian oligotrophic lake. J Fish Biol. 2008; 72(6):1306–20. https://doi.org/10.1111/j.1095-8649.2008.01796.x
Liberoff AL, Quiroga AP, Riva Rossi C, Miller JA, Pascual MA. Influence of maternal habitat choice , environment and spatial distribution of juveniles on their propensity for anadromy in a partially anadromous population of rainbow trout (Oncorhynchus mykiss).Ecol Fresh Fish. 2015; 24(3):424–34. https://doi.org/10.1111/eff.12157
Liberoff AL, Riva Rossi C, Fogel ML, Ciancio JE, Pascual MA. Shifts in δ15N signature following the onset of exogenous feeding in rainbow trout Oncorhynchus mykiss: importance of combining length and age data. J Fish Biol. 2013; 82(4):1423–32. https://doi.org/10.1111/jfb.12063
López H, Miquelarena AM, Menni RC. Lista comentada de los peces continentales de la Argentina. ProBiota, FCNyM, UNLP, Serie Técnica-Didáctica. 2003; 5:1–85.
Macchi PJ. Respuestas de Galaxias maculatus a la depredacion por parte de Percichthys trucha y los salmónidos introducidos en ambientes lénticos de la Patagonia norte (Responses of Galaxias maculatus to predation by Percichthys trucha and introduced salmonids from lentic environment of North Patagonia). [PhD Thesis] Universidad Nacional del Comahue; 2004.
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz F. Invasiones biológicas: causas, epidemiología, consecuencias globales y control. Issues in Ecology. 2000; 5:689–710.
McCarthy ID, Fraser D, Waldron S, Adams CE. A stable isotope analysis of trophic polymorphism among Arctic charr from Loch Ericht, Scotland. J Fish Biol. 2004; 65(5):1435–40. https://doi.org/10.1111/j.0022-1112.2004.00526.x
McCutchan Jr JH, Lewis Jr WM, Kendall C, McGrath CC. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos. 2003; 102(2):378–90. https://doi.org/10.1034/j.1600-0706.2003.12098.x
McDowall RM. Impacts of Introduced Salmonids on Native Galaxiids in New Zealand Upland Streams: A New Look at an Old Problem. T Am Fish Soc. 2003; 132(2):229–38. https://doi.org/10.1577/1548-8659(2003)132%3C0229:IOISON%3E2.0.CO;2
McHugh PA, McIntosh AR, Howard S, Budy P. Niche flexibility and trout-Galaxiid co-occurrence in a hydrologically diverse riverine landscape. Biol Invasions. 2012; 14:2393–406. https://doi.org/10.1007/s10530-012-0237-6
McIntosh AR, Townsend CR, Crowl TA. Competition for space between introduced brown trout (Salmo trutta L.) and a native Galaxiid (Galaxias vulgaris Stokell) in a New Zealand stream. J Fish Biol. 1992; 41(1):63–81. https://doi.org/10.1111/j.1095-8649.1992.tb03170.x
Meador M, McIntyre JP, Pollock KH. Assessing the eficacy of single-pass backpack electrofishing to characterize fish community structure. T Am Fish Soc. 2003; 132(1):39–46. https://doi.org/10.1577/1548-8659(2003)132<0039:ATEOSP>2.0.CO;2
Merrit RW, Cummins KW. An introduction to the aquatic insects of North America. Dubuque: Kendall/Hunt Publishing Company; 1996.
Milardi M, Gavioli A, Soana E, Lanzoni M, Fano EA, Castaldelli G. The role of species introduction in modifying the functional diversity of native communities. Sci Total Environ. 2020; 699:134364. https://doi.org/10.1016/j.scitotenv.2019.134364
Milardi M, Siitonen S, Lappalainen J, Liljendahl A, Weckström J. The impact of trout introductions on macro- and micro-invertebrate communities of fishless boreal lakes. J Paleolimnol. 2016; 55:273–87. https://doi.org/10.1007/s10933-016-9879-1
Minagawa M, Wada E. Stepwise enrichment of 15N along food chains: Further evidence and the relation between d15N and animal age. Geochimt Cosmochim Acta. 1984; 48(5):1135–40. https://doi.org/10.1016/0016-7037(84)90204-7
Moore JW, Semmens BX. Incorporating uncertainty and prior information into stable isotope mixing models. Ecol Lett. 2008; 11(5):470–80. https://doi.org/10.1111/j.1461-0248.2008.01163.x
Muñoz I, Romaní AM, Rodrígues-Capítulo A, Esteban JG, Garcia-Berthou E. Relaciones tróficas en el ecosistema fluvial. In: Elosegi A, Sabater S, editors. Conceptos y técnicas en ecología fluvial. Bilbao: Fundación BBVA; 2009; p.1–22.
Nielsen JM, Popp BN, Winder M. Meta-analysis of amino acid stable nitrogen isotope ratios for estimating trophic position in marine organisms. Oecologia. 2015; 178:631–42. https://doi.org/10.1007/s00442-015-3305-7
Otturi MG, Battini MÁ, Barriga JP. The effects of invasive rainbow trout on habitat use and diel locomotor activity in the South American Creole perch: an experimental approach. Hydrobiologia. 2016; 777:243–54. https://doi.org/10.1007/s10750-016-2792-9
Parnell AC, Inger R, Bearhop S, Jackson AL. Source partitioning using stable isotopes: Coping with too much variation. PLoS ONE. 2010; 5(3):e9672. https://doi.org/10.1371/journal.pone.0009672
Pascual MA, Ciancio JE. Introduced anadromous salmonids in Patagonia: risks, uses, and a conservation paradox. In: Bert T, editor. Ecological and genetic implications of aquaculture activities. Dordrecht: Springer; 2007. p.333–53.
Pascual MA, Cussac VE, Dyer B, Soto D, Vigliano PH, Ortubay S et al. Freshwater fishes of Patagonia in the 21st Century after a hundred years of human settlement, species introductions, and environmental change. Aquat Ecosyst Health Manag. 2007; 10(2):212–27. https://doi.org/10.1080/14634980701351361
Pascual MA, Macchi P, Urbanski J, Marcos F, Riva Rossi C, Novara M et al. Evaluating potential effects of exotic freshwater fish from incomplete species presence-absence data. Biol Invasions. 2002; 4:101–13. https://doi.org/10.1023/A:1020513525528
Penaluna BE, Arismendi I, Soto D. Evidence of interactive segregation between introduced trout and native fishes in Northern Patagonian rivers, Chile. T Am Fish Soc. 2009; 138(4):839–45. https://doi.org/10.1577/T08-134.1
Peterson BJ, Fry B. Stable isotopes in ecosystem studies. Annu Rev Ecol Syst. 1987; 18:293–320.
Phillips DL, Gregg JW. Source partitioning using stable isotopes: Coping with too many sources. Oecologia. 2003; 136:261–69. https://doi.org/10.1007/s00442-003-1218-3
Pinnegar JK, Polunin NVC, Francour P, Badalamenti F, Chemello R, Harmelin-Vivien ML et al. Trophic cascades in benthic marine ecosystems: lessons for fisheries and protected-area management. Environ Conserv. 2000; 27(2):179–200. https://doi.org/10.1017/S0376892900000205
Post DM. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology. 2002; 83(3):703–18. https://doi.org/10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2
Post DM, Pace ML, Hairston NG. Ecosystem size determines food-chain length in lakes. Nature. 2000; 405:1047–49. https://doi.org/10.1038/35016565
Power M, Power G, Caron F, Doucett RR, Guiguer KRA. Growth and dietary niche in Salvelinus alpinus and Salvelinus fontinalis as revealed by stable isotope analysis. Environ Biol Fish. 2002; 64:75–85.
Power ME. Habitat heterogeneity and the functional signifiance of fish in river food webs. Ecology. 1992; 73(5):1675–88.
Power ME, Dietrich WE, Finlay JC. Dams and downstream aquatic biodiversity: Potential food webs consequences of hydrologic and geomorphic change. Environ Manag. 1996; 20:887–95.
Quiroga A, Lancelotti JL, Riva Rossi C, Tagliaferro M, García Asorey M, Pascual MA. Dams versus habitat: predicting the effects of dams on habitat supply and juvenile rainbow trout along the Santa Cruz River, Patagonia. Hydrobiologia. 2015; 755:57–72. https://doi.org/10.1007/s10750-015-2217-1
Riva Rossi C, Arguimbau M, Pascual MA. The range and timing of the spawning migration of anadromous rainbow trout in the Santa Cruz River, Patagonia (Argentina) through radio–tracking. Ecología Austral. 2003; 13(2):151–59
Rounick JS, Hicks BJ. The stable isotope ratios of fish and their invertebrate prey in four New Zealand rivers. Freshw Biol. 1985; 15(2):207–14.
de Ruiter PC, Neutel AM, Moore JC. Energetics, patterns of interaction strengths, and stability in real ecosystems. Science. 1995; 269(5228):1257–60. https://doi.org/10.1126/science.269.5228.1257
R Development Core Team. R: a language and environemnt for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. 2019. Available from: http://www.R-project.org/
Sánchez-Carrillo S, Álvarez-Cobelas M. Stable isotopes as tracers in aquatic ecosystems. Environ Rev.2018; 26(1):69–81. https://doi.org/10.1139/er-2017-0040
Shelton JM, Bird MS, Samways MJ, Day JA. Non-native rainbow trout (Oncorhynchus mykiss) occupy a different trophic niche to native Breede River redfin (Pseudobarbus burchelli) which they replace in South African headwater streams. Ecol Freshw Fish. 2017; 26(3):484–96. https://doi.org/10.1111/eff.12293
Shelton JM, Samways MJ, Day JA, Woodford DJ. Are native cyprinids or introduced salmonids stronger regulators of benthic invertebrates in South African headwater streams? Austral Ecol. 2016; 41(6):633–43. https://doi.org/10.1111/aec.12352
Shuai F, Lek S, Li X, Zhao T. Biological invasions undermine the functional diversity of fish community in a large subtropical river. Biol Invasions. 2018; 20:2981–96. https://doi.org/10.1007/s10530-018-1751-y
Simberloff D, Martin JL, Genovesi P, Maris V, Wardle DA, Aronson J et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol Evol. 2013; 28(1):58–66. https://doi.org/10.1016/j.tree.2012.07.013
Simon KS, Townsend CR. Impacts of freshwater invaders at different levels of ecological organisation, with emphasis on salmonids and ecosystem consequences. Freshw Biol. 2003; 48(6):982–94. https://doi.org/10.1046/j.1365-2427.2003.01069.x
Soto D, Norambuena F, Leal BC, Arismendi VI. Segundo Informe. Monitoreo Ambiental Salmonicultura Centros del Mar y Lago. Laboratorio de Ecología Acuática, Universidad Austral de Chile. 2003.
Sowersby W, Thompson RM, Wong BBM. Invasive predator influences habitat preferences in a freshwater fish. Environ Biol Fish. 2016; 99:187–93. https://doi.org/10.1007/s10641-015-0466-5
Stuart-Smith RD, White RWG, Barmuta LA. A shift in the habitat use pattern of a lentic Galaxiid fish: An acute behavioural response to an introduced predator. Environ Biol Fish. 2008; 82:93–100. https://doi.org/10.1007/s10641-007-9256-z
Syslo JM, Guy CS, Koel TM. Feeding ecology of native and nonnative salmonids during the expansion of a nonnative apex predator in Yellowstone lake, Yellowstone National Park. T Am Fish Soc. 2016; 145(3):476–92. https://doi.org/10.1080/00028487.2016.1143398
Tagliaferro M. Estructura espacial , temporal y trófica de las comunidades acuáticas del Río Santa Cruz. [PhD Thesis]. Universidad de Buenos Aires. 2014.
Tagliaferro M, Miserendino ML, Liberoff AL, Quiroga A, Pascual MA. Dams in the last large free-flowing rivers of Patagonia, the Santa Cruz River, environmental features, and macroinvertebrate community. Limnologica. 2013; 43(6):500–09. https://doi.org/10.1016/j.limno.2013.04.002
Tagliaferro M, Arismendi I, Lancelotti JL, Pascual MA. A natural experiment of dietary overlap between introduced Rainbow Trout (Oncorhynchus mykiss) and native Puyen (Galaxias maculatus) in the Santa Cruz River, Patagonia. Environ Biol Fish. 2014a; 98:1311–25. https://doi.org/10.1007/s10641-014-0360-6
Tagliaferro M, Quiroga A, Pascual MA. Spatial Pattern and Habitat Requirements of Galaxias maculatus in the Last Un-Interrupted Large River of Patagonia: A Baseline for Management. Environ Nat Resources Res. 2014b; 4(1):54–63. https://doi.org/10.5539/enrr.v4n1p54
Tagliaferro M, Pascual MA. First spatio-temporal study of macroinvertebrates in the Santa Cruz River: a large glacial river about to be dammed without a comprehensive pre-impoundment study. Hydrobiologia. 2017; 784:35–49. https://doi.org/10.1007/s10750-016-2850-3
Townsend CR. Individual, population, community, and ecosystem consequences of a fish invader in New Zealand streams. Conserv Biol. 2003; 17(1):38–47. https://doi.org/10.1046/j.1523-1739.2003.02017.x
Tronstad LM, Hall RO, Koel TM, Gerow KG. Introduced Lake Trout produced a four-level trophic cascade in Yellowstone Lake. T Am Fish Soc. 2010; 139(5):1536–50. https://doi.org/10.1577/T09-151.1
Tulian EA. Acclimatization of American fishes in Argentina. Washington: Govt. print. off; 1908.
Vanderklift MA, Ponsard S. Sources of variation in consumer-diet δ15N enrichment: A meta-analysis. Oecologia. 2003; 136:169–82. https://doi.org/10.1007/s00442-003-1270-z
Vander Zanden MJ, Shuter BJ, Lester N, Rasmussen JB. Patterns of food chain length in lakes: A stable isotope study. Am Nat. 1999; 154(4):406–16. https://doi.org/10.1086/303250
Vila I, Fuentes LS, Saavedra M. Ictiofauna en los sistemas límnicos de la Isla Grande, Tierra del Fuego, Chile. Rev Chil Hist Nat. 1999; 72:273–84.
Vitousek PM. Biological invasions and ecosystem processes: towards an integration of population biology and ecosystem studies. Oikos. 1990; 57(1):7–13. https://doi.org/10.2307/3565731
Wiederhold JG. Metal stable isotope signatures as tracers in environmental geochemistry. Environ Sci Technol. 2015; 49(5):2606–24. https://doi.org/10.1021/es504683e
Wolf N, Carleton SA, Martínez del Rio C. Ten years of experimental animal isotopes ecology. Funct Ecol. 2009; 23(1):17–26. https://doi.org/10.1111/j.1365-2435.2009.01529.x
Authors

![]() Marina Tagliaferro1
Marina Tagliaferro1 ![]() ,
, ![]() Sean P. Kelly2 and
Sean P. Kelly2 and ![]() Miguel Pascual3
Miguel Pascual3
[1] Centro Austral de Investigaciones Científicas (CADIC) – CONICET. B. Houssay 200, Ushuaia (9410), Tierra del Fuego, Argentina. azulmarinita@gmail.com (corresponding author).
[2] Universidad de Puerto Rico-Río Piedras, Departamento de Biología. San Juan, Puerto Rico (00931). spkelly.84@gmail.com.
[3] Centro Nacional Patagónico – CONICET. Bld. Brown 2915, Puerto Madryn (9120), Chubut, Argentina. miguelafuera@gmail.com.
Authors’ Contribution 

Marina Tagliaferro: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing- original draft, Writing-review & editing.
Sean P. Kelly: Supervision, Visualization, Writing-original draft, Writing-review & editing.
Miguel Pascual: Conceptualization, Formal analysis, Funding acquisition, Investigation, Project administration, Resources, Writing-original draft.
Ethical Statement
This research project was conducted under the animal care regulations of CONICET.
Competing Interests
The authors declare no competing interests.
How to cite this article
Tagliaferro M, Kelly SP, Pascual M. First study of food webs in a large glacial river: the trophic role of invasive trout. Neotrop Ichthyol. 2020; 18(3):e200022. https://doi. org/10.1590/1982-0224-2020-0022
Copyright
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Distributed under
Creative Commons CC-BY 4.0

© 2020 The Authors.
Diversity and Distributions Published by SBI
![]() Accepted August 30, 2020 by Franco Teixeira de Mello
Accepted August 30, 2020 by Franco Teixeira de Mello
![]() Submitted April 6, 2020
Submitted April 6, 2020
![]() Epub October 09, 2020
Epub October 09, 2020